|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Efectele biologice si mecanismele moleculare de actiune ale radiatiilor UV
In ultimii ani s-a constatat ca incidenta cancerului cutanat a crescut in mod alarmant (Shore, 2001). Aceasta observatie a dirijat interesul specialistilor spre investigarea efectelor biologice ale radiatiilor UV solare. Este cunoscut faptul ca expunerea la radiatii UV determina efecte imediate (eritem si pigmentare) si efecte intarziate (imbatranirea prematura a pielii si diferite forme de cancer) (Robert, 1999).
Spectrul UV al luminii solare cuprinde domeniile: UVC (190-290 nm), UVB (290-320 nm) si UVA (320-380 nm). Intrucat stratul de ozon al stratosferei retine in totalitate radiatiile UVC si partial radiatiile UVB, la suprafata Pamantului ajung numai radiatii UVA, si in mica masura radiatii UVB (Huda, 2000).
Radiatiile din domeniul UVA si UVB actioneaza prin mecanisme diferite. Radiatiile UVB au o energie mai mare comparativ cu radiatiile UVA si sunt absorbite direct de o serie de constituenti celulari, ca de exemplu acizii nucleici, proteinele si acidul urocanic. Radiatiile UVB exercita un efect mutagen, fiind principalul agent responsabil de aparitia cancerului cutanat (de Gruijl, 2000).
Fotonii UVA au o energie mai mica decat fotonii UVB, dar au o pondere de peste 90% din energia solara din spectrul UV de la suprafata Pamantului. In plus, radiatiile UVA penetreaza cu usurinta epiderma, actionand asupra stratului proliferativ bazal al epidermei si chiar asupra componentelor sanguine din derma (Leszczynski si colab., 1996). Radiatiile UVA actioneaza asupra constituentilor celulari in mod indirect, prin mecanisme oxidative ce implica formarea speciilor reactive de oxigen (anioni superoxid, peroxid de hidrogen, radicali hidroxil, radicali peroxil, si oxigen singlet). Speciile reactive de oxigen au o durata de viata relativ scurta, dar sunt extrem de reactive, reactionand cu cea mai mare parte a constituentilor celulari: acizi nucleici, proteine, lipide, polizaharide. Actiunea lor induce de cele mai multe ori modificari ireversibile. Radiatiile UVA exercita un efect mutagen mai slab decat radiatiile UVB, dar joaca un rol important in etiologia cancerului de piele, avand in vedere predominanta lor in spectrul UV la suprafata Pamantului (Stary si colab., 1997).
Desi in cea mai mare parte radiatiile UV solare exercita efecte nocive, nu poate fi neglijat efectul lor benefic legat de sinteza vitaminei D, precum si efectul psihologic pozitiv al expunerii la soare.
1. Efectele radiatiilor UV solare asupra organismului uman
Efectele radiatiilor UV la nivelul pielii
Pielea este organul cel mai expus la radiatiile solare. Pielea este alcatuita din epiderma, derma si tesut subepidermal. In structura epidermei se evidentiaza (1) stratul bazal (stratum germinativum), (2) stratul spinos (stratum spinosum), (3) stratul granular (stratum granulosum) si (4) stratul cornos (stratum corneum). Epiderma este separata de derma printr-o membrana bazala. Derma are o grosime de circa 1000-4000 μm si are in structura sa fibre elastice si de colagen produse de fibroblaste, vase de sange si limfatice, foliculi pilosi, glande sebacee si sudoripare, fibre musculare netede si nervi. Tesutul subepidermal este un tesut adipos.
Expunerea organismului uman la radiatii solare din regiunea 290-315 nm este esentiala pentru formarea vitaminei D3 in piele, cu consecinte benefice asupra dezvoltarii sistemului osos.
Studiile in vivo au aratat ca transmisia prin epiderma umana a radiatiilor cu lungimi de unda de 313, 365 si 436 nm se face in proportie de 9,5, 19 si respectiv 34% din radiatia incidenta (Kohen si colab., 1995). Efectele imediate dupa expunerea la soare, observate la nivelul tegumentului, sunt aparitia eritemului si pigmentarea. Eritemul este o consecinta a dilatarii vaselor de sange din derma. Acest efect este mediat de produsii de reactie ai acidului arahidonic formati sub actiunea ciclo-oxigenazei si lipoxigenazei. Generarea prostaglandinelor, asociata eritemului in primele 6-12 ore de la iradiere, poate fi blocata de agenti anti-inflamatori nesteroidici, ca de exemplu indometacinul. Acesti agenti anti-inflamatori nu sunt eficienti in cazul eritemului inregistrat dupa 24 de ore de la iradiere si care este modulat de produsii lipoxigenazei.
Mai mult de 80% din radiatia UVA este transmisa prin stratul superficial cornos al pielii si determina efectul de pigmentare imediata (Kuusilehto, 2000). Bronzarea pielii implica doua procese separate: pigmentarea imediata si pigmentarea intarziata (melanogeneza). Pigmentarea imediata este determinata de radiatiile UVA si consta in foto-oxidarea melaninei preformate. Pigmentarea intarziata are loc la 72 de ore dupa expunerea la radiatii UVA sau UVB si este asociata cu melanogeneza. In cursul melanogenezei se constata o crestere de 2-3 ori a numarului de melanocite DOPA-pozitive si o dublare a volumului corpului celular al melanocitelor. Radiatiile UV amplifica transferul melanozomilor de la melanocite la keratinocite.
Radiatiile UV pot determina, pe langa efectul de pigmentare, imbatranirea prematura a pielii numita fotosenescenta actinica. Cele mai afectate sunt tegumentele expuse cel mai mult la radiatiile solare. Alterarea structurii cutanate este insotita de deshidratare si reducerea elasticitatii, insa cea mai grava consecinta a expunerii la radiatiile UV consta in aparitia si dezvoltarea unor tumori, cu evolutie posibil letala. Tumorile cutanate pot fi de tip melanom (denumirea provine de la faptul ca au originea la nivelul melanocitelor, celulele producatoare de pigment), sau tip carcinom, numite si cancere tip non-melanom, ce au ca origine keratinocitele (Wang si colab., 2001). Cele mai frecvente forme de carcinom cutanat, pentru care s-a stabilit o relatie de cauza-efect in raport cu expunerea la radiatii UV, sunt carcinoamele de tip bazocelular si squamos (van der Leun, 1996). Keratoza actinica este o stare precanceroasa determinata de expunerea la radiatii UV solare care poate evolua spre o forma de cancer de piele.
Efectele radiatiilor UV asupra sistemului imunitar
Expunerea la radiatiile UV determina o scadere a raspunsului imun (imunosupresie), ce are drept consecinta reducerea capacitatii de aparare fata de infectii si fata de dezvoltarea tumorilor (Halliday si colab., 1998). La persoanele infectate cu HIV, expunerea la radiatii UV amplifica imunosupresia deja existenta. Trebuie subliniat faptul ca pigmentarea pielii nu ofera protectie fata de actiunea de imunosupresie exercitata de radiatiile UV. Exista totusi cateva situatii in care supresia sistemului imunitar prin iradiere poate fi benefica. Astfel, acest efect este exploatat in terapia unor boli de piele, ca de exemplu psoriazis vulgaris.
Modificarile imunologice sunt determinate de actiunea radiatiilor UV asupra celulelor Langerhans. Actiunea se exercita atat direct, cat si indirect, prin intermediul factorilor solubili eliberati in urma lezarii keratinocitelor.
Acidul urocanic (acid 4-imidazol acrilic) este prezent in stratul cornos al pielii. In mod normal, acidul urocanic se afla in forma trans, dar sub efectul iradierii UV sufera izomerizarea la forma cis. Se considera ca acidul cis-urocanic este unul dintre initiatorii procesului de imunosupresie asociat aparitiei tumorilor non-melanom.
Efectele radiatiilor UV la nivel ocular
Dintre efectele negative exercitate de radiatiile solare UV trebuie mentionate o serie de modificari la nivel ocular, cum sunt degenerarea maculara si cataracta (Balasubramanian, 2000). Aceste modificari sunt corelate in principal cu inaintarea in varsta, dar studiile experimentale au dovedit ca si radiatiile UV au un rol important in instalarea si evolutia acestor modificari oculare (Zigman si colab., 1995; Zigman, 2000). Radiatiile UV amplifica oxidarea proteinelor din cristalin (in special a resturilor de Met si Cys) si induc formarea unor agregate proteice cu masa moleculara mare (Finley si colab., 1997). Aceste modificari determina aparitia unor zone opace la nivelul cristalinului corelate cu dezvoltarea cataractei.
Factorii care determina susceptibilitatea la iradierea UV
Susceptibilitatea de a suferi leziuni in urma iradierii UV este conditionata de factorii genetici, de anumite maladii, de factori corelati cu varsta, precum si de administrarea unor medicamente si substante chimice. In functie de sensibilitatea de aparitie a arsurilor si de gradul de pigmentare exista sase tipuri de piele (tipul I corespunde celei mai mari sensibilitati - indivizii sufera intotdeauna arsuri si nu se bronzeaza niciodata; tipul VI corespunde celei mai bune protectii - pielea este pigmentata si nu sufera niciodata arsuri). Exista o serie de maladii ereditare care determina o susceptibilitate marcanta la radiatile UV, dintre care pot fi mentionate: xeroderma pigmentosum, sindromul Bloom, sindromul Rothmund-Thomson, porfiriile, si fenilcetonuria. De asemenea, cresterea susceptibilitatii la leziunile determinate de radiatiile UV poate fi asociata administrarii antibioticelor pe cale orala, medicamentelor antihipertensive, psoralenului, agentilor imunosupresori si anti-inflamatori nesteroidici.
2. Mecanismele de actiune ale radiatiilor UVA
Radiatiile UVA actioneaza asupra mediilor biologice prin mecanisme oxidative, corelate cu formarea speciilor reactive de oxigen: oxigen singlet, radicali hidroxil (OH.), anioni superoxid (-O2), peroxid de hidrogen (H2O2). Speciile reactive de oxigen se impart in: a) specii cu caracter de radical liber: anionul superoxid, radicalul hidroxil, oxidul nitric; b) specii neradicalice: oxigenul singlet, peroxidul de hidrogen, acidul hipocloros, hidroperoxizii lipidici. Desi termenul de specii reactive de oxigen este cel mai adecvat, in literatura biochimica si medicala se foloseste termenul de radicali liberi ai oxigenului, in care sunt incluse toate speciile respective, indiferent daca au sau nu caracter de radical liber.
2.1. Formarea speciilor reactive de oxigen
Radiatiile UVA sunt absorbite slab de acizi nucleici si proteine, dar evenimentul initial in declansarea efectelor biologice consta in absorbtia fotonilor UVA de catre diferiti cromofori din mediul celular: chinone, steroizi, porfirine, flavin coenzime si proteine cu grupari hem (citocromi, peroxidaze, catalaza). Absorbtia fotonilor UVA de catre cromofori are ca rezultat formarea speciilor reactive de oxigen sau transferul energiei acumulate catre anumite molecule tinta.
Peroxidul de hidrogen se formeaza prin iradierea UVA a triptofanului, iar anionii superoxid pot fi produsi prin iradierea UVA a NADH si NADPH (Cunningham si colab., 1985). In prezenta ionilor de fier, peroxidul de hidrogen si anionii superoxid pot participa in vivo la reactia Haber-Weiss soldata cu producerea radicalilor hidroxil:
Fe2+
+ H2O2 → Fe3+ +
Fe3+ + -O2 → Fe2+ + O2
Actiunea UVA asupra constituentilor celulari
O mare parte a constituentilor celulari reprezinta tinte ale atacului speciilor reactive de oxigen generate de iradierea UVA. Astfel, radicalii hidroxil reactioneaza cu aproape toate tipurile de molecule din componenta celulelor: glucide, fosfolipide, nucleotide, acizi organici si aminoacizi (Kohen si colab., 1995). In tabelul 1 sunt prezentate constantele de viteza ale reactiilor radicalilor hidroxil cu diverse molecule din sistemele biologice.
Tabelul 1. Constantele de viteza ale reactiilor radicalilor OH. cu diverse
molecule din sistemele biologice.
|
Compusul testat |
Valoarea pH |
|
|
Adenina |
7,4 |
3,0 x 109 |
|
AMP |
5,4 |
1,8 x 109 |
|
Arginina |
7,0 |
2,1 x 109 |
|
Cisteina |
1,0 |
7,9 x 109 |
|
Citozina |
7,0 |
2,9 x 109 |
|
Glucoza |
7,0 |
1,0 x 109 |
|
Glutation |
1,0 |
8,8 x 109 |
|
Guanina |
- |
1,0 x 1010 |
|
Hemoglobina |
- |
3,6 x 1010 |
|
Histidina |
6-7 |
3,0 x 109 |
|
Metionina |
7,0 |
5,1 x 109 |
|
Timina |
7,0 |
3,1 x 109 |
|
Triptofan |
6,0 |
8,5 x 109 |
|
Uracil |
7,0 |
3,1 x 109 |
Modificarile oxidative ale proteinelor sub actiunea speciilor reactive de oxigen pot determina inactivarea enzimelor si a proteinelor membranare (Dovrat si Weinreb, 1999), si pot produce modificari structurale soldate cu destabilizarea morfologiei celulelor atunci cind tinta atacului oxidativ sunt elementele citoscheletului (Somosy, 2000). In cazul enzimelor, efectul speciilor reactive de oxigen consta, in general, in diminuarea capacitatii catalitice, determinata frecvent de oxidarea gruparilor sulfhidril si modificarea gruparilor amino prin malonilare (Dumitru si Nechifor, 1994; Dean si colab., 1997). Amploarea modificarilor este determinata de localizarea relativa a situsului de formare a speciilor reactive de oxigen, a sistemelor antioxidante si a proteinei tinta.
O tinta majora a atacului radicalic sunt lipidele membranare, datorita prezentei dublelor legaturi din structura acizilor grasi polinesaturati. In structura fosfolipidelor membranare acizii grasi polinesaturati cel mai frecvent intalniti sunt acidul linoleic (C18:2 Δ9,12), acidul linolenic (C 18:3 Δ6,9,12) si acidul arahidonic (C 20:4 Δ5,8,11,14) Radiatiile UVA s-au dovedit initiatori eficienti ai procesului de peroxidare lipidica (Morliere si colab., 1995). Mecanismul peroxidarii acizilor grasi polinesaturati este prezentat in Figura 1.
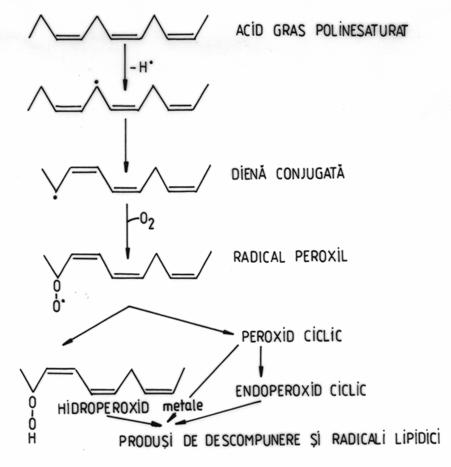
Figura 1. Mecanismul peroxidarii acizilor grasi polinesaturati.
Procesul de peroxidare a
acizilor grasi polinesaturati cuprinde urmatoarele etape: un
radical liber extrage un atom de hidrogen din pozitia alilica, cu
transformarea acidului gras polinesaturat intr-un radical lipidic; urmeaza
rearanjarea intramoleculara a dublelor legaturi cu formarea unei
diene conjugate; in urma reactiei dintre diena conjugata si
oxigenul molecular se formeaza un radical peroxil; radicalul peroxil poate
reactiona cu alta molecula de acid gras polinesaturat, formand
radicali lipidici, cu transformarea radicalului peroxil in hidroperoxid
lipidic. Alternativ, radicalul peroxil poate forma peroxizi ciclici. Peroxizii
lipidici odata formati sufera sciziuni ce duc la propagarea
lantului de reactii, la reactii de ramificare si de descompunere.
Prin descompunerea peroxizilor lipidici se formeaza o mare diversitate de
compusi: alcani (etan, pentan), aldehide (dialdehida malonica,
hexanal, 4-hidroxinonenal), acizi grasi epoxi si hidroxi. Peroxidarea
lipidelor membranare afecteaza structura si functiile membranei
plasmatice si a membranelor organitelor. Astfel, sunt perturbate
potentialele transmembranare, fluxurile ionice si transportul
transmembranar, sunt inactivati receptorii de membrana si sunt
dereglate caile de semnalizare. Prin procesul de peroxidare lipidica
se modifica nu numai componentele membranare de natura lipidica,
ci chiar si proteinele, in urma reactiei unor aminoacizi (Cys,
Intrucat maximul de absorbtie al acizilor nucleici este la 260 nm, radiatiile UVA sunt slab absorbite de acest tip de macromolecule (Kuluncsics, 1999). Se considera ca radiatiile UVA interactioneaza cu DNA prin mecanisme indirecte, mediate de compusii fotosensibilizatori endogeni (Kielbassa si colab., 1997). Faptul ca iradierea celulelor in absenta oxigenului diminueaza considerabil efectul letal al radiatiilor UVA, sugereaza implicarea speciilor reactive de oxigen in interactia UVA-acizi nucleici. Studiile experimentale pe culturi de celule umane au aratat ca radiatiile UVA induc mai multe tipuri de leziuni ale DNA: scindarea legaturilor fosfodiesterice la nivelul unei singure catene, formarea de legaturi incrucisate DNA-proteine, dimerizarea pirimidinelor adiacente. Fotonii UVA determina clivari monocatenare prin scindarea legaturilor fosfodiesterice cu o eficienta foarte mare (de 30 de ori mai mult comparativ cu fotonii UVB si UVC), si acest tip de leziune a fost semnalata intr-o proportie semnificativ mai mare comparativ cu dimerii pirimidinici (Peak si colab., 1987). Repararea clivarilor monocatenare dupa iradierea UVA este un proces extrem de rapid, astfel incat 90% dintre leziuni sunt reparate in numai 15 minute dupa iradiere (Peak si colab., 1991). Formarea dimerilor pirimidinici nu reprezinta o modificare majora a DNA in cursul iradierii UVA, comparativ cu iradierea UVB si UVC.
Radiatiile UVA induc vacuolizarea reticulului endoplasmic, separarea nucleului de citoplasma si formarea corpusculilor Heinz in eritrocite. Formarea corpusculilor Heinz este corelata cu leziuni ale citoscheletului eritrocitar in conditii de stres oxidativ. Studiile dermatologice au indicat ca o iradierea cronica cu UVA determina leziuni ale membranelor mitocondriale din toate tipurile de celule ale dermei.
2.3. Efectul UVA de activare a transcriptiei
Radiatiile UVA activeaza expresia unui set de gene prin mecanisme dependente de redoxul celular. Astfel, chiar la doze reduse care aparent nu determina leziuni tisulare radiatiile UVA determina activarea transcriptionala a genelor ce codifica hem oxigenaza I (HO-1), metalotioneina, CL 100 fosfataza si colagenaza.
Activarea HO-1
reprezinta un mecanism de reglare a nivelului intracelular al ionilor de
fier la valori minime. Un nivel ridicat al ionilor de fier ar promova
reactia Fenton, reactie nociva generatoare de radicali hidroxil
cu efect potential letal la nivel celular (H2O2 + Fe2+
→ Fe3+ +
Radiatiile UVA induc expresia genei ce codifica metalotioneina (Dudek si colab., 1993). Metalotioneina este o proteina cu afinitate mare de legarea a ionilor de zinc, cadmiu si cupru. Metalotioneina este implicata in protectia fata de stresul oxidativ, actionand ca scavenger al speciilor reactive de oxigen. Astfel, metalotioneina poate capta radicalii hidroxil si superoxid si o serie de radicali organici. In plus metalotioneina chelateaza ionii de fier impiedicand desfasurarea reactiei Fenton. Un nivel ridicat al metalotioneinei poate fi asociat unei rezistente crescute la iradierea UVA.
Gena CL 100 codifica o protein fosfataza cu dubla specificitate (Tyr/Thr fosfataza) care inactiveaza specific o MAP kinaza implicata in raspunsul la stres. Activarea colagenazei ca raspuns la iradierea UVA nu reprezinta un mecanism de protectie, ci dimpotriva un raspuns distructiv ce contribuie la degenerarea colagenului interstitial si la fenomenul de imbatranire prematura a pielii.
Organismele sunt protejate de agresiunea speciilor reactive de oxigen prin mai multe modalitati: compartimentalizarea celulara, protectia conferita de compusii antioxidanti si sistemele enzimatice, capacitatea organismelor de a elabora raspusuri adaptative inductibile in conditii de stres oxidativ. In plus, procesele de reparare si turnover ajuta la minimalizarea leziunilor rezultate din atacul speciilor reactive de oxigen.
In conditii normale exista un echilibru intre sistemele antioxidante si sistemele generatoare de specii reactive de oxigen. Perturbarea acestui echilibru in favoarea sistemelor prooxidante determina instalarea stresului oxidativ, cu implicatii patologice (Sies, 1991; Morel si Barouki, 1999).
Sistemul enzimatic de protectie cuprinde: superoxid dismutaza, catalaza, glutation peroxidaza, glutation transferaza, glutation reductaza si glucozo-6-fosfat dehidrogenaza (Hasegawa si colab., 1992). Aceste enzime au o localizare preponderent intracelulara, mediul extracelular fiind mult mai expus atacului radicalic.
Compusii antioxidanti sunt clasificati dupa modul lor de actiune in: a) scavengeri - compusi ce previn stresul oxidativ prin captarea radicalilor liberi; b) antioxidanti "preventivi" - compusi ce actioneaza prin sechestrarea ionilor metalelor tranzitionale, prevenind reactia Fenton (Chapple, 1997). In Tabelul 2 sunt prezentati principalii compusi antioxidanti si mecanismul lor de actiune.
Tabelul 2. Principalii compusi antioxidanti, localizarea si mecanismul lor de actiune.
|
ANTIOXIDANT |
LOCALIZARE |
MOD DE ACTIUNE |
|
Proteine Feritina Transferina Lactoferina Ceruloplasmina Albumina |
- citoplasma - fluid extracelular - fluid extracelular - fluid extracelular - fluid extracelular |
- chelatare Fe2+ - chelatare Fe2+ - chelatare Fe2+ - chelatare Cu2+, oxidare Fe2+, scavenger O2- - chelatare Cu2+, scavenger OH., LOO., HOCl |
|
Compusi cu masa moleculara mica Vitamina E Ubichinona Carotenii Vitamina C Glutationul redus Acidul uric Bilirubuna |
- membrane - membrane - membrane - citoplasma - citoplasma, mitocondrii - sange - sange |
- scavenger OH., LOO., HOCl - scavenger OH., LOO., HOCl - scavenger OH., LOO., HOCl, 1O2 - scavenger OH., O2- - scavenger OH., O2- - previne peroxidarea lipidelor - previne peroxidarea lipidelor |
Mecanismele de protectie fata de radiatiile UVA de la nivelul pielii. Pielea prezinta o serie de structuri cu rol de protectie, cum sunt stratul cornos si melanina. Desi stratul superficial cornos indeplineste functia de bariera optica, determinand reflexia, dispersia si absorbtia radiatiei incidente, cea mai mare parte a radiatiilor UVA patrund profund in piele, pana la nivelul dermei. Se estimeaza ca radiatiile UVA pot fi absorbite de diferite componente ale sangelui, la nivelul vaselor de sange din derma (Tyrrell, 1996; Nechifor si colab., 1999). Radiatiile UVA actioneaza ca inductori ai enzimelor responsabile de sinteza poliaminelor. Poliaminele accelereaza derularea ciclului celular, stimuland proliferarea celulara si implicit ingrosarea epidermei si a stratului cornos. Ingrosarea stratului cornos (hiperkeratinizarea) ajuta la protectia pielii prin reflectarea razelor solare. Totusi, hiperkeratinizarea nu reprezinta un mecanism de protectie eficient pentru radiatiile UVA (eficienta este mai mare pentru radiatiile UVB).
Un mecanism suplimentar de protectie a epidermei consta in stimularea pigmentarii cu melanina. Trebuie subliniat faptul ca protectia prin pigmentarea cu melanina este dependenta nu numai de cantitatea totala de pigment ci si de compozitia sa chimica. Astfel, eumelanina este componenta de culoare neagra a melanozomilor care are un efect protector datorita capacitatii sale de a forma compusi stabili cu radicalii liberi, in timp ce phaeomelanina este componenta de culoare galbena ce nu are efect protector, ci dimpotriva actioneaza ca fotosensibilizator. Pe langa stimularea producerii de melanina de catre melanocite, radiatiile UVA determina efectul de pigmentare imediata ce consta in foto-oxidarea melaninei pre-existente, conducand la o amplificare pe termen scurt a pigmentarii, dar care nu confera protectie fata de expunerea ulterioara la radiatii UV.
In figura 2 sunt prezentate mecanismele de protectie constituiva si inductibila de la nivelul pielii. Nivelul antioxidantilor este constitutiv crescut in piele, cu mentiunea ca acest nivel este semnificativ mai mare in epiderma comparativ cu derma. In schimb, in epiderma nu au fost evidentiate mecanisme de protectie inductibile. In derma a fost evidentiat mecanismul inductibil mentionat anterior, de activare transcriptionala a hem oxigenazei I, corelat cu cresterea nivelului de feritina.
![]() Protectie constitutiva
Protectie constitutiva  Protectie inductibla
Protectie inductibla
Nivelele de antioxidanti sunt constitutiv
mai mari in epiderma comparativ cu derma
Glutation Vitamina
E Acid
ascorbic Acid
uric Ubiquinol Catalaza Superoxid
dimutaza Feritina
Hemoxigenaza-2
Epiderma Necunoscut
Derma Hemoxigenaza-1
![]()
![]() Feritina Protectie celulara
Feritina Protectie celulara
Nu apar leziuni membranare
Preventia
mortii celulare
mutagenezei
carcinogenezei
Figura 2. Mecanismele de protectie constitutiva si inductibila de la nivelul pielii fata de agresiunea radiatiilor UV.
In ultimii ani industria farmaceutica si cosmetica a lansat o gama larga de substante ce actioneaza ca filtre capabile sa absoarba fotonii UV, dintre care pot fi mentionate: Parsol 1789, Mexoryl SX, Octil Metoxicinnamat si Octocrilen. Produsele fotoprotectoare sunt caracterizate prin factorul de protectie. Acesta reprezinta o cifra ce indica de cate ori este prelungita durata proprie de protectie la soare, in conditii de siguranta, fiind exclusa aparitia eritemului actinic. O apreciere corecta a eficientei produselor fotoprotectoare trebuie facuta si in functie de capacitatea lor de a inhiba reactia de izomerizare a acidului urocanic si de a preveni acumularea proteinei p53 (Forestier si Mascotto, 1999; Cole, 2001).
Concluzii
Radiatiile solare exercita un efect benefic antirahitic prin stimularea sintezei vitaminei D, precum si un efect antidepresiv. Cu toate acestea, radiatiile UV constituie un factor de risc pentru aparitia cancerului cutanat si a unor tulburari oculare. Pentru a minimaliza acest risc se recomanda limitarea expunerii la radiatiile solare. Pentru persoanele care fac tratament cu agenti imunosupresori, recomandarea este si mai stricta, intrucat aceste persoane prezinta un risc de 500 ori mai ridicat decat persoanele obisnuite de aparitia a cancerului. De asemenea, copiii si adolescentii sunt mult mai susceptibili la carcinogeneza indusa de iradierea UV decat persoanele adulte. Radiatiile UVA sunt responsabile de fenomenul de imbatranire prematura a pielii.
Un alt aspect care nu trebuie neglijat se refera la faptul ca substantele cu actiune de filtre solare din compozitia lotiunilor cosmetice ofera o protectie eficienta fata de arsurile solare dar nu impiedica efectul imunosupresor al radiatiilor si pana in prezent nu se poate spune cu certitudine daca filtrele solare previn cancerul de piele.
Applegate L.A., Frenk E. (1995) Cellular defense mechanisms of the skin against oxidant stress and in particular UVA radiation. Eur.J.Dermatol. 5: 97-103.
Balasubramanian D. (2000) Ultraviolet radiation and cataract. J.Ocul.Pharmacol.Ther. 16: 285-297.
Chapple I.L.C. (1997) Reactive oxygen species and antioxidants in inflammatory diseases. J.Clin.Paradontol. 24: 287-296.
Cole C. (2001) Sunscreen protection in the ultraviolet A region: how to measure the effectiveness. Photodermatol.Photoimmunol.Photomed. 17: 2-10.
Cunningham M.L., Johnson J.S., Giovanazzi S.M., Peak M.J. (1985) Photosenzitized production of superoxide anion by monochromatic (290-405 nm) ultraviolet irradiation of NADH and NADPH coenzyms. Photochem.Photobiol. 42: 125-128.
Dean R.T., Fu S., Stocker R., Davies M.J. (1997) Biochemistry and pathology of radical-mediated protein oxidation. Biochem.J. 324: 1-18.
de Gruijl F.R. (2000) Photocarcinogenesis: UVA vs UVB. Methods.Enzymol. 319: 359-66.
Dovrat A., Weinreb O. (1999) Effects of UV-A radiation on lens epithelial NaK-ATPasa in organ culture. Invest.Ophthalmol.Vis.Sci. 40: 1616-1620.
Dudek E.J., Peak J.G., Roth R.M., Peak M.J. (1993) Isolation of V79 fibroblast cell lines containing elevated metallothionein levels that have increased resistance to the cytotoxic effects of ultraviolet-A radiation. Photochem.Photobiol. 58: 836-840.
Dumitru I.F., Nechifor M.T. (1994) Decrease in yeast glucose-6-phosphate dehydrogenase activity due to oxygen free radials. Int.J.Biochem. 26: 229-233.
Finley E.L., Busman M., Dillon J., Crouch R.K., Schey K.L. (1997) Identification of photooxidation sites in bovine α-crystallin. Photochem.Photobiol. 66: 635-641.
Forestier S., Mascotto R. (1999) Sun and UVA. SŐFW-Journal 8: 2-6.
Halliday G.M., Bestak R., Yuen K.S., Cavanagh L.L., Barnetson R.S. (1998) UVA-induced immunosuppression. Mutat.Res. 422: 139-45.
Hasegawa T., Kaneko F., Niwa Y. (1992) Changes in lipid peroxide levels and activity of reactive oxygen scavenging enzymes in skin, serum and liver following UVB irradiation in mice. Life Sci. 50: 1893-1903.
Huda A. (2000) UVA, UVB, and UVC and their possible effects. Health Phys.78 (5 Suppl): S75.
Kielbassa C., Roza L., Epe B. (1997) Wavelength dependence of oxidative DNA damage induced by UV and visible light. Carcinogenesis 18: 811-816.
Kohen E., Santus R., Hirschberg J.G. (1995) Biological effects of solar ultraviolet radiation. In: Photobiology, Academic Press, pp.275-302.
Kuluncsics Z., Perdiz D., Brulay E., Muel B., Sage E. (1999) Wavelength dependence of ultraviolet-induced DNA damage distribution: involvement of direct or indirect mechanisms and possible artefacts. J.Photochem.Photobiol. 49: 71-80.
Kuusilehto A. (2000) Transmission of UVA radiation through epithelium of oral mucosa and skin in rat and man. Photodermatol.Photoimmunol.Photomed.16: 189-191.
Leszczynski D., Fagerholm S., Leszczynski K. (1996) The effects of the broadband UVA radiation on myeloid leukemia cells: The possible role of protein kinase C in mediation of UVA-induced effects. Photochem.Photobiol. 64: 936-942.
Maines M.D., Traskshel G.M., Kutty R.K. (1986) Characterization of two constitutive forms of rat liver microsomal heme oxigenase. Only one molecular species of the enzyme is inducible. J.Biol.Chem. 261: 411-419.
Morel Y., Barouki R. (1999) Repression of gene expression by oxidative stress. Biochem.J. 342: 481-496.
Morliere P., Moysan
A.,
Nechifor M.T., Neagu M., Manda G., Gaghes M. (1999) Changes in albumin fraction after in vitro UVA irradiation of human plasma. J.Med.Biochem. 3: 283-287.
Peak M.J., Peak J.G., Carnes B.A. (1987) Induction of direct and indirect single-strand breaks in human cell DNA by far- and near-ultraviolet radiations: Action spectrum and mechanisms. Photochem.Photobiol. 45: 381-387.
Peak J.G., Pilas B., Dudek E.J., Peak M.J. (1991) DNA breaks caused bz monochromatic 365 nm utraviolet-A-radiation and their repair in human epithelioid and xeroderma pigmentosum cell. Photochem.Photobiol. 54: 197-203.
Robert M., Bissonauth V., Ross G., Rouabhia M. (1999) Harmful effects of UVA on the structure and barrier function of engineered human cutaneous tissues. Int.J.Radiat.Biol. 75: 317-26.
Shore R.E. (2001) Radiation-induced skin cancer in humans. Med.Pediatr.Oncol. 36: 549-554.
Sies H. (1991) Oxidative stress: Introduction.
In: Oxidative Stress: Oxidants and Antioxidants, Academic Press,
Somosy Z. (2000) Radiation response of cell organelles. Micron 31: 165-181.
Stary A., Robert C., Sarasin A. (1997) Deleterious effects of ultraviolet A radiation in human cells. Mutation Res. 383: 1-8.
Stoker R. (1990) Induction of heme oxygenase as a defence against oxidative stress. Free Rad.Res.Comms. 9: 101-112.
Stoker R., Lai A.,
Peterhans E., Ames B.N. (1989)
Antioxidant activities of bilirubin and biliverdin. In: Medical, Biochemical,
and Chemical Aspects of Free Radicals, Elsevier,
Tyrrell R. (1996) Activation of mammalian gene expression by the UV component of sunlight - from models to reality. BioEssays 18: 139-148.
Tyrrell R. (1999 Redox regulation and oxidant activation of heme oxygenase-1. Free Rad.Res.
van der Leun J.C. (1996) UV radiation from sunlight: summary, conclusions and recommendations. Photochem.Photobiol. 35: 237-244.
Wang S.Q., Setlow R., Berwick M., Polsky D., Marghoob A.A., Kopf A.W., Bart R.S. (2001) Ultraviolet A and melanoma: a review. J.Am.Acad.Dermatol. 44: 837-846.
Zigman S., McDaniel T., Schultz J.B., Reddan J., Meydani M. (1995) Damage to cultured lens epithelial cells of squirrels and rabbit by UV-A (99.9%) plus UV-B (0.1%) radiation and alpha tocopherol protection. Mol.Cell Biochem.143: 35-46.
Zigman S. (2000) Lens UVA photobiology. J.Ocul.Pharmacol.Ther.
Copyright © 2025 - Toate drepturile rezervate