|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Hibridizarea acizilor nucleici reprezinta procesul prin care 2 monocatene ADN sau ARN din surse biologice diferite reformeaza configuratia dublu catenara (Fig.3.44.). Din punct de vedere chimic, procesul de hibridizare se bazeaza pe :
principiul complementaritatii dintre cele 2 catene ale unui duplex ADN sau ARN
eventuala omologie de secventa dintre cele 2 surse ADN
Pe aceleasi principii se pot forma hibrizi ADN : ADN, hibrizi ARN : ARN sau hibrizi ADN : ARN.
In majoritatea cazurilor, scopul tehnicilor de hibridizare este identificarea sau localizarea anumitor fragmente de acizi nucleici.
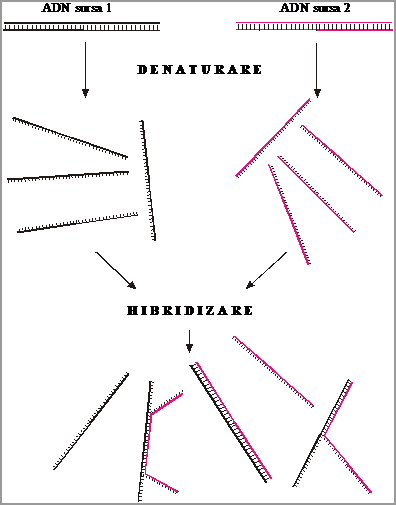
In procesele de hibridizare a acizilor nucleici se definesc notiunile :
molecula TINTA = fragmentul de ADN, ARN sau de proteina de identificat sau de localizat;
molecula SONDA = fragmentul de ADN,ARN sau de proteina cu ajutorul caruia, prin hibridizare, este identificata sau localizata TINTA
In foarte multe variante tehnice, hibridizarea acizilor nucleici se realizeaza pe suport solid (presupunand transferul moleculelor de acizi nucleici pe suportul solid), purtand denumirea de BLOTTING. Exista 3 categorii principale de blotting :
SOUTHERN BLOTTING (dupa numele autorului - Southern, 1975) - tehnica moleculara ce identifica fragmente de ADN folosind sonde ADN sau ARN
NORTHERN BLOTTING - tehnica moleculara ce identifica fragmente ARN folosind sonde ARN sau ADN
WESTERN BLOTTING - tehnica moleculara ce identifica fragmente proteice folosind anticorpi specifici
2. Caracteristicile generale ale sondelor
O sonda moleculara de acizi nucleici reprezinta o secventa de nucleotide, scurta, monocatenara, alcatuita din ADN sau din ARN, ce se va lega specific de o secventa tinta; cu ajutorul sondei va fi identificata sau localizata molecula tinta. Gradul de omologie de secventa dintre sonda si tinta va determina stabilitatea hibridului. Sondele moleculare de acizi nucleici se construiesc astfel incat sa prezinte urmatoarele caracterisitici:
a) sa prezinte un marcaj chimic care sa permita vizualizarea hibridului molecular sonda:tinta
b) sa aiba dimensiune cuprinsa intre 10 si 10.000 nucleotide; de regula insa, se folosesc sonde de 14 - 40 nucleotide. Sondele prea scurte hibridizeaza foarte rapid (in cateva minute), dar prezinta si hibridizari nespecifice si sunt si mai greu de marcat; sondele prea lungi hibridizeaza foarte incet (in cateva ore), dar hibrizii sunt mai stabili si mai specifici
c) specificitatea de reactie a sondei este data de secventa acesteia; sonda nu trebuie sa hibridizeze cu ea insasi, deci sa nu contina complementaritate intracatenara; totodata, sonda trebuie sa nu hibridizeze cu non-tinte
3. Modalitati de sinteza a sondelor moleculare de acizi nucleici
Sondele moleculare de acizi nucleici pot fi marcate uniform (prin nick-translation, primeri randomizati, ADNc total, transcriere in vitro) sau doar la capete.
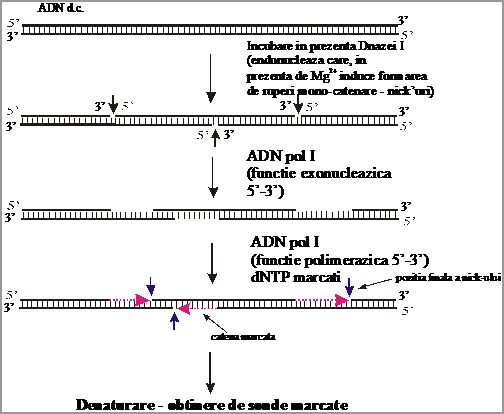
3.1. Sinteza de sonde prin nick-translation
In aceasta tehnica se porneste de la o molecula de ADN d.c., de obicei o copie a moleculei tinta :
a) in prima etapa, aceasta molecula este supusa unei digestii cu DNaza I, care este o endonucleaza situs-nespecifica, ce in prezenta ionilor de Mg++ produce ruperi monocatenare randomizate (denumite "nicks");
b) in a 2-a etapa, actioneaza ADN polimeraza I de E.coli, care prin activitatea sa exonucleazica 5'-3' largeste ruperile monocatenare;
c) aceasta activitate are loc aproape simultan cu activitatea de polimerizare a ADN polimerazei I (in directie 5'-3') folosind molecule dNTP marcate. Ceea ce se obtine este o deplasare a ruperii monocatenare, de unde si denumirea de "nick-translation" ;
d) ultima etapa este reprezentata de denaturarea termica a moleculelor, pentru obtinerea unor monocatene marcate uniform.
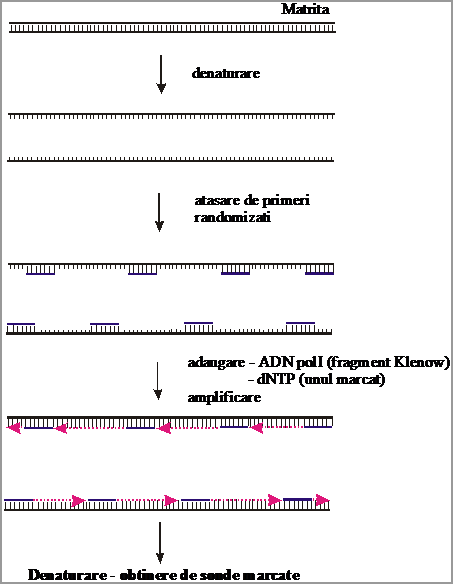
3.2. Sinteza de sonde marcate uniform folosind primeri randomizati
In aceasta varianta tehnica se folosesc ca primeri oligonucleotide cu secventa heterogena, astfel incat ei vor hibridiza in multe pozitii ale matritei. Asemenea primeri poarta denumirea de primeri randomizati si pot fi fie cumparati, fie obtinuti astfel:
a) prin digestia ADN din sperma de somon sau din timus de vitel cu DNaza I, astfel incat se obtine o populatie mare de fragmente de ADN m.c., cu dimensiuni intre 6 si 12 nucleotide;
b) prin sinteza cu ajutorul unui sintetizator automat de oligonucleotide (octameri ce contin toate cele 4 baze azotate in toate pozitiile);
In ceea ce priveste enzimele utilizate, in aceasta tehnica se folosesc o ADN polimeraza ADN-dependenta (revers-transcriptaza) si Klenow-ADN pol I (care nu are activitate exonucleazica 5'-3'). Pentru a obtine sonde marcate se recomanda utilizarea, in amestecul de dNTP, a unui nucleotid marcat si 3 nemarcati.
3.3. Sinteza de sonde ADNc total
Pentru obtinerea unei populatii de molecule de ADNc (complementar cu o populatie de ARN m.c.) se pot folosi urmatoarele tehnici:
a) intr-o prima varianta tehnica, se izoleaza ARNm total din celule si se desfac structurile secundare ale acestor molecule prin incalzire 5 min la 70oC. Se ataseaza apoi primeri randomizati (de ex. octameri sintetici). Pornind de la acestia va fi initiata reactie de polimerizare a dNTP-urilor introduse in reactie cu ajutorul unei revers-transcriptaze (rTth sau revers-transcriptaze din MLV (Murine Leukaemia Virus) sau din AMV (Avian Mieloblastosis Virus). Intr-o etapa urmatoare, pentru a obtine denaturarea moleculelor hibride, amestecul de reactie se trateaza cu NaOH la peste 70oC, iar rezultatul final este reprezentat de o populatie de molecule scurte de ADNc "partiale".
b) intr-o a doua varianta tehnica, dupa izolarea ARNm total si desfacerea structurilor secundare, se folosesc primeri oligo (dT) care se ataseaza la cozile poli(A) de la capatul 3' al majoritatii moleculelor ARNm mamaliene. Si in aceasta situatie se foloseste tot o revers-transcriptaza, urmata de tratament cu NaOH la 70oC. Spre deosebire de prima varianta tehnica, aici rezultatul este reprezentat de o populatie de molecule ADNc intregi.
Sinteza de sonde ARN prin transcriere in vitro
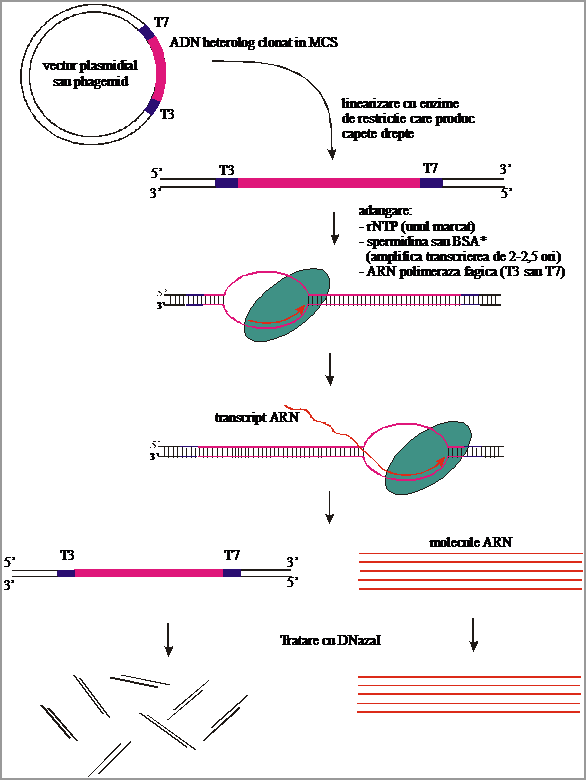
Pentru sinteza de sonde ARN prin transcriere in vitro este recomandabil ca ADN heterolog sa fie in prealabil clonat intr-un vector plasmidial sau intr-un fagimid, care sa contina promotori puternici, asa cum sunt promotorii din fagii T3 si, respectiv, T7. Acesti promotori au fost alesi pentru ca sunt recunoscuti extrem de specific de ARN polimeraza din fagii respectivi. Astfel, enzima are o specificitate distincta pentru promotorul cognat si nu foloseste promotorii recunoscuti de alte ARN polimeraze (aceste doua ARN polimeraze nu recunosc nici promotori bacterieni cromozomali sau plasmidiali si nici promotori de la eucariote).
Principalele etape ale acestei tehnici sunt (Fig.3.45.):
a) vectorul recombinat (ce contine si ADN heterolog) este linearizat cu ajutorul unor endonucleaze de restrictie ce produc capete netede si care taie in afara celor 2 promotori (de T3 si T7); de ex., in cazul clonarii unei secvente de ADN in vectorul pBluescript SK+, intreaga caseta se scoate prin digestie cu endonucleaza de restrictie BssH II;
b) fragmentul ADN linear ce contine ADN heterolog marginit de cei 2 promotori se introduce in amestec cu ARN polimeraza de fag T3 sau de fag T7 (functie de catena ce urmeaza a fi transcrisa), spermidina sau albumina serica bovina (ce activeaza transcrierea de 2 - 2.5 ori) si rNTP (dintre care unul marcat radioactiv sau neradioactiv); intreg amestecul de reactie se tine 1 - 2 h la 37oC, timp in care are loc un proces de transcriere in vitro, ce conduce la acumularea de molecule ARN marcate;
c) in ultima etapa, moleculele ADN folosite ca matrita sunt transformate in fragmente foarte mici prin tratatment cu DNaza I, dupa care moleculele de ARN sunt precipitate cu etanol 100% in prezenta de acetat de amoniu 2.5 M.
a) Marcarea capetelor 3' ale ADN d.c.
Pentru a realiza marcarea radioactiva a unor molecule de ADN d.c., intr-o prima etapa se folosesc endonucleaze de restrictie, fie ce produc capete coezive, fie ce produc capete netede:
- in cazul digestiei ADN cu enzime de restrictie ce produc capete coezive, ADN-ul restrictat se amesteca apoi cu un dNTP marcat (functie de enzima cu care a fost in prealabil digerat ADN) si cu Klenow-ADN polimeraza I de E.coli (aceasta enzima va adauga dNTP-ul introdus in reactie pe baza de complementaritate); de ex., in cazul digestiei ADN cu EcoR I, se adauga P -dATP, iar Klenow-ADN polimeraza I va adauga resturi de adenine marcate complementare cu cele 2 resturi de timine (Fig.3.46.);

- in cazul digestiei ADN cu enzime de restrictie ce produc capete netede, acestea pot fi marcate prin inlocuirea nucleotidei de la capul 3'OH- in amestecul de reactie se introduce nucleotida de inlocuit ca P -dNTP;
- in cazul marcarii capetelor 3' monocatenare, se foloseste enzima terminal-deoxinucleotidil transferaza din timus de vitel; aceasta enzima este o ADN polimeraza "neobisnuita", gasita numai in prelimfocite si in stadii timpurii ale diferentierii limfoide; in prezenta unui cation divalent, enzima produce aditia de dNTP-uri la capetele 3'OH, preferabil monocatenare: in prezenta de Mg++, se aditioneaza o purina, iar in prezenta de Co++, se aditioneaza o pirimidina (Fig.3.47.)
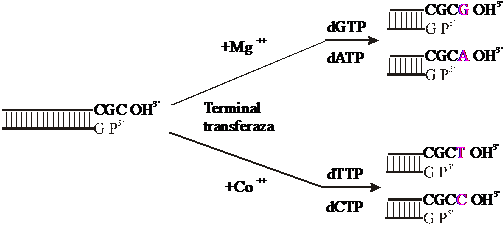
Fig.3.47. Marcarea capetelor 3' monocatenare
Marcare radioactiva In marea majoritate a cazurilor, in marcarea radioactiva a moleculelor de acizi nucleici se folosesc P sau S . Avantajul marcarii radioactive a acizilor nucleici il reprezinta faptul ca au o sensibilitate foarte mare, putand detecta cantitati foarte mici de acizi nucleici. Principalele deazavantaje ale marcarii radioactive sunt reprezentate de riscul manipularii unor materiale radioactive, de instabilitatea isotopilor, precum si de timpii lungi de expunere.
Marcare neradioactiva
In marcarea neradioactiva se folosesc molecule reporter ce sunt detectabile direct sau indirect.
2.a. metode directe de marcare neradioactiva : molecula reporter este detectabila direct si se leaga direct de sonda, astfel ca hibrizii pot fi vizualizati direct; in asemenea situatii, moleculele reporter sunt de obicei reprezentate de fluorocromi.
In cazul marcarii fluorescente a sondelor de acizi nucleici se folosesc analogi ai nucleotidelor cuplati cu fluoresceina (de ex. fluorescein-dUTP), ce se incorporeaza in acizii nucleici prin tehnici enzimatice standard (de ex. prin reactie PCR). Marcajul cu fluoresceina poate fi folosit si ca metoda indirecta, caz in care fluoresceina este folosita ca haptena.
2.b. metode indirecte de marcare radioactiva: sonda contine o molecula reporter introdusa chimic sau enzimatic, care va deveni detectabila prin citochimie de afinitate; in aceste situatii, se foloseste, fie biotina cuplata cu avidina sau cu streptavidina, fie digoxigenina cuplata cu anticorpi-antidigoxigenina.
Biotina (sau vitamina H) poate fi incorporata in acizii nucleici ca biotin-dUTP (Fig.) prin tehnici enzimatice standard (de ex. PCR). Dupa folosirea sondei biotinilate intr-o reactie de hibridizare moleculara, hibrizii de acizi nucleici sunt detectati prin cuplarea biotinei cu avidina din complexul avidina-fosfataza alcalina (sau avidina-peroxidaza din hrean).
Avidina este o glicoproteina din albusul de ou, care are 4 subunitati, fiecare continand un situs de legare cu biotina (in utlimii ani, in loc de avidina se foloseste streptavidina, care nu contine reziduuri glicozilate ce dau un fond ridicat al reactiilor nespecifice, reducand contrastul de culoare)
Enzima va degrada un compus cromogen, obtinandu-se in final o reactie de culoare. Astfel, fosfataza alcalina va degrada substratul cromogen NBT (nitro-blue-tetrazolium) sau BCIP (5-Br-4-Cl-3-indolilfosfat), obtinandu-se un precipitat de culoare albastra. In locul fosfatazei alcaline se poate folosi peroxidaza din hrean (HPOX), care va degrada substratul cromogen DAB (3,3'-diaminobenzidina tertahidroclorid), obtinandu-se un precipitat rosu-brun.
O alta varianta de marcare din acelasi sistem, foloseste digoxigenina in loc de biotina. In acest caz, in sonda este incorporat enzimatic, fie digoxigenina-dUTP, fie digoxigenina-dTTP. Dupa realizarea reactiei de hibridizare moleculara, se adauga anticorpi-anti-digoxigenina cuplati cu fosfataza alcalina si apoi substratul cromogen (NBT sau BCIP), iar hibrizii sunt detectati ca spot/banda albastra.
1. Sinteza sondei marcate Functie de molecula de ADN tinta, de lungimea sondei dorite, precum si de tipul de marcare, se sintetizeaza (prin tehnicile prezentate mai sus) o specie moleculara de sonda ADN sau ARN complementara cu molecula tinta.
2. Procesarea ADN tinta Moleculele de ADN sursa in care se gaseste si secventa de ADN tinta, trebuie izolate si purificate. Ulterior, ADN sursa se digera cu o endonucleaza de restrictie, astfel incat secventa de ADN tinta sa fie obtinuta ca molecula d.c. lineara.
Amestecul de digestie se supune unei electroforeze in gel de agaroza, in care se separa diversele fragmente obtinute in reactia de restrictie.
Denaturarea ADN tinta Gelul de agaroza in care se afla si moleculele de ADN se introduce in solutii de NaOH, in care va avea loc denaturarea acestora, rezultand molecule monocatenare.
4. Transferul ADN tinta pe suport solid Fragmentele de ADN d.c. lineare separate in gelul de electroforeza se transfera pe un suport solid (hartie de nitroceluloza sau membrana de nylon), dupa ce in prealabil se realizeaza o denaturare a moleculelor de ADN prin imersarea gelului in NaOH 0.5N. Transferul poate fi realizat prin 3 variante tehnice:
- transfer prin capilaritate, in sistem sandwich: fragmentele de ADN sunt transportate din gel de catre un lichid (de obicei, tampon SSC) si depozitate pe suportul solid; miscarea lichidului se realizeaza prin capilaritate si se datoreaza unui teanc de hartie prosop asezat in sistem sandwich; rata transferului depinde de dimensiunea fragmentelor si de concentratia agarozei.
- transfer electroforetic: se foloseste numai pe membrane de nylon incarcate electric si reprezinta o metoda eficienta chiar pentru transferul unor fragmente mici: 56 bp.
- transfer prin vacuum: este mult mai eficient si mai rapid decat celelalte 2 variante tehnice; in acest caz, gelul de agaroza se gaseste in contact direct cu suportul solid (nitroceluloza sau membrana de nylon), amandoua asezate pe un suport poros deasupra unei camere de vacuum; tamponul este asezat in camera superioara a dispozitivului si este tras prin vacuum, eluand acizii nucleici din gel si depozitandu-i pe suportul solid.
Suportul solid cu acizii nucleici depozitati pe el este apoi expus la radiatii UV, ceea ce creeaza o serie de legaturi de tip cross-links intre moleculele de acizi nucleici si structura suportului; astfel, fragmentele de acizi nucleici (inclusiv ADN tinta) sunt fixate de suportul solid.
5. Hibridizarea moleculara In aceasta etapa, suportul solid pe care se afla fixata tinta se imerseaza in tampon 5xSSC care contine si sonda marcata, si se lasa in agitare usoara 14-16h. Dupa hibridizare, sonda nehibridizata se indeparteaza prin spalari succesive cu tampon SSC.
Copyright © 2025 - Toate drepturile rezervate