|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
STRUCTURA MATERIALULUI GENETIC
1. SUBSTRATUL BIOCHIMIC AL EREDITATII
1.1. Dovezile indirecte care indicau substratul biochimic al ereditatii
ADN - 1869, extras din leucocitele umane, functia necunoscuta
o ulterior - izolat din diferite tipuri de nuclei = acid nucleic
1910 - 2 tipuri de acizi nucleici: acid deoxiribonucleic si acid ribonucleic
1924 - studii cu coloranti afinitate = cromosomii formati din ADN si proteine
toate celulele somatice - contin o cantitate constanta de ADN
o cantitate variabila de ARN si proteine
proteinele - foarte diverse ca varietate de la un tip celular la altul
nucleii celulele din seria germinala (meiotice) = ½ din ADN-ul celulelor somatice
geneticienii au respins posibiul rol ereditar al ADN
1.2. Identificarea substratului biochimic al ereditatii
v Experienta Griffith - 1928 a identificat colonii mutante de pneumococ
o injectarea soarecilor cu pneumococi (S) vii => pneumonie mortala
o injectarea soarecilor cu pneumococi (S) morti = lipsa oricarui efect
o injectarea soarecilor cu pneumococi (R) vii si pneumococi (S) morti
=> uneori pneumonie mortala
Concluzie:
o exista ceva material in pneumococii S
o care transmitea tulpinilor R fenotipul de virulenta (prezenta capsulei)
Griffith nu a putut explica acest fapt
o a atribuit ciudatul efect unui ipotetic principiu de transformare
nici aceasta descoperire nu a reusit sa atraga atentia geneticienilor.
v Experienta Avery, MacLeod si McCarty - 1944
o extractul fara lipide = activ
o extractul fara polizazaride = activ
o extractul fara o parte din proteine = activ
o extractul fara acizi nucleici = inactiv
o culturi R + extract de ADN din tulpinile S + proteaze = colonii R si S
o culturi R + extract de ADN din tulpinile S + ribonuclaze = colonii R si S
o culturi R + extract de ADN din tulpinile S + deoziribonucleaze = colonii R
Concluzie: substratul biochimic al ereditatii este ADN
prin intermediul lui se transmitea fenotipul S la pneumococii R
Aceasta descoperire a constituit a-II-a mare piatra de hotar, dupa ipoteza lui Beadle si Tatum, care marca inceputul geneticii moderne.
v Nici aceste rezultate nu au fost acceptate usor
v Experienta A.Hershey si Martha Chase - 1952
Concluzii:
v numai ADN fagic patrunde in bacterii pentru a le infecta
v proteinele raman in afara celulelor
v dupa multiplicarea fagilor
50% din 32P fixata de generatia anterioara era prezenta la descendenti
S transmisa la descendenti era sub 1% din cea fixata de ascendenti
v ulterior - suportul ereditatii la metazoare si metafite este tot molecula de ADN
v ADN este suportul universal al ereditatii in lumea vie, exceptie virusii cu ARN
1. Compozitia chimica a ADN
ADN = macropolimer: acid fosforic + glucid + baza azotata
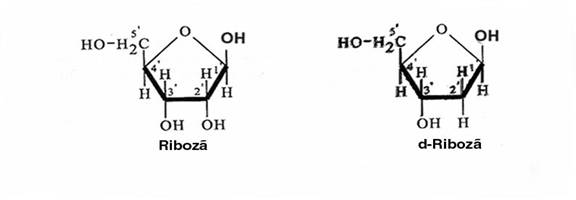
Figura 3-1. Structura chimica a ribozei si d-ribozei.
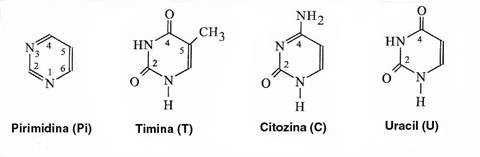
Figura 3-2. Structura chimica a pirimidinei si a bazelor pirimidinice.
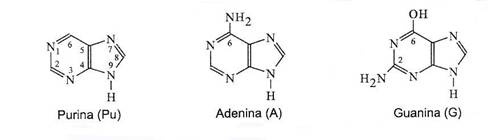
Figura 3- Structura chimica a purinei si a bazelor purinice.
Nucleozide = dezoxinucleozide (d-nucleozide):
adenina + d-riboza = dezoxiadenozina (d-adenozina)
guanina + d-riboza = dezoxiguanozina (d-guanozina)
timina + d-riboza = dezoxitimidina (d-timidina)
citozina + d-riboza = dezoxicitidina (d-citidina)
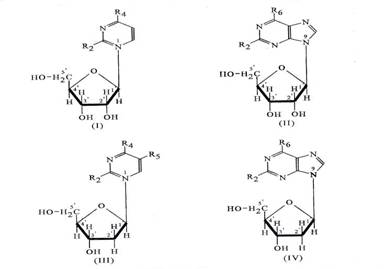
Figura 3-4. Structura chimica a nucleozidelor.
nucleozid + acid fosforic (P) = nucleozid 5'-monofosfat sau nucleotid:
2'-deoxiadenozina + acid fosforic = 2'-deoxiadenozin 5'-monofosfat (dAMP) sau acid 2'-deoxiadenozin 5'-monofosforic;
2'-deoxiguanozina + acid fosforic = 2'-deoxiguanozin 5'-monofosfat (dGMP) sau acid 2'-deoxiguanozin 5'-monofosforic;
2'-deoxitimidina + acid fosforic = 2'-deoxitimidin 5'-monofosfat (dTMP) sau acid 2'-deoxitimidin 5'-monofosforic;
2'-deoxicitidina + acid fosforic = 2'-deoxicitidin 5'-monofosfat (dCMP) sau acid 2'-deoxicitidin 5'-monofosforic.
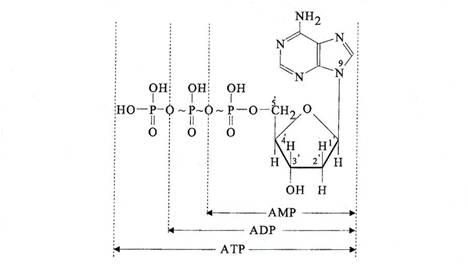
Figura 3-5. Structura chimica a d-AMP, d-ADP si dATP.
curent - simbolurile: dAMP, dGTP, dTMP si dCTP
nucleotidele constitue monomerii (caramizile) din care este construit ADN-ul
Monomerii se leaga prin legaturi fosfodiesterice intre radicalii 5'-P si 3'-OH
la capatul unei macromolecule de ADN se gaseste
o in pozitia 5' un radical P
o in pozitia 3'un radical -OH
v orientarea moleculei este in sensul 5'-3'
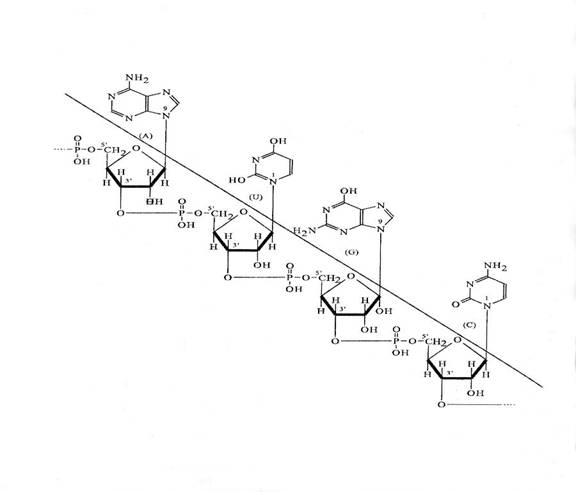
Figura 3-6. Legaturile fosfodiesterice din catena ribozofosforica.
Nr. molecule de d-riboza sau de acidul fosforic = nr. Monomeri
compozitia in d-riboza si in radicali P constituie partea invariabila a structurii ADN
E.Chargraff - 1950 -compozitia in baze azotate si raportul dintre ele:
o concentratia purinelor = concentratia pirimidinelor
[A] + [G] = [C] + [T]
o concentratia unei purine - A = concentratia unei pirimidine - T
numarul lor este egal, adica raportul lor este unitar [A] / [T] = 1
o concentratia celeilalte purine - G = concentratia celeilalte pirimidine - C
numarul lor este egal, respectiv ca si raportul lor este unitar [G] / [C] = 1
o compozitie baze - neunitara (proportia dintre bazele purinice sau pirimidinice)
raportul ([G] + [C] ) / ([G] + [C]) + ([A] + [T]) 1 = % G+C
Tabelul 3-1. Compozitia in baze la diferite specii [%].
|
Specia |
A |
T |
G |
C |
G + C |
|
Fag T-7 | |||||
|
E. coli | |||||
|
Neurospora | |||||
|
Porumb | |||||
|
Drosophila |
| ||||
|
Somon | |||||
|
Om |
regulile lui Chargraff
o constante pentru toate celulele unui organism = constanta de specie
o domeniul de variatie la diferitele specii - cuprins intre 26-74%
2. STRUCTURA FIZICA A MOLECULEI DE ADN
consecinta a regulilor lui Chargraff
o proportia bazelor in compozitia moleculei de ADN nu este intamplatoare
o evaluarea % GC => predictie sigura si facila a % celor 4 baze azotate
exemplu: compozitia in GC este 44% => G = 22% si C = 22%
AT = 100 - 44 = 56 => A = 56/2 = 28 si T = 56/2 = 28
concluzia - secventa de baze = modalitatea de stocare a informatiei genetice
J.Watson si F.Crick - 1952 - difractie cu raze X pe moleculele de ADN purificate:
o ADN- forma regulata de helix, pasul = 34Ǻ, 10 nucleotide / tur de spirala
o ADN - densitate crescuta => helix cu doua lanturi polinucleotidice
o ADN - diametru constant => perechi de baze intre o purina si o pirimidina
doua purine - d > 20
doua pirimidine - d < 20 Ǻ
Watson si Crick au propus modelul in dublu helix al structurii ADN
o 2 lanturi polinucleotidice cu orientare antiparalela - sens 5' =>
o 2 lanturi - cuplate prin legaturi de hidrogen - steric - 2 posibilitati:
A - T = 2 legaturi
G - C = 3 legaturi
Figura 3-10. Complementaritatea perechilor de baze.
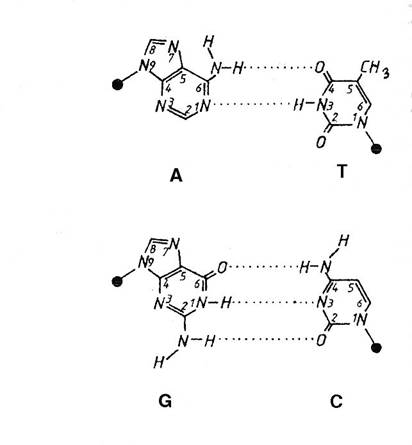
ADN - trasatura esentiala = pb => complementaritatea => matrita
o flexibilitatea conformationala a moleculei => forme alternative (A, B si Z)
difera intre ele prin pb / tura si prin orientarea lor
formele B si A prezinta o rotatie spre dreapta a helixului
forma B = conformatia ADN in conditii fiziologice
forma A - la concentratii saline mari
forma Z - orientata spre stanga = varianta minora
formele A si Z -unt interconvertibile
v Structura in dublu helix si complementaritatea reprezinta:
o una din cele mai mari descoperiri stiintifice din toate timpurile
o a-3-a piatra de hotar - genetica clasica => genetica moleculara
Descoperirea structurii ADN a permis intelegerea
o modului de stocare a informatiei ereditare
o modului cum aceasta informatie se poate propaga pe doua filiere distincte
ADN => ADN = replicare => reproducerea celulelor si organismelor
ADN => ARN => produsi genici =>
dogma centrala a geneticii moleculare
exprimarea informatiei genetice in ontogeneza si adaptare
Macromoleculele de ADN observabile cu microscopul optic:
o cromatina nucleara intre diviziunile celulare (in interfaza)
o cromosomii in timpul diviziunii celulare (in mitoza si meioza)
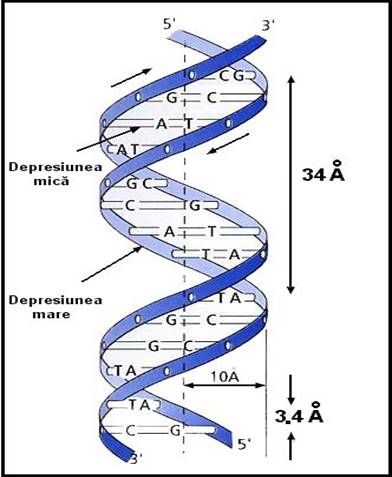
Figura 3-7. Modelul in dublu-helix al ADN.
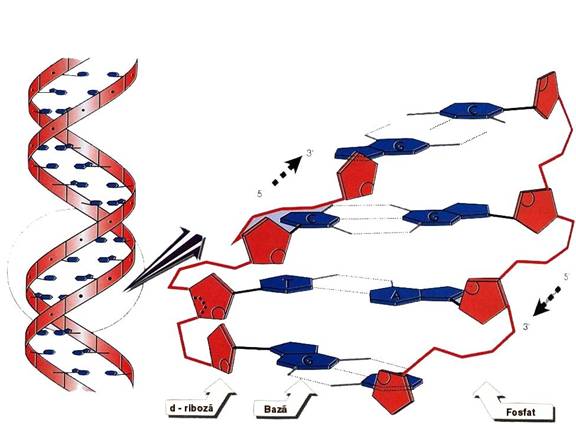
Figura 3-8. Structura tridimensionala a ADN.
Figura 3-9. Cuplarea celor doua catene antiparalele prin legaturi de hidrogen.
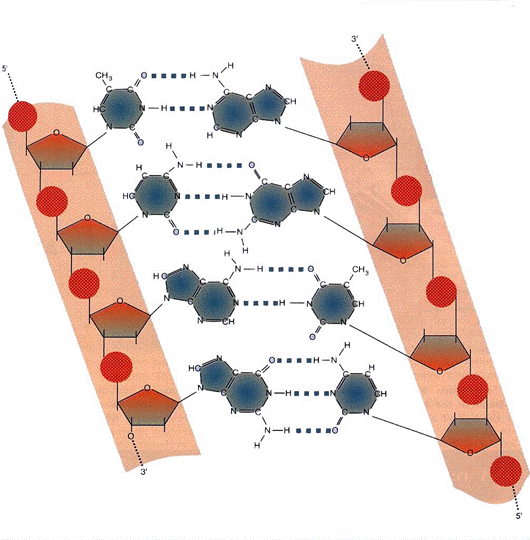
ARHITECTURA MACROMOLECULARA A ADN
LA EUCARIOTE
Numarul moleculelor de ADN (cromosomilor) = caracteristica de specie = 2n
o organismele superioare primesc de la fiecare genitor n cromosomi =>
o set haploid de cromosomi = genomul - genomul uman = 23 de cromosomi
o setul diploid cuprinde 46 de cromosomi
contin 6 X 109 pb
lungimea lineara virtuala de 1,8 m (1.800.000 m)
cromosomul 1 = 82 mm (82.000 m)
cromosomul 21 = 14 mm (14.000 m)
lungimea masurata la microscop
cromosomul 1 = 0,01 mm (10 m)
cromosomul 21 = 0,002 mm (2 m)
diametrul = 0,6 m
o scurtare de ordinal a 7.000-8.000 de ori
o grad foarte mare de impachetare = arhitectura moleculara complexa
1. Unitatea structurala de baza a cromatinei: nucleosomul
Cromatina = structura observabila microscopic in nucleii interfazici
o compusa din ADN, ARN, proteine histonice si proteine nehistonice
o histone = proteine mici = 100-200 a.a., 20-30% reziduuri de Lis si Arg
caracter bazic = electric pozitiva - rol = legarea ADN
5 clase: H , H2a, H2b, H si H ½ din masa moleculei de ADN
secvente a.a. conservate la H2a, H2b si inalt conservate la H si H
exemplu diferenta de secventa vaca - mazare:
H = 4 a.a. din 135; H = 2 a.a. din 102
rata de mutatie = 0,6 a.a. / 100 a.a. / 100.000.000 ani
Nucleosomul
o miez = 8 molecule de histone, cate 2 molecule din H2a, H2b, H si H
o pe miez se infasoara de doua ori dublul helix de ADN = 200 pb
o histona H poate fi indepartata fara a afecta structura nucleosomului
o digestia enzimatica => reduce lungimea ADN de pe nucleosomi la 146 pb
diferenta de 200 - 146 = 54 pb = ADN de legatura sau ADN linker
o digestia, extractia si separarea ADN din cromatina =>
fragmente multiplu al unui fragment de baza de 200 pb
fragmente de 200pb, 400 pb, 600 pb, 800 pb etc =>
nucleosomul = stuctura invarianta a cromatinei, indiferent ca este sub forma de heterocromatina, eucromatina sau cromosomi
o nucleosomul - forma unui sirag de margele
o particole cilindrice cu diametrul de 11 nm si inaltimea de 6 nm
4 nm = grosimea celor doua spire de ADN infasurate pe miez
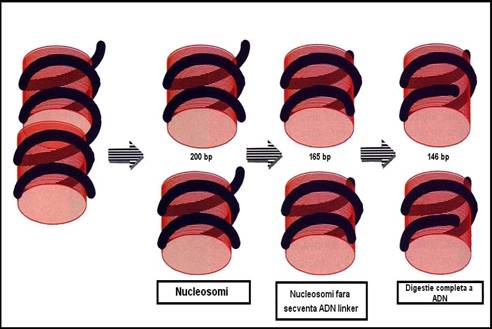
Figura 3-11. Nucleosomul: 2½ spire de ADN infasurate pe miezul proteic. Din cele 2½ spire de ADN o parte pot fi digerete cu nucleaze, iar cealalta nu poate fi indepartata de pe miezul proteic.
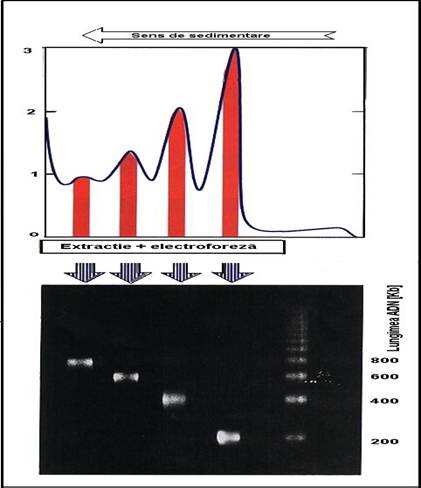
Figura 3-12. ADN-nucleosomic ca unitate invarianta.
2. Impachetarea ADN in cromosomi
Impachetarea ADN are grade succesive prin care este posibila
o adaptaarea materialului genetic la volumul nucleului
o controlul exprimarii genelor de pe cromosomi
- 1. Fibra (subtire) ADN de 11 nm
o un sir de nucleosomi - interactiunea ADN-histone => tensiune =>
o tendinta de suprarasucire cu conservarea flexibilitatii ADN =>
o reducere a lungimii liniare a ADN circa 7 ori
o se gaseste in structura eucromatinei, heterocromatinei si a cromosomilor
- 2. Fibra (groasa) ADN de 30 nm
o rasucire sub forma de solenoid a fibrei de 11 nm stabilizata de histona H1
o se formeaza prin rasucirea a 6 nucleosomi / tura
o indice de impachetare de 40 (reduce lungimea de 40 ori)
o se gaseste in structura eucromatinei, heterocromatinei si a cromosomilor
Buclele de cromatina
o formate prin indoirea fibrelor de 30 nm intr-o structura cu lungimea de 300 nm (1/2 din grosimea unei cromatide)
o impachetarea: eucromatina = 1.000-2.000X; cromosomi = 7.000-8.000X
o o bucla = 75 kb - atasata la un ax proteic central prin sute de pb - AT
- 4. Cromatida
o una din cele doua jumatati simetrice ale unui cromosom metafazic
o reprezinta o molecula de ADN ce rezulta dupa replicare
o poate stoca in medie o secventa de 80 Mb
o cromatidele <= spiralizare ax proteic central de care sunt atasate buclele
grosime de circa 600nm - 2 cromatide = 1.400 nm
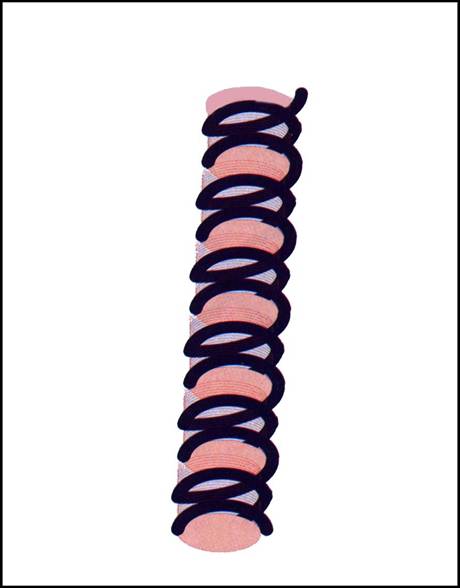
Figura 3-1 Fibra (subtire) de ADN de 11 nm.
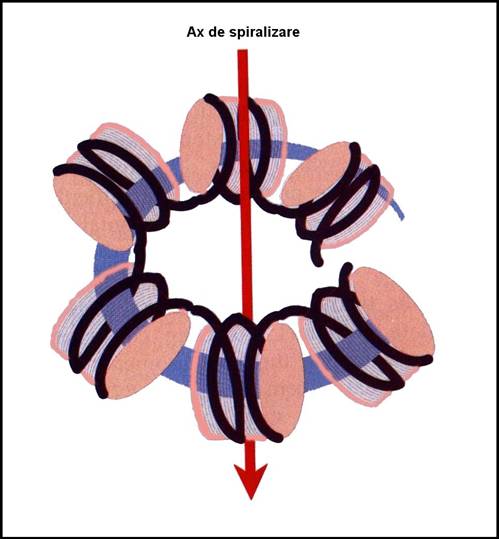
Figura 3-14. Fibra (groasa) de ADN de 30 nm.
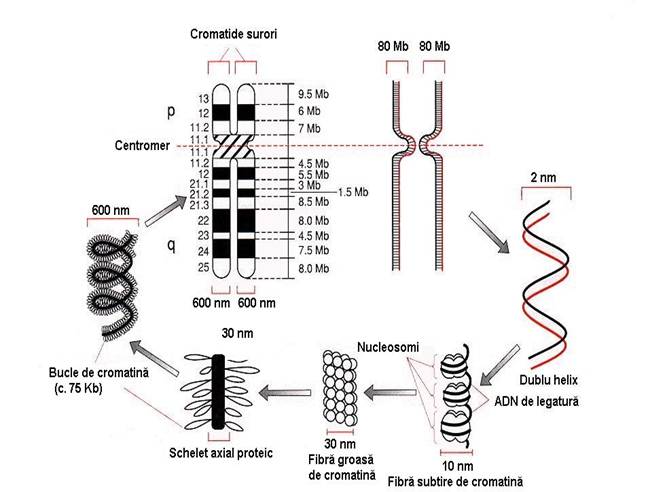
Figura 3-15. Gradele de impachetare succesiva a ADN in cromosom.
Aspectul microscopic al ADN
variaza in functie de ciclul celular compozitia = constanta
aspectul diferit - datorat gradului diferit de impachetare
1. Aspectul ADN in interfaza
Intre diviziunile celulare - ADN = forma de corpusculi cu refringenta diferita
o reprezinta in ansamblu cromatina nucleara
o evidentiati cu coloranti bazici
1) eucromatina = fibre groase (300 nm) de ADN
o impachetare mai putin densa (1.000-2.000 ori) slab colorata
o relativ dispersata in nucleu - sub forma unor filamente foarte fine
o se condenseaza numai in timpul diviziunii celulare
o este activa genetic, dar nu toate genele sunt active simultan
2) heterocromatina = zone dens impachetate ale moleculei de ADN
o densitate comparabila cu cea a cromosomilor din diviziune
o vizibila sub forma unor corpusculi densi, refringenti
o este inactiva genetic
2.a. Aspectul ADN in diviziunea celulara
cromosomii = corpusculi filamentosi (cromatide) dens colorati
o pereche = identici (bicromatidici) in profaza si metafaza
o neimperecheati (monocromatidici) in anafaza si telofaza
o fiecare cromatida: un centromer + 2 brate + 2 telomere
centromer - constrictie primara
o constituit dintr-un complex proteic, numit kinetocor
o locul de atasare a fibrelor fusului de diviziune
o centromerul contine zone de heterocromatina constitutiva
la nivel molecular - secventa => segregarea cromatidelor surori
pierderea aceste secvente = pierderea capacitatii de segregare
o zonele centromerice 1, 9, 16 si Yq = heterocromatina constitutiva
polimorfism genetic = variatii de la individ la individ
bratele cromosomilor
o centromerul imparte cromatida in doua brate, lung (q) si scurt (p)
o clasificarea morfologica - functie de raportul lungimii bratelor cromosomiale
metacentrici q p
submetacentrici q > p
acrocentrici q >> p
telomere = structuri specializate - contin heterocromatina constitutiva
o compozitie specifica - secvente intre 250-1.500 pb bogate in GC
o roluri
mentinerea integritatii structurale impiedecand digestia prin nucleaze
asigurarea replicarii complete a moleculei de ADN
imperecherea cromosomilor omologi
stabilirea structurii tridimensionale a nucleului
2.b. Aspectul ADN in diviziunea celulara
gradul de impachetare = 7.000-8.000 ori - variaza in functie de:
o conformatia cromatinei,
o timpul de replicare
o compozitia in baze
o densitatea de gene
o numarul de secvente repetitive
marcajul in benzi - succesiune de benzi colorate si interbenzi mai putin colorate
o coloratii specifice pentru ADN si tratamente specifice
digestie enzimatica, denaturare termica, solutii hipertone
o aranjamente liniare specifice pentru fiecare cromosom
o tehnici diferite
bandarea G - benzi G pozitive si negative
colorant Giemsa = amestec de azur si eozina
coloreza slab secventele GC si inchis pe cele bogate in AT
o banda = 1,5 Mb
identifica 550 benzi pe un set hapliod (genom)
bandarea R - invers decat benzile G
bandare Q - similara cu benzile G - florocrom - quinacrina
bandare C - specifica pentru heterocromatina centromerica
bandare T - specifica pentru heterocromatina telomerica
o aplicabilitate medicala = cariotipul prin marcaj in benzi
functiile cromosomilor
o perpetuare ADN => mitoza => dezvoltare ontogenetica
o perpetuare ADN de la o generatie la alta <= gametogeneza
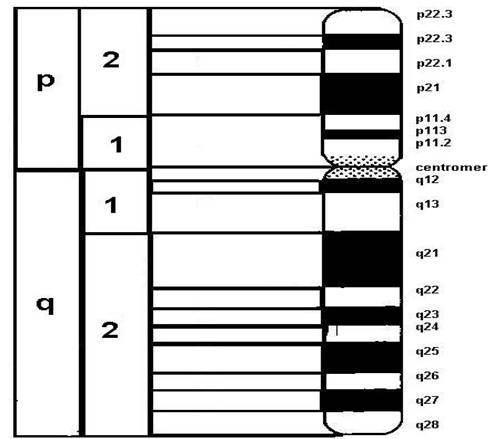
Figura 3-16. Morfologia cromosomului la eucariote.
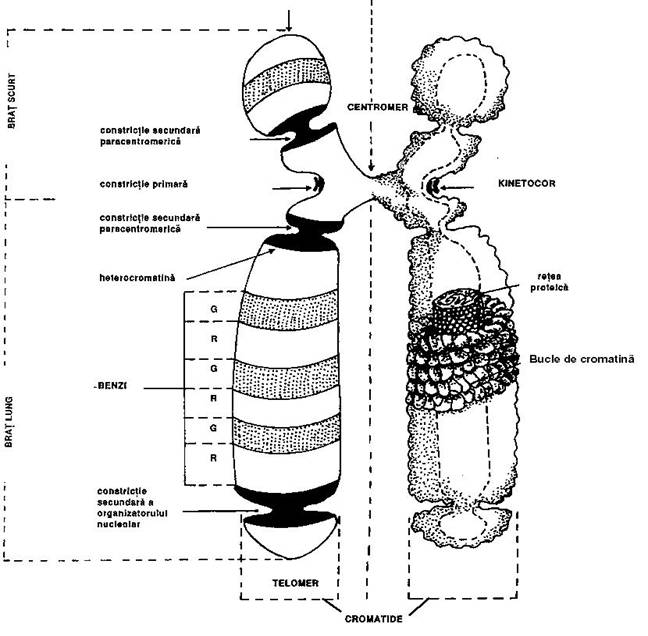
Figura 3-17. Organizarea moleculara a cromosomilor eucariotici.

Figura 3-18. Marcajul cu benzi G al cromosomilor umani.
4. ORGANIZAREA INFORMATIONALA A ADN: GENELE
geneticia clasica - gena = segment continuu si bine delimitat de pe molecula de ADN
P.Sharp (1977) - diferenta de lungime gena - transcript primar la adenovirusuri =>
o gena are secvente ADN care nu sunt informationale
W. Gilbert (1978) - ? - structura genomului viral este similara cu a celulei gazda
o genele eucariotelor = discontinuitate informationala
Ulterior s-a constatat ca genele sunt formate din:
o exoni = portiuni codificatoare
o introni = secvente necodificatoare
o gena incepe si se termina cu un exon
o promotor = colectie de secvente netraduse - la capatul 5' al genei
recunoscut de transcriptaze
locul de legare a enzimei - sinteza a mARN
o secventa terminator = secventa netradusa - la capatul 3'
marcheaza sfarsitul mesajului genetic
la capatul 3' - netradusa
o amplificatori (enhancers) - atenuatori (silencers) = secvente cu rol reglator
genetica moleculara - gena = secventa polinucleotidica de pe o molecula de ADN
o suportul unui program pentru sinteza unui produs specific
genele intrerupte au fost identificat in:
o genomul nuclear al metazoarelor si metafitelor
o genomul mitocondriilor si cloroplastelor
o genomul virusurilor
eucariotele = genom discontinuu / procariotele = genom continuu
4.1. Exonii
secvente polinucleotidice, conservate din interiorul unei gene
o codifica pozitia unui grup de aminoacizi dintr-un polipeptid
o secventele care se regasesc in mARN matur
numarul exonilor - foarte diferit - fiind putin corelat cu marimea genei
o gena distrofinei - lungime = 2.500 kb (2.500.000 pb) => 79 exoni
o gena ce codifica tipul VII de colagen - lungime = 31 kb => 118 exoni
o gena factorului VIII de coagulare - lungimea = 186 kb => 26 exoni
o gena CFTR - lungimea = 250 kb => 27 exoni
o orice gena are n exoni si n - 1 introni
lungimea medie a exonilor variaza in limite largi
o 50 pb - genele tARN
o 150 pb - genele insulinei si -globinei => 180 pb -gena distrofinei
toate genele umane = mai multi exoni, exceptie - gene histone si interferoni
4.2. Intronii
secvente polinucleotidice, mai putin conservate, din interiorul unei gene care
o nu codifica nici un produs specific
o exonii ar fi secventele care nu se regasesc in mARN matur
eliminati din transcriptul primar inainte de transportul mARN in citoplsma
o matisare (spicing) <= complex nucleoproteic - spliceosom
numar - foarte diferit - de la un intron => zeci => peste o suta de introni
o genele ce codifica unele specii de tARN - un singur intron
o genele insulinei si globinelor - doi introni
o genele factorului VIII de coagulare si a CFTR - 25, respectiv 26 introni
o numar introni => hibridizarea ADN genic - mARN matur complementar
zonele care nu hibridizeaza apar sub forma de ADN monocatenar
marimea cumulata a intronilor = 50 - 90% din lungimea genei
lungimea medie a intronilor = 102-103 pb
o la genele insulinei si globinelor - introni 500 pb
o la gena colagenului de tip VII - intronii 1.100 pb
o la gena fenialanin-hidroxilazei - introni 500 pb
functia intronilor - ? separare a genelor / reziduuri genetice ale evolutiei ADN
o in autoclivarea ARN / reglarea exprimarii genelor
o in unele cazuri - functii codificatoare
4. Promotorul
o succesiune de secvente inalt conservate, cu pozitie fixa, orientati 5'-3'
o situati pe aceiasi catena ADN ca si gena
o reprezinta codul de acces al transcriptazei la informatia genetica
factori de transcriere + promotor => ansamblarea ARN polimerazei
zona promotorului 200pb amonte de capatul 5' al genei:
o initiatorul - secventa -5 => +1 -= situsul de start al transcrierii
o cutia TATA = secventa heptanucleotidica TATAAAA
situata la pozitia -25-30 - rol de secventa de consens
modificarea ei prin mutatie duce la inactivarea genei
o cutia CAAT = secventa de 9 pb (GGCCAATCT)
situata in intervalul de la -70 la -80
influenteaza eficienta transcrierii
o secventa GC = secventa hexanucleotidica GGGCGG
situata in pozitia -110 - rol de crestere a eficientei transcrierii
o secventele octamerice ATTTGCAT dispersate in secventa promotor
secvente modulare = copii dispersate in promotor => modularea exprimarii genelor
4.4. Secventa terminator
secvente de trei baze (TCC, ATT si ATC) situate la capatul 3' al genei
o semnificatie de terminare a programului informational genetic = codon stop
secventa hexanucleotidica TTATTT situata in sensul 3' dupa codonul stop
o cu 11-30 nucleotide in fata situsului de poliadenilar, inalt conservata
o antisecventa AAUAAA de pe mARN are ca si functii marcarea
situsului de poliadnilare
locului de clivare al transcriptului
4.5. Intensificatorii si atenuatorii
enhancers, respectiv sillencers
secvente care potenteaza / diminueaza de 102-103 activitatea promotorilor
organizare similara cu a promotorilor
o situate la 102-103 pb in fata situsului de initiere
o pe aceiasi catena sau pe catena opusa = orientare 5'-3' sau 3'-5'
servesc ca si situsuri de legare specifica a proteinelor de reglare a genei
o stabilesc conexiuni cu complexul activ al ARN polimerazei
o functioneaza selectiv in functie de factorii tisulari
rol in diferentierea celulara
deosebirile dintre cele doua tipuri de secvente
o densitate mult mai mica a atenuatorilor
o posibilitatea lor de a fi localizati in unii introni
elementele de raspuns = secvente asociate cu gene specifice controlate prin:
o hormoni steroizi, factori de crestere, acid retinoic, cAMP etc.
4.6. Limbajul genetic
corelatie intre secventa bazelor - codificarea si stocarea informatiei in gene
ADN - 4 baze azotate = alfabet chimic cu o vechimea de peste 3 x 109 ani
o ARN o pirimidina - timina - inlocuita cu o alta pirimidina - uracilul
cod genetic => traducerea din limbajul bazelor azotate in limbajul aminoacizilor
asocierea bazelor in triplete - demonstrat de Niremberg, Mathaei si Ochoa in 1961
o triplet sau codon cuvant in limbajul genetic = 64 de combinatii posibile
o codonii - aliniati succesiv, in sensul 5'-3', fara secvente noncodificante
o 61 codificatori = codoni sens
o 3 nu codifica nimic = codoni nonsens
semnificatie = terminarea programului unei gene = codoni terminator
gena = secventa polinucleotidica dintre situsul de initiere si situsul terminator
o cadru de citire (reading frame) = trei posibilitati
o codonii nu se suprapun unctionare
o prin decalare => 3 posibilitati
2 apar codoni stop => mesaj inoperant
limbajul genetic este universal = organizare este comuna tuturor vietuitoarelor
Figura 3-19. Diferenta de lungime dintre gena si transcriptul primar.
Figura 3-20. Organizarea genelor la eucariote.
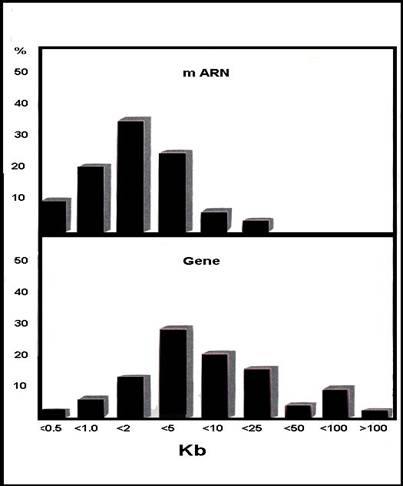
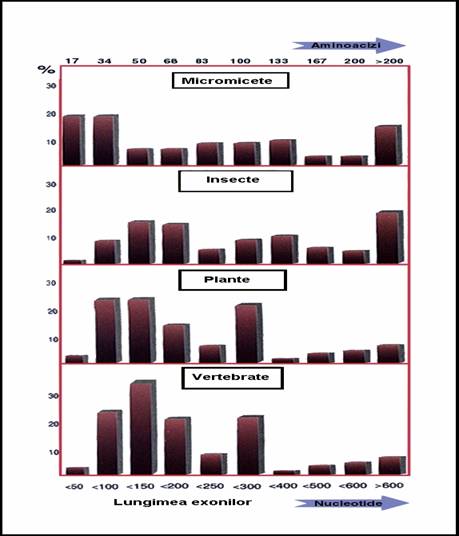
Figura 3-21. Variabilitatea lungimii exonilor la eucariote.
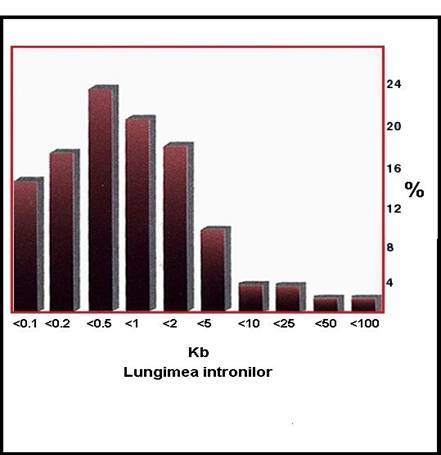
Figura 3-22. Variabilitatea lungimii intronilor la eucariote.
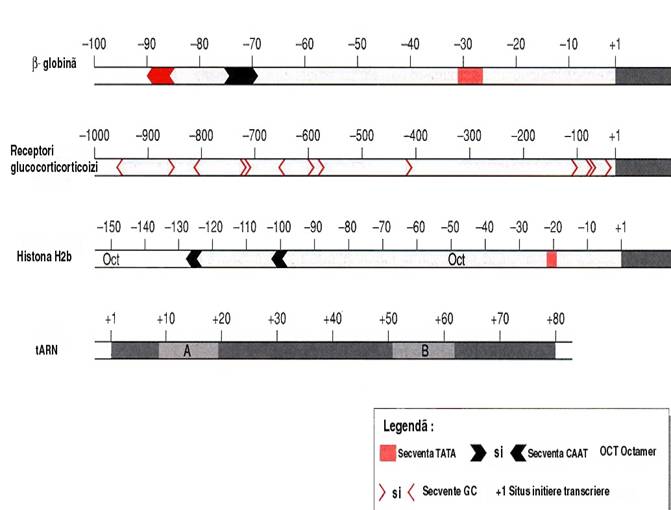
Figura 3-2 Organizarea secventelor promotor la eucariote.
Copyright © 2025 - Toate drepturile rezervate