|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
INTERACTIUNI NECOVALENTE (REVERSIBILE)
Acizii nucleici interactioneaza reversibil cu diversi reactanti. molecule de apa, ioni metalici sau complexi metalici, molecule organice mici sau proteine.
In categoria moleculelor mici, alaturi de apa intra si compusii organici cu M < 100 Da; ele includ medicamente, cancerigeni, antiobiotice s.a.
Interactiunile necovalente pot conduce la efecte contrarii datorita caracterului reversibilal acestora; astfel, in functie de concentratia si intensitatea interactiunilor, apa, ionii metalici sau cationii organici pot fie sa conduca la stabilizarea structurilor, fie, dimpotriva, sa determine tranzitii B-Z, formarea de structuri cruciforme sau alte modificari structurale. Impachetarea acizilor nucleici in capsida virala, in cromatina nucleara etc. este foarte sensibila la actiunea reversibila a moleculelor mici.
Importante sunt si interactiunile cu molecule cu actiune medicamentoasa, numeroase droguri cu efect anticancerigen, antiviral, antibacterian isi exercita actiunea prin intermediul unor interactiuni reversibile, intens studiate au fost atat antibioticele naturale de tipul adriamicinei, cat si cele sintetice .
Desi interactiunile se realizeaza atat cu molecule de ADN, cat si cu molecule de ARN, totusi majoritatea acestora au vizat ADN din mai multe motive:
reprezinta "tinta" atat in procesele de cancerizare, cat si de reversie a neoplaziilor;
deoxinucleozidele sunt mai usor de sintetizat, devenind astfel posibila studierea unor aspecte de tip conformational, tipuri de legaturi etc. si identificarea, in ultima instanta, a unor noi tipuri de medicamente.
1. Tipuri de interactiuni reversibile
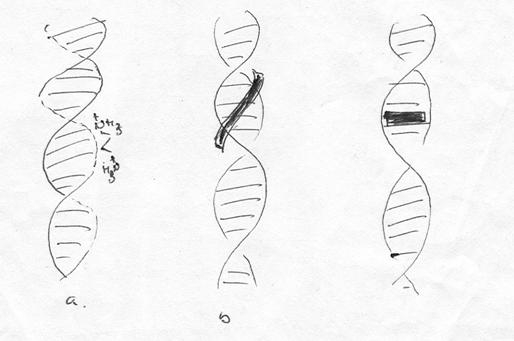
Pana in prezent au fost identificate trei variante de interactiuni primare reversibile intre molecule mici si duplexul ADN:
|
|
||
|
Interactiuni electrostice |
Legare in sant |
Intercalari |
interactiuni de tip electrostatic - se realizeaza cu resturile fosforice aflate spre exteriorul elicei;
interactiuni de tip "legare in sant" - molecula care se leaga se plaseaza fie in santul major, fie in cel minor, interactionand cu partile laterale ale bazelor azotate;
intercalari - molecula se intercaleaza intre planurile bazelor azotate, realizandu-se interactiuni intre nucleele aromatice.
Exemple de particule (cationi) care se leaga reversibil de ADN:
Interactiuni electrostice: Na , Mg2+, H3N+CH2CH2CH2N+H3
|
|
|
Agenti de legare in sant si de intercalare |
Interactiuni electrostatice "externe" elicei
Primele doua tipuri de interactuni nu conduc in mod necesar la modificari conformatonale, in schimb, intercalarile conduc la modificari atat ale unghiurilor, cat si a distantelor dintre baze, precum si din coloana fosfo-zaharica.
Interactiuni de tip "condensare"
Acizii nucleici sunt
polielectroliti cu o inalta incarcare electrica,
determinata de gruparile fosforice; pentru B-ADN, distanta - in
proiectie - dintre doua sarcini este de 1,7 Å (daca toate
gruparile ar fi ionizate) - distanta reala, in spatiu, este
insa ceva mai mare. S-a calculat ca daca distanta dintre
sarcini ar fi mai mica de 7 Å, molecula devine instabila. Cum
insa moleculele sunt stabilizate, s-a presupus ca in solutie se
realizeaza interactiuni intre gruparile fosforice si ionii
de semn contrar din solutie (cationi), proces denumit "condensare de contraioni". Aceasta produce o variatie
nefavorabila a valorii entropiei, variatie care este
insa compensata de variatia entalpiei libere. Pentru o
anumita conformatie, densitatea de sarcina negativa,
si deci gradul de "condensare de contraioni" este
constant astfel ca s-a presupus ca, chiar daca concentratia
de sare a solutiei variaza, "condensarea" ramane
Termenul de "condensare" a fost imaginat presupunand ca ionii metalici (in general cationii) isi pastreaza apa de hidratare, desi interactioneaza cu ADN. Complexul ion-apa este relativ liber, nefiind legat de un anumit centru, putand astfel aluneca de-a lungul coloanei fosfo-glucidice; este afectat insa gradul de hidratare al acidului nucleic. De asemenea, se modifica valoarea Tm - "temperatura de topire" a ADN. Defecte similare pot fi determinate si de celelalte tipuri de interactiuni.
Ionii cu sarcini mai mari (Mg2+, diamine ionizate) interactioneaza mai puternic, ei putand dezlocui ionii monovalenti. Alti cationi - cei ai metalelor tranzitionale (Zn2 , Cu2+, Hg2+) - se pot lega atat prin interactiuni similare celor de mai sus (nelocalizate), dar pot realiza si interactiuni localizate la o anumita grupare fosforica, avand astfel efecte contrare asupra Tm.
In concluzie, orice cation se poate lega de ADN printr-o interactiune de tip condensare; rezulta un complex nespecific, instabil, situat spre exteriorul coloanei fosfo-glucidice. Constantele de asociere / disociere sunt ridicate, astfel ca procesele sunt rapide.
Se considera ca si celelalte moduri de interactiune "in sant" sau intercalarea se realizeaza, initial, printr-o interactiune de tip Debye-Huckel, urmata de condensarea si alunecarea de-a lungul coloanei pana cand se atinge "tinta", la care legarea este specifica. De asemenea, disocierea acestor complecsi implica tranzitia prin forme "condensate".
Legari "in sant"
Diverse molecule, atat de dimensiuni mici, cat si molecule mai mari - oligonucleotide, proteine etc. se pot fixa si interactiona cu acizii nucleici in unul dintre cele doua santuri. Cele doua santuri difera, de regula, semnificativ in privinta potentialului electrostatic, capacitatea de formare de legaturi de hidrogen, grad de hidratare sau efecte sterice, astfel ca apare o "selectie" a modurilor de legare: de obicei moleculele mici prefera santul minor, in timp ce moleculele mari se leaga, preferential, in santul major.
De obicei, moleculele mici capabile sa
se lege in santul minor contin nuclee aromatice de tipul
pirolului, furanului sau benzenului. Aceste molecule, printr-o rotire
adecvata, se pot fixa in santul minor, deplasand moleculele de apa. De obicei, legarea se realizeaza in regiunile
A:T. Molecula intra in "vale" si interactioneaza cu
"peretii" acesteia, reprezentati de "marginile" resturilor de baze
azotate. Interactiunile sunt diverse: van der Waals, legaturi de
hidrogen, care se pot forma cu O2(T), N-3
(A). In cazul dubletului G:C, se pare ca
gruparile aminice blocheaza steric legarea
2.2.1. Legarea netropsinei si distamicinei
Una dintre cele mai studiate molecule, din punct de vedere al capacitatii de legare in "sant" este netropsina; Dickenson si colaboratorii au reusit sa obtina cristale formate din netropsina si un duplex d(CGCGAATTCGCG). Netropsina se leaga in zona centrala AATT, deplasand "spinii" formati de moleculele de apa. Cele trei grupari -NH ale moleculei se situeaza spre interiorul complexului, formand legaturi de hidrogen cu N-3(A) si O2(T). Ciclurile pirolice sunt aproximativ paralele cu peretii santului. Datorita rotirii elicei, doua inele pirolice formeaza un unghi de aprox. 33
Capetele cationice ale netropsinei sunt asociate cu prima molecula A din secventa AATT. Interactiunile sterice dintre resturile CH din ciclurile pirolice si ADN impiedica molecula de netropsina sa "coboare" prea mult in sant, ca urmare, legaturile de hidrogen formate sunt mai lungi (3,3 - 3,8 Å) fata de valorile normale (< 3 Å). Ca urmare a legarii netropsinei, santul se largeste putin, iar axa elicei sufera o indoire in regiunea de legare. Nu se produc alte modificari semnificative ale duplexului ADN.
Acelasi autor, Dickenson, impreuna cu Cown au reusit sinteza unei alte molecule toxitropsina (exista de fapt mai multe variante care recunosc atat grupurile A:T, cat si G:C.) In toxitropsina, unul sau mai multe inele pirolice sunt inlocuite prin cicluri imidazolice. Atomul de azot imidazolic poate forma legaturi de hidrogen cu grupul G:C, existenta acestora fiind confirmata prin determinari RMN.
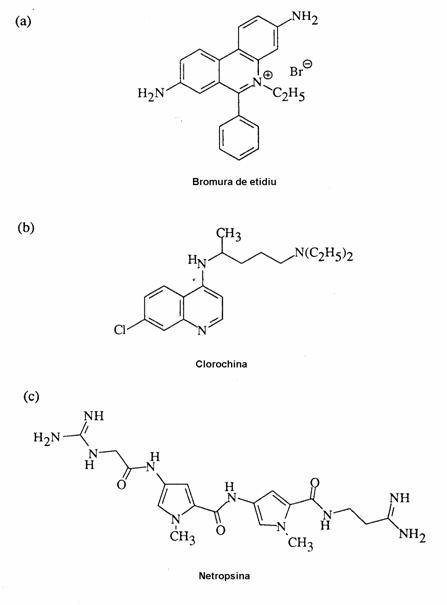
Alte molecule studiate au fost distamicina, cu o structura asemanatoare netropsinei, compusul Hoechst 33258 - un antibiotic, ce contine inele benzimidazolice.
Acestea dau legaturi "in sant" care sunt similare celor realizate de netropsina. Asemanator se comporta si SN 6999 sau medicamentul antimalaric clorochina.
Se pare insa ca ultimul compus poate da si intercalari, deoarece nu are o structura optima pentru a realiza legaturi "in sant".
Intercalari
Primul care a descris complecsii de tip intercalativ a fost Leiman, care la inceputul anilor '60 a realizat o serie de studii fizice - interactiunea ADN cu cationii aromatici plani. Studiile realizate de Leiman au condus la elaborarea modelului "clasic" de intercalare, conform caruia moleculele care se intercaleaza indeparteaza bazele cu 3,4 Å, spira alungindu-se in mod corespunzator.
Ulterior studiile s-au extins mult, numeroase molecule dand compusi de intercalare cu ADN: bromura de etidiu, bromura de propiliu, daunomicina, adriamicina.
Formarea complecsilor de intercalare poate fi decelata prin determinari de coeficienti de sedimentare sau de vascozitate, utilizandu-se fragmente scurte de ADN. O alta metoda utilizata a fost dicroismul circular, care permite identificarea compusilor de intercalare fata de cei legati "in sant". Intercalarea in stiva de baze a unei molecule nu conduce la o modificare apreciabila a valorii determinate, in schimb in cazul legarii "in sant" valoarea de dicroism isi schimba semnul.
Prin utilizarea unei metode denumite electric dicroism, Crotheus si colab. au presupus ca bazele adiacente moleculei intercalate isi modifica unghiul de rotire fata de planul perpendicular pe ax, pentru a se asigura o suprapunere cat mai adecvata a acestor molecule, sistemul inclinandu-se apoi cu 20-25 , pentru a se optimiza interactiunile cu restul de baze. Se pare ca molecula de ADN nu se incovoaie in punctul de intercalare.
Determinarile au demonstrat ca distanta dintre baze creste cu mai putin de 3,4 Å; se modifica si unghiul de rotire, pentru fiecare pereche de baze.
1. Complexul de intercalare format de Daunomicina
Daunomicina - un antibiotic - formeaza un complex de intercalare cu oligomerul d(CGTACG). La legare, daunomicina are axa mare a moleculei aproape perpendiculara pe cele doua baze azotate adiacente. Restul amina-glucidic se gaseste in santul minor, in timp ce gruparea metoxi de la celalalt capat al moleculei patrunde in santul major, iar nucleele centrale B si C se situeaza intre bazele azotate. Hexamerul leaga doua molecule de Daunomicina, care se intercaleaza intre nucleotidele CG terminale. Zona centrala isi pastreaza o structura de tip B-ADN, dar cu o slaba distorsiune a coloanei fosfo-glucidica.
In schimb, restul moleculei pentru a lega antibioticul, se modifica accentuat, astfel ca perechea G:C este deplasata lateral fata de santul major, modificand axa elicei, dar ea nu este incovoiata la punctele de legare, mentinandu-se unghiul normal de rotire al bazelor, de aprox. , bazele fiind insa inclinate cu 8 . Existenta acestor modificari conformationale demonstreaza flexibilitatea moleculei de ADN si capacitatea acesteia de a accepta un numar mare de molecule de intercalare.
Ca urmare a intercalarii, distanta dintre bazele G:C creste cu aprox. 3,4 Å. Gruparea hidroxil din ciclul A da legaturi de hidrogen cu N-3(G) (legatura donoare) si, in acelasi timp, da o legatura acceptoare cu gruparea - N2H2 a aceleeasi molecule G.
Trebuie insa precizat si faptul ca rezultatele anterioare au fost obtinute pe un fragment ADN foarte scurt. Se presupune ca in molecula nativa legarea se realizeaza la perechi A:T.
Copyright © 2025 - Toate drepturile rezervate