|







 |  |
|
| |
 | Alimentatie | Asistenta sociala | Frumusete | Medicina | Medicina veterinara | Retete |
Citoscheletul si unele miscari celulare
Biologia celulei se intrepatrunde cu ecologia. Miscarile din fibrele musculare sunt implicate esential in relatiile trofice ale organismelor, in reproducere, in comunicare de informatii, aparare, raspandire, etc.
Spermatozoidul asigura cu flagelul sau fecundatia.
Cilii vibratili determina miscarile la foarte multe protiste, asigura eliminarea poluantilor din caile respiratorii iar cili speciali produc mecanoreceptia. Implicarea SNV este de asemenea notabila aici.
Pseudopodele cu citoscheletul lor foarte mobil asigura diapedeza si endocitozele necesare leucocitelor in apararea antiinfectioasa
Toate felurile de diviziuni cariochinetice implica structuri microtubulare de tipul fusului de diviziune cu implicatii esentiale in cresterea si dezvoltarea fiintelor care apartin la toate speciile de eucariote din ecosistemele cunoscute.
Patrundem in celula eucariota stabatand membrana celulara sau plasmalema. Citoplasma evidentiaza o retea de filamente subtiri. Aceste structuri se numesc diferentieri citoplasmatice si strabat tot spatial citoplasmei aflat in afara organitelor celulare , in spatiul citoplasmatic din afara organitelor celulare numit citosol, hialoplasma sau matricea citoplasmatica. In citosol, celula depoziteaza ca intr-un fel de magazie, rezervele ei de glicogen, lipide si ioni, etc. Citosolul este un fel de mediu intern al celulei care scalda toate organitele celulei, putand fi denumit si mediu interior pentru a distinge acest mediu de mediul intern al organismului pluricelular. In acest mediu interior al celulei au loc relatii metabolice intre el si organitele celulare. In citosol se produce un sir de reactii chimice de degradare a glucozei, numit glicoliza, tot aici glucoza se depune in glicogen prin glicogenogeneza si tot aici glicogenul e desfacut prin glicogenoliza in molecule de glucoza. In citosol se metabolizeaza aminoacizii si se sintetizeaza acizii grasi. In citosol exista ribozomi liberi care sintetizeaza proteine proprii celulei, nu proteine de export ca in reticulul endoplasmic rugos tapetat si el cu ribozomi.
Din punct de vedere fizic, citoplasma este o solutie de macromolecule proteice dizolvate in apa, deci un sistem omogen. Aceste macromolecule au dimensiuni de 10 la minus 5-10 la minus 7nm(nanometri, 1nm=10 la minus 9) si de aceea nu trebuie sa mire faptul ca citoplasma manifesta fenomene comune cu sistemele eterogene coloidale (cum sunt de, de exemplu, sarurile coloidale de aur): aspectul e opalescent (pentru ca particulele imprastie in toate partile lumina ce cade asupra lor; fenomenul Faraday-Tyndall) , in citoplasma observam miscarea browniana; la sedimentare prin centrifugare obtinem aranjate componentele citoplasmei in ordinea densitatii lor si de asemenea citoplasma e schimbatoare, prezentand tranzitii sol-gel. In starea de sol citoplasma ofera o miscare libera a particulelor in apa si capata o fluiditate mare. In starea de gel, particulele sunt legate intr-o retea in ochiurile careia se afla apa si astfel citoplasama capata o vascozitate mare sau foarte mare.
Exista treceri continue intre starea de sol si cea de gel si invers ceea ce da acel dinamism interesant al citoplasmei vii. Chiar si diferentierile citoplasmatice au un caracter foarte dinamic. In toate celulele eucariote gasim 4 tipuri de diferentieri citoplasmatice:
a)filamentele de miozina;
b)filamente de actina;
c)filamentele intermediare;
d)microtubulii.
Ceea ce este caracteristic pentru toate cele 4 tipuri de diferentieri este faptul ca ele sunt formate prin polimerizarea unor proteine monomer aflate in citosol.
a) Filamentele de miozina se formeaza prin polimerizarea moleculelor de miozina-macromolecule cu greutatea moleculara de 470 000 daltoni si alcatuite din 6 lanturi polipeptidice. Aceste filamente au aspectul unui baston cilindric gros si prevazut cu doua capete acoperite cu noduri si la mijloc e un fel de coada sau zona goala ce uneste cele doua capete. Capetele globulare ale filementului de miozina pot interactiona cu actina si de asemenea au activitate ATP-azica. Polimerizarea miozinelor se face prin cozile lor-filamentul de miozina are o zona goala sau nuda aflata la mijloc si doua zone mai groase la extremitati unde proemina capetele globulare ale miozinelor de jur imprejurul filamentului de miozina.
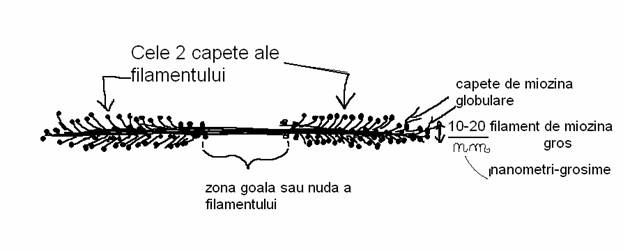
In celulele nemusculare filamentele de miozina sunt scurte si formate din 10-20 de molecule. Pe langa scurtimea lor mai sunt si labile. In celulele nemusculare miozina e asociata cu actina si are rol in unele miscari celulare.
In celulele musculare polimerizeaza aproximativ 500 de molecule de miozina si se formeaza filamente groase cu diametrul de 10-20 nm. Aceste sunt structuri stabile aranjate ordonat intr-o retea hexagonala impreuna cu filamentele de actina, cu care tot impreuna realizeaza contractia musculara.
b)Filamentele de actina. Daca ele se afla in citoplasma celulelor musculare, atunci ele mai contin si alte proteine asociate impreuna cu care formeaza filamentele subtiri. In celulele nemusculare, filamentele de actina poarta denumirea de microfilamente. Actinele sunt proteine cu greutate moleculara mica( in jur de 40 000 daltoni). Ele se observa in diferite celule cu structuri asemanatoare fiind in proportie de 5-15 % din totalul proteinelor celulei. Toate actinele au o propietate caracteristica prin faptul ca pot exista sub 2 forme: cea de monomer- actina globulara (G) si actina polimerizata filamentoasa (F) in care actinele monomer se insira ca margele dintr-o bratara sau colier. ATP (adenozintrifosfatul) accelereaza polimerizeaza actinele G sau monomer. Daca ATP se hidrolizeaza in ADP (adenozin-difosfat) si fosfat anorganic (Pi) se elibereaza energia necesara polimerizarii. Aceasta hidroliza are loc numai dupa legarea actinei G sau monomer in actina F sau polimer. Polimerizarea mareste vascozitatea citoplasmei, iar depolimerizarea produce o scadere a vascozitatii citoplasmei. Deci polimerizarea si depolimerizarea sunt strans corelate cu trecerile intre starea de gel si cea de sol a citoplasmei. Mucegaiurile citocalazine care perturba polimerizarea, iar ciuperca otravitoare mortala 100% numita palaria sarpelui- Amanita phalloides contine un alcaloid foarte toxic numit faloidina care tulbura grav polimerizarea actinelor.
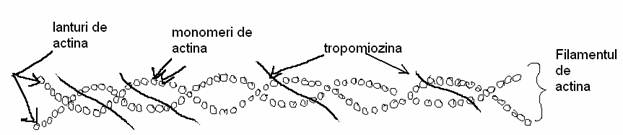
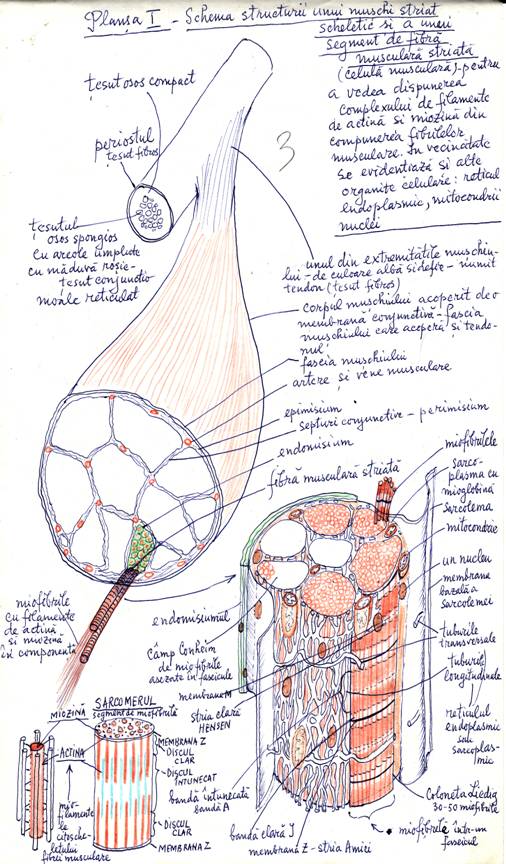
Microfilamentele apar mai ales in manunchiuri dispuse la periferia citoplasmei si in apropierea membranei celulare a celulelor nemusculare. Microfilamentele de actina intra in structura prelungirilor permanente ale celulei (cum sunt microvilii celulelor mucoasei intestinale, stereocilii celulelor receptoare ale epiteliului senzorial al urechii interne). Celulele vii aflate in culturi de celule au la suprafata lor prelungiri filifome temporare care de asemenea contin microfilamente de actina. In toate cazurile aratate microfilamentele au doua roluri: un rol structural de sustinere a prelungirilor celulare si un rol dinamic in miscarile ce au la baza mecanismul actina-miozina. Mecanismul molecular care elaboreaza toate aceste miscari este acelasi. Intre filamentele paralele de actina si miozina exista punti laterale ce leaga capetele globulare ale miozinelor cu monomerii de actina din filamentele de actina. Explicatia contractiei consta in desfacerea puntii cu o actina si refacerea ei cu actina urmatoare din microfilament. Fiecare cap de miozina se deplaseaza in lungul filamentului de actina de pe un monomer pe altul care urmeaza si trage pas cu pas filamentul de actina. Aceasta deplasare a fost asemanata cu mecanismul " rotii cu clicheti". Dupa contractie urmeaza relaxarea care se produce prin desfacerea simultana a tuturor puntilor si revenirea filamentelor in pozitia initiala. Acest mod de actiune este caracteristic pentru toata miscarile bazate pe interactiunile actina-miozina; exemplu: contractie musculara, miscarile din microvili si miscarea de locomotie amiboidala. Urmariti structura muschiului striat si al fibrei striate cu citoschelet action-miozinic - plansa I.
Microvilii sunt prelungiri permanente in forma de deget dispuse cu miile, unul langa altul pe suprafata membranelor celulare de la acele celule ce indeplinesc rol in absorbtie. Exemplu: celulele intestinale si celulele din tubul contort proximal renal . Microvilii favorizeaza absorbtia prin marirea suprafetei de absorbtie a membranei si printr-o interventie activa ce impinge substantele absorbite spre interiorul celulei.
Microvilii realizeaza aceasta prin contractia filamentelor de actina si miozina ce sunt aranjate longitudinal in interiorul lor.si care, ca niste micropistoane, imping in citoplasma substantele patrunse prin plasmalema sau membrana celulara.
Miscarea de deplasare amiboidala se observa atat la protozoare - exemplu amiba de unde si numele, dar si la animalele pluricelulare (metazoarele) si la om. Importanta medicala a acestei miscari consta in faptul ca acest mecanism foloseste globulelor albe -leucocitelor- pentru a se strecura prin peretele foarte subtire al vaselor capilare, pe care le parasesc si apoi folosind spatiile interstitiale stramte dintre celule se deplaseaza la locul infectiei spre "a se lupta " cu microbii pe care ii inghit la propriu prin fenomenul de endocitoza numit in acest caz fagocitoza. Alte celule denumite fibroplaste se deplaseaza pe suprafata ranilor depunand o proteina numita collagen care are rol in formarea cicatricei si vindecarea ranilor. Celula in deplasarea ei emite prelungiri care la protozoare sunt niste pseudopode cilindrice sau in cazul leucocitelor si fibroblastelor sunt pseudopode lamelare latite. In timpul formarii prelungirii citoplasmatice, citoplasma interna a acesteia sau endoplasma devine fluida si trece in starea de sol scurgandu-se in prelungirea care se alungeste. Ajunsa aproape de capatul prelungirii citoplasma se rastoarna pe fata interna a membranei celulare sau plasmalemei si se gelifica prin contractia actomiozinei. Prelungirea fiind fixata pe suport celula este trasa pe directia prelungirii. Prin treceri successive ale citoplasmei in stare de sol (endoplasma) si apoi de gel (ectoplasma), se produce miscarea. Aceasta miscare citoplasmatica a fost asemuita cu deplasarea "senilei unui tractor".
c)Filamentele intermediare au un diametru cu grosimea de 10 nm, deci au o valoare intermediara intre valorile diametrelor celor celor doua tipuri. Aceste filamente ale citoscheletului se afla mai ales in profunzimea citoplasmei cu toate ca se observa si la periferia celulei, de exemplu in desmozonii sau in prelungirile neuronului la axonul acestuia.
Aceste filamente sunt mai stabile ca cele de actina sau decat microtubulii. Odata ce s-au asamblat ele nu se destrama prin depolimerizare. Proteinele ce compun filamentele intermediare nu sunt globulare ci filiforme, iar polimerizarea se face printr-un fel de alaturare si impletire ca intr-o franghie ce le dau o rezistenta sporita. Acelasi filament are mai multe tipuri diferite de proteine. Aceste proteine difera dupa tipul celulei si dupa specie. Se observa 5 tipuri de filamente intermediare:
filamentul de cheratina sau tonofilamentele care sunt caracteristice celulelor epiteliale unde fac o retea neregulata ancorata in desmozomi ce asigura rezistenta la rupere a pielei si la alti factori mecanici. Pe masura imbatranirii filamentele de cheratina se acumuleaza in celulele epiteliale si se leaga prin punti disulfhidrice ce rezista si dupa moarte dand durabilitate stratului cornos al pielei, parului si unghiilor;
filamentele din neuroni (neurofilamentele) cu rol de sustinere a axonilor ce ating lungimea de 1 m la unii neuroni (neuronal are un singur axon si de obicei mai multe dendrite;
filamentele gliale sunt alcatuite din vimentina si dintr-o proteina gliala fibrilara acida observata in astrocite;
filamentele de desmina din muschii scheletici
striati si din muschii netezi (exceptand pe cei netezi din peretii vaselor de sange). Probabil ca la fel ca si neurofilamentele au rol de structural de sustinere a miofibrilelor vecine care au actina si miozina discutata. Filamentele intermediare nu indeplinesc functii bine clarificate, dar identificarea tipului de filamente intermediare folosind anticorpi cu afinitate speciala pentru proteinele ce le compun este de importanta practica pentru depistarea cancerului deoarece se stabileste tipul de celula din care provine o tumora. Cancerele celulelor epiteliale au filamente de cheratina, cancerele muschilor numite si sarcoame au filamente de vimentina, roadomiosarcoamele sau cancerele muschilor striate au filamente de desmina, glioamele - cancere ale celulelor gliale din tesutul nervos au filamente gliale, neuroblastoamele au neurofilamente. Chiar si cand apar metastaze se pastreaza si in acestea tipul de filament intermediar caracteristic celulei de origine a cancerului.
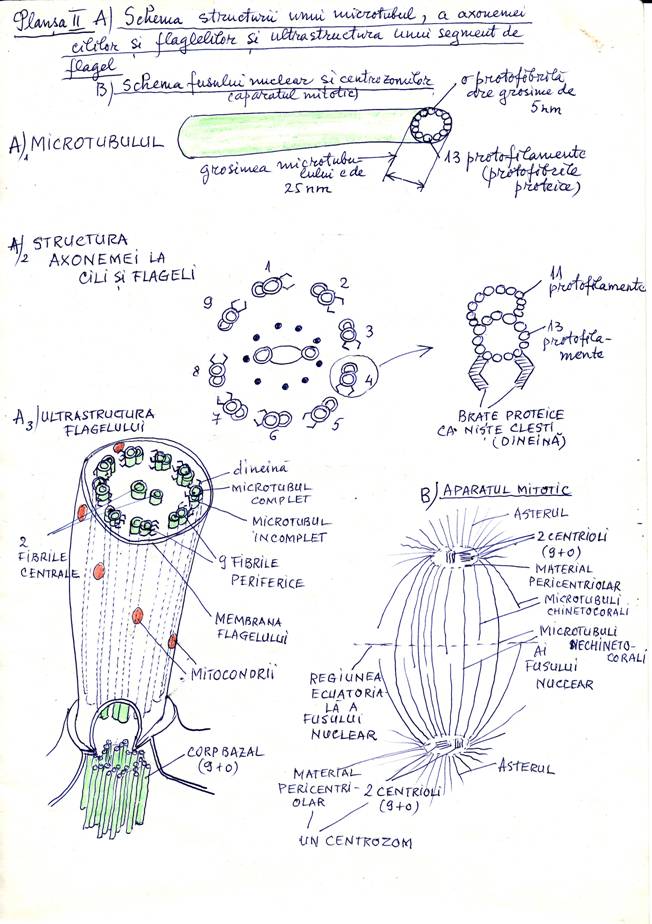
b) Microtubulii sunt cilindrii cu diametrul de 25 nm formati prin aranjarea in cerc a 13 protofilamente sau protofibrile cu diametrul de 5 nm. Aceste protofibrile se formeaza prin polimerizarea unor proteine numite tubuline. Ca si la actine polimerizarea se accelereaza in prezenta unor nucleotide dar aici intervine GTP (guanozintrifosfatul) in loc de ATP (adenozintrifosfatul). La temperatura de 37 grade Celsius polimerizarea are loc spontan iar la 0 grade Celsius are loc procesul invers de depolimerizare. Pe langa temperatura, polimerizarea aceasta e influentata de prezenta unor proteine speciale specifice si a ionilor de Ca 2+ si Mg 2+ .In concentratie mica, Ca 2+ produce polimerizarea iar daca se mareste concentratia 1 mM (milimol) se produce depolimerizarea microtubulilor. Blocarea polimerizarii se face cu substante ca vinblastina si vincristina, folosite in tratamentul cancerului pentru ca distrug microtubulii fusului de diviziune si omoara celulele canceroase care au proprietatea de a se divide rapid. Microtubulii se pot gasi liberi in citoplasma sau formeaza ansambluri de microtubule la cili, flageli in centrul celular si aparatul mitotic (fusul de diviziune si asterul) vezi plansa II.
Microtubulii au doua roluri importante; primul lor rol este structural deoarece ei determina forma celulei si a prelungirilor celulelor (axoni si dendrite la neuroni, cili si flageli), ei determina orientarea si distributia filamentelor intermediare si a filamentelor de actina. Asadar ei sunt un fel de "organizatori ai citoscheletului". Microtubulii ofera o orientare a dispunerii mitocondriilor ce sunt asezate in spirala in spatial cilindric al flagelului de la spermatozoid. Al doilea rol al microtubulilor este cel dinamic, deoarece microtubulii sunt implicati in miscarile celulare ce au la baza mecanismul microtubul-dineina. Acest mecanism microtubul-dineina contribuie la miscarile cililor, miscarea flagelului, deplasarea ordonata a cromozomilor in timpul diviziunilor celulare.
Cilii si flagelii sunt prelungiri permanente ale suprafetei celulare ce prezinta asemanari si deosebiri. Celulele mucoasei respiratorii de la om au numerosi cili dispusi ca un lan de grau iar la spermatozoidul omului si la numeroase specii animale exista un singur flagel
Singura celula flagelata la om este spermatozoidul.
Miscarea cililor (dispusi adesea foarte ordonat in siruri longitudinale, de exemplu la parameci) este facuta la toti cilii dintr-un sir deodata in doi timpi: bataia active, urmata de revenirea lenta la pozitia initiala, urmand apoi miscarea in doi timpi a sirului de cili urmator - asa numita miscare metacrona. Flagelul executa o miscare "in melc" sau helicoidala prin care se imping si inoata inainte celulele flagellate in mediul apos. Miscarea cililor de pe suprafata celulelor se face deci ordonat formand "unde sau valuri battue de vant".Astfel, materiale ca mucusul din mediul extern inferior al cailor respiratorii este impins din plaman spre faringe ducand odata cu el praful, microbii si alte particule materiale. Patrunse prin respiratie. Cilii celulelor mucoasei trompei uterine imping tot in acelasi mod oul spre uter.
Cilul are trei parti: o parte libera care este in afara de corpul celular si in jos mai are doua parti aflate in citoplasma: corpul bazal si radacinile. In partea libera cilul contine un complex de microtubuli numit axonema acoperit circular cu o pelicula subtire de citoplasma peste care se afla membrana celulara sau plasmalema. Axonema are acelasi plan de organizare in toate celulele organismelor eucariote (protiste, ciuperci, plante, animale, om) dovedind unitatea lumii vii. Axonema are 9 dublete de microtubuli asezate in jurul a 2 microtubuli izolati, formand asa zisa structura "9+2". Fiecare dublet are un microtubul complet cu 13 protofilamente si de acesta sta alipit un microtubule incomplete ce are doar 10 - 11 protofilamente. Pe microtubulul complet se afla din loc in loc in lungul sau, brate ca niste clesti, alcatuiti din proteina denumita dineina. Flagelul are o structura asemanatoare, microtubulara, de tip "9+2",la axonema dar are in jurul axonemei, 20-24 mitocondrii dispusi helicoidal. Mitocondriile furnizeaza energia mai multa necesara miscarii flagelului scindana ATP (adenozintrifosfatul) care are acumulata energia. Scindarea ATP se face cu enzima numita ATP-aza. Mecanismul molecular microtubul - dineina este acelasi la cili si flageli. "Ghearele" de dineina de pe un dublet de microtubul pot stabili punti pe dubletul vecin de microtubuli. Dubletele de microtubuli aluneca unul fata de altul indoind axonema si in acest timp se desfac puntile dineina microtubul de la un nivel superior sau inferior si se refac la alt nivel inferior sau respective superior. Oamenii la care lipseste dineina sunt bolnavi de sindromul cililor imobili (boala genetica descrisa de curand) si in care apar infectii respiratorii repetate pentru ca nu se pot curati caile respiratorii iar la barbati se produce sterilitatea pentru ca spermatozoizii devin imobili. Corpii bazali ai cililor si flagelilor au o structura identica cu centriolii ce intra in componenta centrului celular sau centrozomului. Corpii bazali au 9 triplete de microtubuli aranjate circular ca un fel de cilindru, gol la mijloc care poarta structura microtubulara "9+0". Corpii bazali coordoneaza miscarile cailor si flagelilor precum si polimerizarea tubulinei din axoneme. Centrul celular sau centrozomul contine 2 centrioli cu o asezare perpendiculara unul fata de altul, sunt inconjurati de un nor sau halou de citoplasma numit material pericentriolar.
In interfaza sau perioada dintre diviziunile celulare, centrozomul se afla in apropierea nucleului si de la el pleaca in toate directiile microtubule liberi in toata citoplasma. Se crede ca acest material pericentriolar ar regla polimerizarea microtubulilor citoplasmei. Inaintea inceperii diviziunii celulare centriolii sunt dublati si exista deci doua perechi de centrioli. Cele doua perechi de centrioli se separa in timpul diviziunii celeulare si se dispun la cei doi poli ai celulei, fiecare pereche de centrioli fiind afundata intr-un centrozom. Centrozomii au fibre radiare ce fac asa numitul aster, iar intre cei doi centrozomi fibrele microtubulare fac fusul de diviziune. Asterii si fusul de diviziune formeaza aparatul mitotic al celulei. Fibrele fusului fac deplasarea ordonata a cromozomilr spre polii celulei in timpul diviziunii celulare. Toata citoplasma unei celule vii strabatuta in toate directiile de filamente proteice intretaiate ce inlesnesc forma celulei si miscarile celulei vii. Acesta este deci citoscheletul format din: microfilamente, filamente intermediare si microtubuli. Citoscheletul celulei are rol structural si de miscare a celulei eucariote. El este foarte, foarte dynamic deoarece se compune si se destrama dupa cum sunt nevoile de viata si adaptare ale celulei. Microfilamentele si microtubulii se polimerizeaza si se depolimerizeaza facand intre cele doua procese contrare un fel de echilibru permanent. Intre microtubuli si microfilamente pe de o parte si monomerii de actine si tubuline din citosol este alt echilibru permanent. Nucleotidele ciclice (CAMP) =adenozinmonofosfatciclic favorizeaza polimerizarea actinelor si tubulinelor, alti factori sunt specifici pentru microtubuli, pentru microfilamente, iar altii regleaza echilibrul polimerizare - depolimerizare al ambelor diferentieri structurale. Hematia sau globula rosie din sange are forma de disc biconcav din cauza proteinei numita spectrina care formeaza cu actina o retea dispusa sub membrane celulara. Chiar daca plasmalema sau membrane celulara e distrusa, "scheletul" de spectrina - actina al hematiei ii mentine forma.
Citoscheletul interactioneaza cu organitele celulare mentinandu-le in anumite pozitii sau inlesnind deplasarile lor, uneori foarte rapide. Mitocondriile, lizozomii sar intr-o directie si apoi revin la loc sau se aseaza in alte parti ale celulei. Celula eucariota spre deosebire de celula procariota are o miscare a componentelor citoplasmei, in jurul nucleului, pe care nu-l pot destrama din cauza membranei nucleare duble. Este o miscare numita cicloza, prevazuta cu curenti citoplasmatici deosebiti de miscarea dezordonata browniana a moleculelor dizolvate intr-o solutie. Un curent citoplasmatic este transportul axoplamic din lungul axonului, de la corpul neuronului spre sinapsa. Se deplaseaza astfel organite celulare, vezicule sinaptice, proteine si hormoni neurosecretati ce sunt transportati prin citosol (citoplasma). Exista un microscop electronic cu voltaj suprainalt (in care electronii sunt accelerate de 10 ori mai mult ca la microscopul electronic "obisnuit") si cu care savantul Keith Porter a descoperit in citoplasma o retea foarte fina de filamente ce mentin organitele si microtubulii in anumite pozitii si care au fost numite microtrabecule. Acestea apara celula de variatiile continutului de apa, de deplasarea granulelor de pigmenti, iar unele enzime s-ar agata de microtrabecule.
Copyright © 2024 - Toate drepturile rezervate