|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
SINAPSELE NEURONALE
In functionalitatea tesutului nervos, transferul impulsului de la un neuron la altul se realizeaza prin intermediul unor jonctiuni intercelulare specializate, denumite sinapse interneuronale.
Astfel, impulsul nervos este transmis din neuron in neuron (din neuron presinaptic in neuron postsinaptic), de-a lungul unor circuite
|
|
|
Fig.
IV.2.4. Fibre nervoase amielinizate |
bine determinate, ceea ce permite o activitate integrativa a tesutului nervos in interiorul unui organism. Se apreciaza ca numarul sinapselor in sistemul nervos central uman este de ordinul a 1014.
Cunoasterea ultrastructurii sinapselor a facut posibila intelegerea si acceptarea "doctrinei neuronale", mult dezbatuta anterior. Structuri de tip sinaptic pot fi prezente si intre fibra nervoasa si o tinta nonneuronala, de exemplu muschi striat scheletal, muschi neted (vezi Capitolul III - Tesutul muscular) sau glande.
Functional, exista doua tipuri de sinapse: electrice si chimice. Acestea difera prin sistemul de transmitere a impulsului nervos si prin procesul de declansare a potentialului de actiune.
Sinapsele electrice
sunt mai putin frecvente la mamifere. Din punct de vedere ultrastructural
si molecular, fac parte din categoria jonctiunilor
distantate (gap), care permit (ca si in alte
tesuturi, respectiv muscular neted si muscular cardiac), schimburi
ionice intercelulare bidirectionale.
Ca regula generala, datorita pozitiei unui neuron intr-un
circuit, schimburile sunt unidirectionale, astfel incat impulsul electric
difuzeaza direct de la un neuron la altul. Transmiterea prin aceste
sinapse este foarte rapida. Sunt prezente la nivelul trunchiului cerebral,
cortexului cerebral si in retina.
Sinapsele chimice
reprezinta marea majoritate a sinapselor existente in organismul uman. La
nivelul lor transmiterea se realizeaza
prin intermediul unor substante chimice (neurotransmitatori si neuromodulatori).
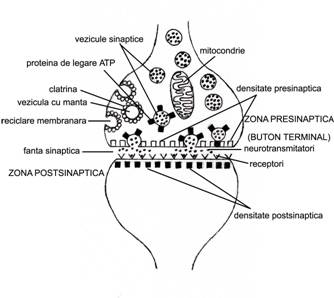
Aceste sinapse contin trei elemente constitutive: componenta presinaptica (eliberatoare de neurotransmitatori), fanta sinaptica (spatiu intercelular de 20-30 nm, care contine glicoproteine si glicozaminoglicani) si componenta postsinaptica (care contine receptorii pentru neurotransmitatori) (fig. IV.1). In acest tip de sinapse, influxul nervos este obligatoriu unidirectional.
In cursul functionarii unei sinapse chimice, sub influenta neurotransmitatorilor se poate realiza:
depolarizarea postsinapsei (prin deschiderea canalelor de sodiu) si generarea unui potential de actiune, numit potential postsinaptic excitator; in aceasta situatie, o astfel de sinapsa este considerata excitatorie;
nemodificarea polarizarii sau mai frecvent hiperpolarizarea (prin deschiderea canalelor de clor), cu instalarea unui potential postsinaptic inhibitor, situatie in care sinapsa este considerata inhibitorie.
Neurotransmitatorii nu sunt direct responsabili de reactia membranei postsinaptice, ci ei au rol de a induce modalitatea de raspuns a acesteia.
In sistemul nervos central sinapsele sunt localizate exclusiv in substanta cenusie, iar in sistemul nervos periferic - in ganglionii autonomi (simpatici si parasimpatici) si in organe periferice (receptoare sau efectoare).
|
Observatie |
In afara de sinapse, considerate jonctiuni intercelulare specializate, intre neuronii sistemului nervos central au mai fost evidentiate jonctiuni aderente si contacte in focar, similare celor din tesutul epitelial.
Rolul principal al acestora este de consolidare si mentinere in pozitie a neuronilor, fara a avea implicatii in transmiterea impulsului nervos.
|
|
|
Fig.
IV.1. Sinapsa - ultrastructura,
organizare moleculara |
STRUCTURA SI ULTRASTRUCTURA SINAPSEI
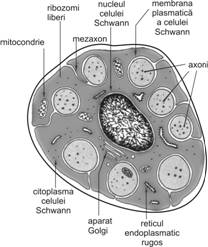
Desi in microscopia optica si coloratia de rutina nu este posibila identificarea sinapselor, coloratiile speciale (impregnare argentica - metoda Golgi) pentru evidentierea neuronilor fac posibila vizualizarea acestora, sub forma unor corpi ovali. Acesti corpi ovali reprezinta terminatiile axonale ale neuronului presinaptic in asociere intima cu suprafata neuronului postsinaptic.
La
nivelul sinapselor, cele doua membrane
(pre si postsinaptica) sunt paralele intre ele, chiar daca
traiectul este usor curbat. Intre cele doua membrane materialul
extracelular apare granular, omogen, de densitate electronica mica,
lipsit de structuri filamentoase sau fibrilare de ancorare.
Zona presinaptica
este formata frecvent din butonul
terminal, echivalentul unui corp
oval identificabil in coloratii speciale. Butonul terminal reprezinta
o portiune scurta si mai dilatata a fibrei nervoase (de
cele mai multe ori axonala). Exista insa si posibilitatea
ca zonele presinaptice sa apara sub forma unor dilatari situate pe lungimea axonului, numite butoni "en passant".
In vecinatatea butonului terminal, fibra nervoasa pierde teaca de
mielina si teaca celulelor Schwann. Citoplasma contine un
numar relativ mare de mitocondrii, elementele de citoschelet si
cativa tubuli scurti de reticul endoplasmatic neted.
Elementele ultrastructurale caracteristice sunt veziculele sinaptice (sinaptozomi) si particularitatile membranei presinaptice.
Veziculele sinaptice apar ca structuri delimitate de o membrana de tip unitate membranara, cu diametrul intre 30-100 nm. Enzimele din axoplasma impiedica degradarea neurotransmitatorilor.
La
o sinapsa in repaus veziculele numeroase sunt situate la oarecare
distanta de membrana presinaptica, iar cateva sunt atasate
acesteia.
La sinapsa in activitate, veziculele, extrem de numeroase, sunt atasate
membranei presinaptice.
Pe criterii ultrastructurale, se pot deosebi: vezicule mici (care contin neurotransmitatori) si vezicule mari cu miez dens (care contin neuromodulatori).
Veziculele
sinaptice mici, cu diametru de
40-50 nm, au un continut omogen electron-clar si o forma
sferica (vezicule S). In marea lor majoritate sunt localizate in
vecinatatea membranei presinaptice. Continutul lor este reprezentat,
in principal, de acetilcolina, purine si/sau aminoacizi. Exista
si un numar redus de vezicule mici cu un centru electron-dens, care
contin monoamine si/sau purine. Pe preparate electrono-microscopice
au fost identificate si vezicule mici electron-clare cu forma
(posibil artefactuala) aplatizata, cilindrica sau ovalara
(vezicule F), al caror continut este constituit in special de acidul γ-aminobutiric
(GABA).
Veziculele mari cu miez dens ating 70 nm diametru si prezinta central sau paracentral o zona electron-densa, separata de membrana printr-un halou clar. Aceste vezicule sunt situate la distanta de membrana presinaptica (zona activa), exocitoza lor avand loc pe laturile butonului terminal. Continutul lor este reprezentat in principal de neuromodulatori, uneori in asociatie cu neurotransmitatori "clasici". Unele dintre aceste vezicule contin proteine de asociatie; sunt asemanatoare cu granulele cromafine, acestea din urma atingand insa diametre de 250-280 nm.
Spre deosebire de datele clasice, s-a demonstrat in prezent ca intr-o vezicula sinaptica pot coexista mai multi mediatori.
Membrana presinaptica apare in microscopia electronica de transmisie ca avand o foita interna mai groasa si mai electron-densa, aspect datorat prezentei unui material dens, asociat cu numeroase mitocondrii. Prin tehnici speciale (crio-fracturare, microscopie electronica de baleiaj) s-a constatat ca de pe fata interna a membranei plasmatice se desprind proiectii in forma de conuri, orientate in interiorul citoplasmei. Acestea au un aranjament ordonat si sunt legate intre ele prin microfilamente de actina. Structura complexa a fost numita grila presinaptica sau densitate presinaptica si, impreuna cu veziculele sinaptice asociate, formeaza zona activa a sinapsei. Pe fata externa a membranei presinaptice, in ariile corespunzatoare dintre bazele expansiunilor coniforme, apar infundari ale membranei plasmatice numite sinaptopori.
Fanta sinaptica, delimitata intre neuronul presinaptic si neuronul postsinaptic, face parte din spatiul extracelular al sistemului nervos central.
Zona postsinaptica se gaseste situata fata in fata cu membrana presinaptica si apare la microscopul electronic ca o membrana asimetrica, cu foita interna ingrosata si electron-densa, prin depunerea unui material omogen. (Ingrosarea si densitatea postsinapsei depaseste pe cea a presinapsei.) Aceasta densificare este numita densitate postsinaptica sau lacis jonctional postsinaptic. Ea contine moleculele receptor pentru neurotransmitatori care, cuplate cu acestia, determina deschiderea canalelor ionice, permitand astfel modificarea permeabilitatii si, in consecinta, a polarizarii postsinapsei. Electrono-microscopic, la mica distanta de membrana postsinaptica se remarca o aglomerare de cisterne aglomerate paralele, constituind aparatul spinos, descris in special la baza spinilor dendritici.
Sinapsele in care membrana postsinaptica prezinta o densificare si ingrosare extrem de accentuata si o fanta sinaptica de 30 nm sunt denumite sinapse asimetrice; functional, sunt sinapse de tip excitator.
Mai putin frecvent, unele sinapse prezinta o membrana postsinaptica cu aceeasi grosime si densitate cu presinapsa si o fanta sinaptica ceva mai mica (20 nm); acestea sunt denumite sinapse simetrice si, functional, sunt, in general, de tip inhibitor.
In functie de componentele neuronale care intra in structura unei sinapse exista mai multe tipuri morfologice:
sinapsa axo-dendritica, in care axonul reprezinta presinapsa, iar dendrita, postsinapsa; numarul mare de dendrite determina o suprafata receptoare mai mare si, consecutiv, un numar mare de postsinapse;
sinapsa axo-somatica, in care postsinapsa este reprezentata de corpul celular al unui neuron;
sinapsa axo-axonala, in care postsinapsa este reprezentata tot de un axon; se realizeaza fie in segmentul initial al axonului, fie in vecinatatea extremitatii terminale a acestuia.
Mai pot exista sinapse dendro-dendritice, dendro-somatice si somato-somatice (rare).
Desi sinapsele sunt definite ca jonctiuni de comunicare intre neuroni, ele nu au numai rol de transmitere a impulsului nervos de la neuron presinaptic la neuron postsinaptic, ci pot chiar interveni asupra informatiei transmise. Aceasta interventie se realizeaza prin intermediul sinapselor axo-axonale, unde neuronul presinaptic, prin efectul (inhibitor sau excitator) pe care il produce asupra neuronului postsinaptic, participa indirect in sinapsele axo-dendritice sau axo-somatice pe care acesta din urma le stabileste, la randul lui.
In afara tesutului nervos, se pot realiza sinapse intre celule receptoare si neuroni (epiteliu senzorial) si intre celule efectoare si neuroni (fibre musculare, celule secretoare).
HISTOFIZIOLOGIA SINAPSEI
Impulsul nervos reprezinta un potential de actiune transmis de-a lungul axonului determinand depolarizarea membranei plasmatice care, la nivelul presinapsei, declanseaza deschiderea canalelor de calciu voltaj-dependente. Patrunderea calciului in terminatia presinaptica este urmata de procesul de eliberare a veziculelor sinaptice, fenomenele avand o anumita succesiune: deplasarea spre zonele active ale membranei presinaptice, atasarea la aceasta, fuziunea urmata de deschiderea sinaptoporilor, eliberarea neurotransmitatorului.
Etapa de ancorare a veziculelor sinaptice la membrana sinaptica este influentata de prezenta a doua proteine transmembranare: sinaptotagmina, cu afinitate pentru calmodulina si sinaptofizina, din familia conexinelor.
Etapa de fuzionare este influentata de existenta, pe membrana veziculelor sinaptice, a unui factor proteic NSF (eng. N-ethyl-maleimide-sensitive factor) - ATP-dependent - care se cupleaza cu un factor solubil de atasare a proteinelor (SNAP), pentru care exista receptori membranari. Consecutiv, are loc eliberarea de neurotransmitatori prin exocitoza, difuziunea lor in fanta sinaptica si, ulterior, cuplarea cu receptorii corespunzatori.
Exista, in raport cu localizarea, doua categorii de vezicule sinaptice: asociate zonei active si adiacente acestei zone. Acestea din urma constituie populatia veziculara de rezerva si sunt mentinute intr-un "conglomerat" datorita unei molecule proteice mici, sinapsina I, atasata de suprafata.
"Conglomeratul" intracelular este conectat la filamentele de actina ale citoscheletului, prin intermediul altor doua molecule, sinapsina II si proteina rab3a. El se desface in momentul in care sinapsina I este fosforilata, iar veziculele de rezerva se orienteaza inspre zona libera.
In cadrul acestui proces, membrana veziculelor sinaptice mici se adauga progresiv membranei presinaptice, determinand un excedent (surplus) de membrana. Se realizeaza astfel o permanenta deplasare a excedentului de membrana spre fetele laterale ale butonului presinaptic. La acest nivel se formeaza vezicule cu manta (continand clatrina), dupa modelul endocitozei, care se ataseaza apoi tubulilor REN. Aici va avea loc o sinteza locala de neurotransmitatori, pentru ca, printr-un proces de "inmugurire", REN sa produca noi vezicule sinaptice. Acest ciclu este cunoscut si sub termenul de reciclare membranara a veziculelor.
Pentru veziculele mari nu apare fenomenul de reciclare membranara, deoarece neuropeptidele sunt sintetizate la nivelul corpului celular, impachetate in vacuole golgiene si transportate prin flux axonal anterograd spre zona presinaptica.
Exista o mare varietate de neurotransmitatori si de neuromodulatori.
In categoria neurotransmitatorilor se inscriu cei considerati "clasici" (acetilcolina, catecolamine, serotonina sau 5-hidroxitriptamina, histamina), alaturi de purine (adenozina, ATP-ul), acizi aminati (glutamat, aspartat - cu rol excitator, glicina, GABA - cu rol inhibitor), monoxidul de azot.
Acetilcolina este eliberata la nivelul placii motorii, fibrelor preganglionare simpatice si parasimpatice, fibrelor postganglionare parasimpatice, fibrelor preganglionare din medulosuprarenala, fibrelor simpatice asociate glandelor sudoripare, precum si la nivelul unor sinapse in sistemul nervos central. Neuronii care sintetizeaza acest mediator poarta denumirea de neuroni colinergici. Postsinaptic, exista doua categorii de receptori: muscarinici (in fibrele musculare netede, cord, glande endocrine, ganglioni autonomi si sistem nervos central) si nicotinici (in ganglioni autonomi, sistem nervos central si fibrele musculare striate). Structura receptorului nicotinic este de tip pentamer - 2αβγδ, cele 5 subunitati delimitand central un canal ionic; prezinta un domeniu citoplasmatic, un domeniu membranar si un domeniu extracelular, extins pe o distanta de 6 nm in fanta sinaptica. Consecutiv actiunii unor medicamente sau toxine (de exemplu, toxina botulinica), cantitatea de acetilcolina poate deveni insuficienta, fie prin defect de eliberare, fie prin defect de cuplare cu receptorii; aceasta are consecinte grave la nivelul musculaturii respiratorii si scheletice.
Adrenalina,
noradrenalina si dopamina sunt denumite, in ansamblu, catecolamine datorita nucleului
catecolic comun existent in structura lor biochimica. Ele se formeaza
printr-o serie de reactii enzimatice, pornind de la aminoacidul
fenilalanina. Noradrenalina
este specifica sistemului nervos autonom, neuronii care sintetizeaza
acest mediator fiind numiti neuroni
adrenergici. Adrenalina este specifica sistemului nervos central. In
terminatia presinaptica, veziculele de catecolamine sunt asociate, in
proportie de 4:1, cu ATP-ul care are rol de cotransmitator.
Noradrenalina si adrenalina sunt sintetizate si la nivelul
medulosuprarenalei
(20-25% noradrenalina, 75-80% adrenalina); eliberarea de
adrenalina este stimulata de situatii critice: frica
acuta, stress intens, care determina reactiile de "lupta
sau fuga".
Monoxidul de azot se sintetizeaza in concentratii mici, printr-o reactie enzimatica in lant, direct in fanta sinaptica, unde difuzeaza si, actionand asupra membranei postsinaptice, declanseaza un potential de actiune.
Neuromodulatorii sunt reprezentati de neuropeptide cu actiune reglatoare pe receptori mai frecvent extrasinaptici; exista neuropeptide opioide si non-opioide. Categoria neuropeptidelor opioide cuprinde substante derivate din pro-opio-melanocortina, pro-enkefalina, pro-neoendorfina. Categoria substantelor non-opioide include produsii elaborati de celulele sistemului endocrin difuz, cu localizare digestiva (VIP, bradikinina, colecistokinina, substanta P, neurotensina), neurohormoni eliberati de hipotalamus (TRH, somatostatina), hormoni eliberati de neurohipofiza (oxitocina, vasopresina).
La
nivel postsinaptic, receptorii
pentru neurotransmitatori reprezinta canale de sodiu ligand-dependente; legarea
transmitatorului la acesti receptori determina deschiderea
acestor canale si influx de sodiu in postsinapsa, urmat de
depolarizarea acesteia. Consecutiv, are loc deschiderea canalelor de sodiu
voltaj-dependente din zona adiacenta si generarea potentialului
de actiune (raspuns excitator) care se va transmite
de-a lungul celui de al doilea neuron. Stimularea prin
neurotransmitatori este rapida (mai putin de
1 msec). Rezultatul actiunii
neurotransmitatorilor depinde de cantitatea de
neurotransmitator si durata eliberarii acestuia,
controlata si reglata printr-un proces de recaptare si/sau
degradare, la nivelul fantei sinaptice si a membranei postsinaptice. Recaptarea, denumita si preluare de inalta afinitate, se
realizeaza printr-un proces de endocitoza si
actioneaza pentru 80% din cantitatea totala de molecule
eliberate; astfel, este posibila reutilizarea, prin reciclare, a
neurotransmitatorilor. Restul de 20% este degradat prin
interventia unor enzime specifice (de exemplu, acetilcolin-esteraza pentru
acetilcolina, catecol-o-metil-transferaza si monoamin-oxidaza pentru noradrenalina),
localizate la nivelul membranei postsinaptice.
De asemenea, in transmiterea impulsului nervos intervin natura, numarul si distributia receptorilor postsinaptici, precum si conditiile existente in momentul activarii sinapsei, astfel incat un neurotransmitator poate produce efecte diferite, in situatii diferite.
|
Observatie |
Spre deosebire de neurotransmitatori, neuromodulatorii actioneaza asupra unor receptori specifici care, prin intermediul unui traducator de semnal (de tip proteine G sau kinaze), genereaza un sistem mesager secundar - AMP ciclic-protein-kinaze-fosforilaze - cu efecte reglatoare asupra postsinapsei.
Stimularea prin neuromodulatori este mai lenta (cateva minute).
Copyright © 2025 - Toate drepturile rezervate