 |
 |
|
|
 | Alimentatie | Asistenta sociala | Frumusete | Medicina | Medicina veterinara | Retete |
Dezvoltarea sexuala normala
I. Sexul genetic
Diferentierea sexuala la mamifere este rezultatul traducerii sexului genetic in sex gonadal. Gonadele elaboreaza pe urma steroizi sexuali si la masculi o proteina inhibitoare care duce la cresterea si diferentierea organelor sexuale interne si externe. Fara semne masculine evidente, gonadele vor suferi o diferentiere in ovare. Embrionul care se va modifica in femela are doi cromozomi X, in timp ce partile echivalente mascule au un singur cromozom X si unul Y. La mamifere, fiecare celula femela are de doua ori mai multe gene cu cromozomi X decat fiecare celula mascula. Cromozomii X contin pana la 5 din numarul total de gene gasite la mamifere. La mamiferele femele, al doilea cromozom X din fiecare celula este inactivat, asa incat vasta majoritate a genelor sale nu se manifesta niciodata. Cromozomul X inactiv se numeste corpul Barr. Inactivarea este intamplatoare in fiecare celula, astfel incat este afectat fie cromozomul matern, fie cel patern. Odata ce are loc inactivarea, toti descendentii acelei celule vor purta structura inactivata a celulei parentale. Concretizarea vizuala a acestui fenomen este pisica calico, care, in mod normal este intotdeauna femela. Un cromozom X poarta gena pentru culoarea neagra a blanii, in timp ce celalalt- pentru culoarea galben/orange a blanii. Insa, in embrionul cu 64 de celule, una din cele 2 perechi de cromozomi X si genele sale sunt inactivate la intamplare. Cum celulele se replica, celulele fiice mostenesc cromozomii X in starea lor activa sau inactiva, creind o pisica calico cu pete de blana neagra sau galbena (fig. 1).
Cercetari recente au aratat ca o gena specifica XIST (pronuntata "exist"= exista) poate regla inactivarea cromozomilor X. Cercetatorii au aratat ca aceasta gena este exprimata doar pe cromozomul X inactiv ( corpul Barr). Interesant este faptul ca gena s-a dovedit a fi doar transcrisa in ARNm si nu tradusa intr-o proteina. Omologi izolati ai genei XIST au fost gasiti in cromozomii X de la caini, pisici, soareci, oameni, vaci si iepuri. Mecanismul precis dupa care acest ARNm cauzeaza inactivarea unui cromozom X este subiectul unei investigatii intense. Cercetatorii speculeaza ca o doza dubla de gene ale cromozomului X activ ar putea fi letala sau cel putin ar necesita ca celulele la femele sa aiba o structura celulara biochimica complet diferita de celulele de la masculi. Un dezechilibru in numarul genelor cromozomului Y intre celulele masculine si feminine nu ar fi o problema deoarece cromozomul Y poarta foarte putine gene, din care majoritatea sunt necesare doar la masculi in spermatogeneza si in determinarea sexului.
Trebuie totusi subliniat ca sexul genetic este mult mai complex decat poate fi explicat doar de structura cromozomilor sexuali. Gene autozomale necesare pentru diferentierea completa a ovarelor si a testiculelor au fost identificate la mai multe specii. Cateva dintre aceste gene influenteaza migratia celulelor embrionare primordiale in timp ce altele regleaza prelucrarea sau functia antigenilor diferentiati necesari pentru dezvoltarea gonadala. Alte gene, pentru codificarea enzimelor, necesare sintezei de hormoni steroizi care vor media diferentirea tracturilor reproductive mascule sau femele (Fig.2-5).

FIGURA 1: INACTIVAREA CROMOZOMULUI X IN GAMETOGENEZA SI DEZVOLTAREA TIMPURIE
![]()
 Din motive necunoscute
Din motive necunoscute
un cromozom X este temporar inactivat Pisica calico, care este mereu femela
un mozaic de blana negru cu galben,
Din motive necunoscute, cromozonul cu un singur X este culori ce sunt codificate de o gena pe
 temporar inactivat cromozonul X. Femelele feminine ale
temporar inactivat cromozonul X. Femelele feminine ale
celule germinative majoritatii speciilor au mozaicul de
![]() cromozoni
x, desi fenotipul nu este de
cromozoni
x, desi fenotipul nu este de
![]()
![]()
![]() Meioza X ramane activ in tesuturile obicei la fel de vizibil precum culoarea
Meioza X ramane activ in tesuturile obicei la fel de vizibil precum culoarea
![]()
![]() somatice
masculine blanii.
somatice
masculine blanii.
![]()
Mascul
![]()


![]() X reactivat
X reactivat
![]()
![]() Zigot
Zigot
Blastocist La aproape cea de-a 64 etapa, chiar
Fertilizare timpuriu inaintea diferentierii celulare, un
cromozon x este inactivat la intamplare
![]()
![]()
![]()
![]() in fiecare celula
in fiecare celula
![]()
![]() Blastocist Activ Inactiv
Blastocist Activ Inactiv
tarziu A a
ococite Femela B b
![]()
![]()
![]()
![]()
![]()
![]()
Activ Inactiv (diviziunea celulara)
![]()
![]() meioza Reactivarea Inactivarea A a Statutul inactivarii x este
meioza Reactivarea Inactivarea A a Statutul inactivarii x este
X inactiv aleatorie rezulta B b mostenit de fiecare celula,
![]() in 2 tipuri de celule rezultand
intr-un mozaic de
in 2 tipuri de celule rezultand
intr-un mozaic de
cu cromoxonul x clone celulare exprimand genele
celule germinative derivat matern sau diferitilor cromozoni x.
patern
Tipic, femelele nu sufera de boli asociate cromozomului X, cum ar fi hemofilia. Celulele care poarta copia eronata a genei (genelor) active ar pute fi selectate impotriva. Alternativ, 50% ditre celulele care poarta cromozomul X cu gena eronata ar putea fi insuficiente sa cauzeze boli.
![]()

![]()
![]()

![]()
![]()

![]()
![]()
![]()
![]()
![]()
![]()



FIGURA 2: DOUA ETAPE IN DIFERENTIEREA GONADALA
tubul neutru
aorta
ansa intestinala
d.p.c. mezoderm intermediar
mezenter
arie ce va deveni
gonada bipotentiala
dezvoltarea gonadei WT 1 SF - 1 (gena FTZ - F1)
bipotentiale si alte gene
glomerus mezonefric tub mezonefric
(degenereaza) (degenereaza)
aorta canal Wolffian
mezenter canal Mülllerian
11 d.p.c. (WNT - 4)
ansa intestinala gonada bipotentiala
determinarea sexuala si SRY gene ce determina
diferentierea gonadala si alte gene ovarul
canal Mülllerian
In prima etapa, o parte a mezodermului intermediar se ingroasa pentru a da nastere gonadei bipotentiale. In a doua etapa, gonada bipotentiala se diferentiaza fie intr-un testicul, fie intr-un ovar. WT1 and SF-1 probabil participa in prima etapa si ar putea participa, de-asemena, si in a doua etapa. Sry nu functioneaza in cadrul generatiei gonadei bipotentiale,dar directioneaza ulterior diferentierea testiculara.Timpii aproximati in timpul dezvoltarii MURINE sunt aratati in stanga.Este posibil ca determinarea sexului sa se initieze mai devreme decat este indicat aici. Linia orizontala de-a lungul diagramei de sus indica nivelul sectiunii transversale reprezentata in diagramele ulterioare. Se observa ca SF-1 continua sa fie exprimat doar in testicule, unde se presupune ca activeaza enzime in celulele interstitiale necesare in sinteza de androgen si in plus activeaza genele MIS in sinciciu seminal. WNT-4 induce dezvoltarea canalului Mullerian la mamifere atat la embrionii femeli ca si la embrionii masculi.

![]()


![]() degenerativ canal Mülllerian
degenerativ canal Mülllerian
![]()
![]() 13 d.p.c. canal
Wolffian canal Wolffian
13 d.p.c. canal
Wolffian canal Wolffian
( (devine canal deferent) degenerativ
![]()
![]() testicol
testicol
![]()
![]()
![]()
![]() SF - 1 ovar
SF - 1 ovar
substanta Müllleriana testosteron
inhibatoare (MIS) estrogen
![]()
![]()
![]()
![]()

![]()
![]()
II. SRY-TDF, antigenul H-Y si diferentierea gonadala( vezi Fig. 2-5)
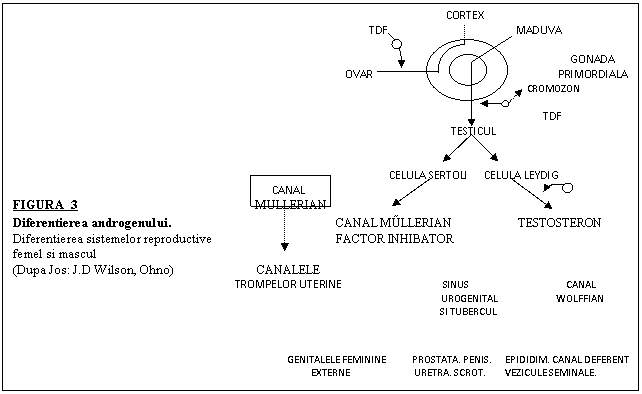
Diferentierea sexuala la mamiferele infraclasei Eutheria poate fi considerata ca fiind alcatuia din 3 procese fundamentale: (1) Diferentierea gonadala care este predeterminata genetic; (2) diferentierea oragnelor sexuale interne ( uter, tesuturi sexuale accesorii mascule) si (3) diferentierea organelor genitale externe mascule si femele. Ultimele doua procese sunt mediate de steroizii sexuali secretati de gonada diferentiata femela sau mascula ( vezi mai jos). In completare, testiculul secreta si substanta inhibitoare a conductului Mulerian (MIS), ceea ce cauzeaza regresia acestei structuri in dezvoltarea embrionului mascul ( vezi mai jos). O stiinta a acestor procese este esentiala pentru intelegerea anormalitatilor care ar putea surveni in ceea ce priveste dezvoltarea unui individ ambiguu sexual.
III .Diferentierea gonadala(vezi Fig. 2-5)
Dezvoltarea gonadei nediferentiate intr-un ovar sau testicul este mediata genetic si este evenimentul critic initial din care va rezulta ulterior din punct de vedere fenotipic un mascul sau o femela. In trecut se credea ca gonada bipotentiala se va diferentia pasiv intr-un ovar numai cand genele determinante testiculare nu sunt prezente si active in timpul embriogenezei. Dovezi recente au aratat, totusi, ca nu ar fi cazul in totalitate. O gena numita DAX-1, purtatoare de cromozom X, controleaza, probabil, diferentierea gonadei primordiale intr-un ovar. S-a banuit pentru mult timp ca o singura gena pentru inducerea diferentierii testiculare era purtatoare de cromozom Y. Dovezi mai vechi au sugerat ca diferentierea testiculara a fost mediata, la indivizii purtatori de cromozomi Y, de o glicoproteina, care a fost numita H-Y( antigen histocompatibil). S-a dovedit acum a nu fi cazul. Recent, un singur fragment de 35 Kb, localizat pe bratul scurt al cromozomului Y, denumit SRY( pentru determinarea sexului regiunii Y) a fost identificat la oameni si la majoritatea mamiferelor, ceea ce induce producerea unei proteine numita factor determinant testicular( TDF) (Fig. 2 si 3). Se stie acum ca TDF este responsabil cu diferentierea gonadei bipotetiale intr-un testicul. TDY a fost caracterizat ca o proteina indicatoare de zinc care probabil se fixeaza in aumite regiuni superioare ale genelor care participa la diferentierea testiculelor. Deci, se pare ca gena SRY este cheia reglatoare in dezvoltarea testiculelor. Totusi, se banuieste ca alte gene, purtatoare de autozomi, cum ar fi WT1( vezi Fig.2 luata dintr-o revizuie a Celulei 76:603-607, 1994) participa de-asemenea in evenimentele initiale si, probabil si-n cele ulterioare diferentierii gonadale. Recent, s-a demonstrat ca o gena FTZ-F1 ar codifica o proteina receptoare denumita factor steroidogenic 1( SF1), a carei expresie este gasita in crestele urogenitale mascule si femele in dezvoltarea timpurie a mamiferelor. Fara expresia lui SF-1, gonadele femele sau mascule si glandele suprarenale nu s-ar dezvolta. In acest fel genele functionale WT-1 si FTZ-F1, par a fi esentiale in dezvoltarea gonadala la ambele sexe. Este, de-asemenea, probabil ca proteina SF-1 sa medieze alte functii odata ce se diferentiaza testiculele( vezi mai jos). Este, de-asemenea, posibil ca MIS( vezi discutia urmatoare ) sa influenteze dezvoltarea gonadala , din moment ce expunerea ovarelor in mediul acestei glicoproteine le poate cauza pierderea ovocitelor si dezvolta structuri cordonale asemanatoare testiculelor.Acest lucru ar putea fi important, luand in considerare dezvoltarea gonadala anormala gasita la o vita cu sindromul Freemartin ( vezi notitele despre diferentierea sexuala anormala). Probabil, cercetarile viitoare vor arata ca alte gene, purtatoare de autozomi, sunt implicate in diferentierea gonadala.
IV. Factorii endocrini asociati cu dezvoltarea tracturilor reproductive si organele sexuale accesorii( vezi Fig.2-5)
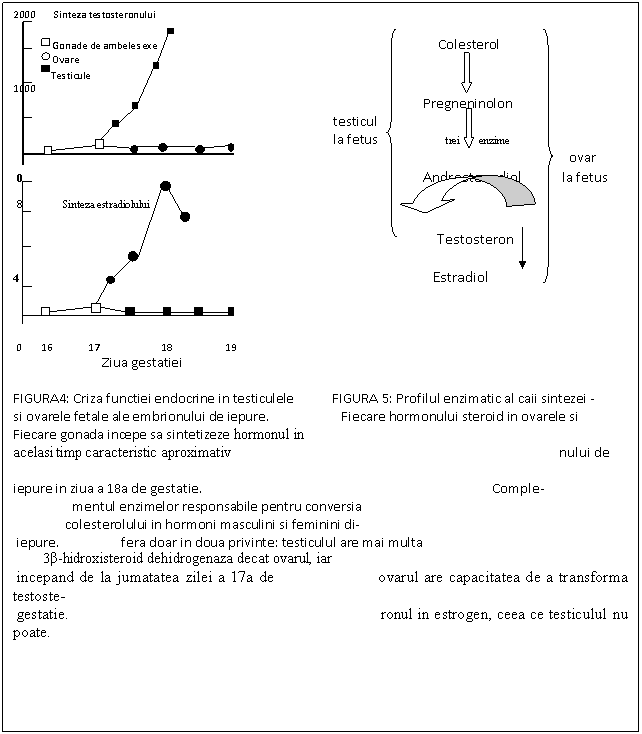
O data formate, testiculele si ovarele vor coordona toate celelalte aspecte ale dezvoltarii sexuale care duc la caracteristicile fenotipice fie mascule, fie femele. In cea mai mare parte, substantele care cauzeaza dezvoltarea sexuala fenotipica sunt steroizii sexuali (steroixii C-19= androgeni si steroizii C-18= estrogeni). Testiculele si ovarele dobandesc capacitatea de a secreta hormonii caracteristici in acelasi timp in dezvoltarea embriologica (Fig.4). La mascul, coincide cu dezvoltarea celulelor lui Leydig si precede atacul virilitatii tractului genital. Dovezi recente au aratat ca SF-1 activeaza enzime steroidogene in celulele lui Leydig,cum ar fi 3β-hidroxisteroid dehidrogenaza, care sunt necesare pentru sinteza de testosteron( vezi Fig.1). Totusi , la femele, ovarele secreta estrogeni inainte de a avea loc, din punct de vedere histologic, diferentierea gonadala. Testiculul fetal contine cam de 50 de ori cantitatea de 3β- hidroxisteroid dehidrogenaza prezenta in ovar( Fig. 5). Aceasta enzima este limitata cantitativ in sinteza testosteronului si in asa fel semnificativ mai mult testosteron este sintetizat in testicule decat in cazul ovarelor. In contrast, ovarul fetal are capacitatea de a transforma cantitatea mica de testosteron sintetizat in estradiol 17β. Testiculul, in aceasta etapa, este lipsit de enzimele aromatizate necesare in formarea estrogenilor. Toate celelalte enzime in drumul sintezei hormonilor steroizi din colesterol sunt egale acum si in ovare si in testicule. Totusi, diferente in cazul vitezei de sinteza a steroizilor sau prezenta doar a catorva enzime in gonade in aceasta perioada critica poate avea consecinte profunde pentru diferentierea sexuala a unui individ.
Intrebarea,daca vitezele formarii de hormoni steroizi sunt si ele regulate in aceasta etapa timpurie de dezvoltare, momentan nu este rezolvata.Majoritatea dovezilor sugereaza ca inceputul sintezei hormonilor steroizi in ovarul embrionic este autonom in timp ce proteina SF-1 mediaza sinteza androgenilor (vezi mai sus). Tarziu in embriogeneza, productia hormonilor la toate mamiferele este gonadotropin-dependenta( gonadotropina corionica(hCG) la primate , ser de iapa gestanta la cai, si probabil hormoni luteinizanti(LH) la alte specii).
Pentru dezvoltarea tracturilor reproductive si a organelor sexuale accesorii pentru oricare sex, sunt prezente tesuturi discrete indiferent de etapa bisexuala. In absenta testiculelor, componentele femelei (conductul Mullerian) se vor dezvolta si conductul Wolffian va regresa. In aceasta etapa de dezvoltare , testiculele contin in mod normal doua tipuri importante de celule- celule interstitiale( Leydig) si sincitiu seminal( Sertoli), care vor fi decisive pentru virilizarea tracturilor reproductive. Celulele Sertoli embrionare secreta o proteina cunoscuta ca substanta inhibitoare mulleriana (MIS) care induce regresia activa a conductului mullerian. Interesant este ca, probe noi sugereaza ca probabil ambele SF-1 si SRY sunt necesare pentru activarea genelor MIS in celulele Sertoli. Asadar SF-1 nu pare a fi esentiala doar pentru dezvoltarea gondala dar este de-asemenea necesara pentru producerea de androgen si a MIS in testiculele embrionare( vezi Fig. 2 si 3). In urma discutiei anterioare, celulele Leydig embrionare secreta testosteron ceea ce cauzeaza dezvoltarea conductului Wolffian in epididim, canal deferent, canale ejaculatoare si vezicule seminale. Diferentierea organelor sexuale accesorii( penis si scrot) nu este, totusi, controlata direct de catre testosterone. Precursorii lor embrionici( sinusul urogenital, tubercul genital, pliurile uretrale si umflaurile labioscrotale) contin o enzima, 5α- reductaza, care transforma testosteronul in steroidul 5α-dihidrotestosteron( 5α- DHT)( Fig.3). Apoi, acest hormon este legat de receptori nucleari specifici din celulele primordiale. Legatura nucleara, complexul receptor-hormon cauzeaza traducerea ARNs specific, care induce diferentierea acestor tesuturi in prostata, glande bulbo-uretrale( sinus urogenital), penis( tubercul genital), si umflaturile labioscrotale( scrot).In absenta MIS, din sistemul Mullerian se va dezvolta oviductul, uterul si o mare parte a vaginului. Aseanator, in absenta unei secretii suficiente de testosteron, sistemul conductului Wolffian va regresa si organele genitale externe ale femelei se dezvolta din schita comuna, care are potentialul sa se dezvolte in organe genitale mascule.
Recent ( Natura 397:405, 1999), cerecetatorii au identificat o gena, WNT-4, care in mod precis induce dezvoltarea conductului Mullerian atat la mamiferele mascule cat si la cele femele. Mutatii ale WNT-4 nu au nici un efect asupra fenotipului unui mascul, dar derivatii conductului Mullerian sunt absenti la femele. In cazul femelelor afectate, conductul Wolffian continua totusi sa se dezvolte. De asemenea, se pare ca WNT-4 suprima dezvoltarea celulelor Leydig in gonadele femelei si ar putea fi, de asemenea, necesara pentru mentinerea celulelor embrionare femele.
Embrionii femeli au acelasi sistem receptor androgen si aceeasi abilitate de a raspunde la androgeni ca si embrionii masculi. Diferentele intre dezvoltarea fenotipica la femela si mascul a tracturilor reproductive si a organelor genitale externe constau doar in natura produsilor hormonali. De exemplu, daca un embrion femel este expus androgenilor, se va viriliza ca masculul. ( Vezi discutia despre Hermafroditism in notitele ce urmeaza.)
Copyright © 2025 - Toate drepturile rezervate