|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
La noi in tara, prin cercetarile lui V. Parvulescu si A. Oprescu, s-a putut stabili raspandirea geografica a genurilor de Ixodidae. Caracteristicile acestei distributii sunt zonarea dupa latitudine si dependenta de factorii climatici, temperatura si umiditate.
In Romania, Familia Ixodidae este reprezentata prin trei subfamilii: Ixodinae, Haemaphysalinae si Amblyomminae, ce cuprind 6 genuri si 24 specii (Tabelul 12, Figura 2) (Feider 1965).
Zonarea este modificata de influenta Carpatilor, care face ca hotarul nordic al genurilor Hyalomma si Rhipicephalus sa coboare mai spre sud, ele preferand locuri mult mai uscate; astfel, frontiera nordica a acestor genuri este cuprinsa intre izotermele anuale de 10 si 11°C.
Genurile Ixodes si Dermacentor au o raspandire generala in toata tara, primul gen avand insa o preferinta marcata pentru umiditate si regiuni de padure si lunca.
Studiul raspandirii geografice pe specii in Romania a fost facut de catre Z. Feider, C. Rauchbach si I. Mironescu (1958) (Plansa 61
Controlul populatiilor de ixodide vectoare
Modificarile de mediu si controlul fizic
In anumite situatii modificarea mediului propice ixodidelor a condus la reducerea semnificativa a populatiilor si chiar la disparitia lor.
Pot fi utile pentru distrugerea ixodidelor active: incendiile controlate, reducerea numarului de animale ce pot servi drept gazde si crearea unor habitate improprii pentru eclozare, naparlire, depunerea oualor si activitatea de cautare a gazdelor.
Ixodidele necesita un grad crescut de umiditate pentru desfasurarea activitatii; prin arderea vegetatiei microclimatul de la nivelul solului devine prea uscat pentru acestea.
Tabel 11. Boli transmise de capuse.
|
Boala infectioasa |
Specia vectoare |
Distributia geografica |
|
Bacterioze |
||
|
Lyme disease |
Ixodes ricinus, Ixodes pacificus, Ixodes scapularis, Ixodes dentatus, Ixodes uriae, Ixodes persulcatus, Ixodes trianguliceps, Ixodes hexagonus, Ixodes nipponensis | |
|
Febra recurenta |
Ornithodoros moubata, Ornithodoros sonrai, Ornithodoros graingeri, Ornithodoros erraticus Ornithodoros verrucosus Ornithodoros tartakovski, Ornithodoros tholozani Ornithodoros hermsi Ornithodoros talaje, Ornithodoros parkeri Ornithodoros turicata Ornithodoros rudis |
Africa, Asia, Statele Unite ale Americii, Europa, Orientul Apropiat, zona Mediteranei, Canada, |
|
Tularemia |
America, Europa, Japonia, Israel, Africa |
|
|
Febra Q |
Ixodes dentatus, Ixodes trianguliceps, Rhipicephalus sanguineus, Amblyomma americanum, Dermacentor andersoni, Dermacentor reticulatus | |
|
Ehrlichioza |
I. scapularis, I. pacificus, I. ricinus, Amblyomma americanum | |
|
Febra Muntilor Stancosi (tifos exantematic) |
Statele Unite ale Americii, Canada, Mexic, America de Sud |
|
|
Tifosul de capusa Siberian |
Ixodes spp | |
|
Febra butonoasa |
Rhipicephalus sanguineus Haemaphysalis spp, Ixodes spp, Hyalomma spp, Amblyomma spp, Boophilus spp, Dermacentor spp |
Africa, Europa, Asia, zona Mediteranei, |
|
Tifosul de capusa de Queensland |
Ixodes holocyclus | |
|
Arboviroze |
||
|
Febra hemoragica Omsk |
Ixodes persulcatus, Ixodes apronophorus, Dermacentor pictus, Dermacentor marginatus |
Siberia |
|
Encefalita rusa de primavara -vara |
Ixodes persulcatus |
Europa Centrala, fosta URSS |
|
Louping ill |
Ixodes ricinus |
Insulele Britanice, Spania |
|
Powassan |
Dermacentor andersoni, Ixodes spp |
Statele Unite, Canada |
|
Febra de padure Kyasanur |
Haemaphysalis spp, Ixodes spp, Rhipicephalus turanicus |
India |
|
Febra hemoragica Crimeea-Congo |
Hyalomma marginatum |
Asia, Europa, fosta URSS, zona Mediteranei, China, Orientul Mijlociu, Africa |
|
Febra de capusa de Colorado |
Dermacentor andersoni, Ixodes spp |
Statele Unite ale Americii, Canada |
|
Virusuri negrupate sau alte virusuri |
||
|
Coriomeningita limfocitara |
Amblyomma variegatum, Rhipicephalus sanguineus, Dermacentor andersoni |
Etiopia, Canada |
|
Piroplasmoze |
||
|
Babesioza umana |
Ixodes scapularis, Ixodes ricinus |
Statele Unite ale Americii, Europa |
Modificarea vegetatiei
Incendiile provocate primavara tarziu pot omori nimfele de Ixodes dammini, reducand astfel riscul expunerii oamenilor la muscaturile de capuse. Incendiile provocate toamna sau primavara devreme pot fi defavorabile; reducand numarul de gazde animale, oamenii sunt mai accesibili atacurilor ixodidelor. Indepartarea gunoaielor care atrag rozatoare (gazde pentru ixodide) ar putea reduce partial prezenta capuselor in apropierea locuintelor. Acelasi efect l-ar putea avea si indepartarea surselor de hrana pentru pasarile ce se hranesc la nivelul solului.
Limitarea accesului ixodidelor la gazdele vertebrate adecvate
In anumite regiuni ixodidele din specia Boophilus microplus au fost reduse numeric sau chiar eradicate prin rotatia gazdelor pe pasuni; astfel, a fost interzisa hranirea vitelor pe pasunile infestate pana la disparitia ixodidelor. O reducere asemanatoare a numarului de indivizi apartinand speciei Ixodes ricinus a fost inregistrata prin rotatia ovinelor (principalele gazde pentru adulti) intre pasuni, pe perioade mai indelungate de timp.
|
Subfamilia / Genul |
Specia |
|
Subfamilia Ixodinae |
||
Genul Ixodes |
Ixodes ricinus (Linné) 1758 Ixodes laguri Olenev, 1931 Ixodes redikorzevi Olenev, 1927 Ixodes apronophorus Schulze, 1924 Ixodes hexagonus Leach 1815 Ixodes crenulatus Koch, 1844 Ixodes rugicollis Schulze & Schlottke 1929 Ixodes vespertilionis Koch, 1844 Ixodes chiropterarum (Babos, Janisch) 1958 Ixodes trianguliceps Birula 1895 |
|
Subfamilia Haemaphysalinae |
||
|
Genul Haemaphysalis |
Haemaphysalis punctata Canestrini & Fanzago 1877 Haemaphysalis sulcata Canestrini & Fanzago 1878 Haemaphysalis otophila Schulze 1918 Haemaphysalis concinna Koch 1844 Haemaphysalis inermis Birula 1895 |
|
Subfamilia Amblyomminae |
||
|
Genul Dermacentor |
Demacentor pictus Hermann 1804 Dermacentor marginatus (Sulzer) 1770 |
|
|
Genul Rhipicephalus |
Rhipicephalus annulatus (Boophilus calcaratus) Birula 1895 Rhipicephalus bursa Canestrini & Fanzago 1877 Rhipicephalus sanguineus Latreille 1806 Rhipicephalus rossicus Yakimoff & Kohl-Yakimova 1911 |
|
|
Genul Hyalomma |
Hyalomma aegyptium (Linné) 1758 Hyalomma plumbeum (Panzer) 1795 Hyalomma scupense Schulze 1919 |
|
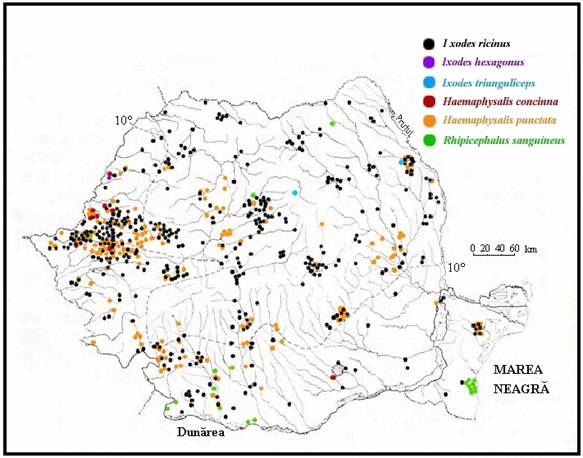
(Coipan & Vladimirescu, 2003, dupa Feider 1965)
Figura 2. Raspandirea geografica a speciilor mai importante de capuse
in Romania.
Controlul chimic
Primele pesticide erau de natura vegetala: nicotina, rotenona si piretrumul. Mai tarziu (dupa al II-lea Razboi Mondial), insecticidele reziduale sintetice ca DDT (diclor-difenil-triclor-etan) si mai putin HBC (hexaclor-benzen) au revolutionat controlul populatiilor vectoare. Pe masura ce rezistenta vectorilor si alti factori au impiedicat utilizarea insecticidelor clorurate, au inceput sa fie folosite alte clase de insecticide. O noua generatie de insecticide, incluzand piretroizii sintetici, Bacillus thuringiensis israelensis (Bti) si reglatorii de crestere a insectelor, a devenit foarte populara (Tabel 13).
Acaricidele: sunt folosite pentru tratarea animalelor domestice si salbatice infestate cu ixodide, precum si a vegetatiei, solului, caselor sau adaposturilor de animale. Animalele domestice pot fi tratate cu acaricide prin imbibare, stropire sau prin atasarea unei zgarde impregnata cu substante acaricide volatile.
In cazul animalelor salbatice se poate folosi metoda materialelor artificiale impregnate cu acaricide. Aceste materiale sunt lasate in camp si folosite ulterior de rozatoare la construirea cuiburilor; astfel sunt omorate toate stadiile de dezvoltare ale ixodidelor aflate in cuiburile acestora.
Acaricidele reziduale pot fi aplicate pe vegetatie, sol si resturile de pe sol, de-a lungul drumurilor, potecilor facute de animale, in curti, pe pasuni sau in alte locuri in care tind sa se adune ixodidele. Aplicarea acaricidelor reziduale pentru controlul genului Ixodes poate fi eficienta pe arii relativ mici, frecventate de om; aplicarea pe arii extinse, de padure sau desis este mult prea costisitoare.
Acaricidele granulare cum sunt carbarilul si diazinonul pot fi folosite pentru controlul ixodidelor active toamna. Controlul continua in primavara urmatoare sau in perioadele de activitate a larvelor si nimfelor, limitand astfel numarul de stadii de dezvoltare gasite pe gazde.
Permetrinul produce moartea rapida a ixodidelor. El poate fi aplicat direct in vizuinile rozatoarelor pentru controlul stadiilor preimaginale.
Aplicarea de carbaril si diazinon toamna, impotriva adultilor activi, reduce foarte mult numarul acestora, atat in perioada imediat urmatoare cat si in primavara urmatoare. Dar aceste aplicatii nu au nici un efect asupra nimfelor. Aplicarea unuia dintre aceste acaricide sau a cloropirifos-ului, realizata la momentul oportun (in mai-iunie) poate fi eficienta si in reducerea numarului de nimfe active (aflate in cautarea unei gazde).
Suprafetele extinse pot fi tratate prin stropiri efectuate din avion sau elicopter. Suprafetele mici pot fi tratate prin stropire de la nivelul solului sau prin aplicarea unor emulsii de insecticide pe baza de apa sau a unor insecticide sub forma de pudra. Efectul acestor stropiri dureaza putin mai mult de o luna, in functie de conditiile zonei stropite si de dimensiunea acesteia.
Rezistenta speciei Boophilus microplus (parazita pe bovine) la anumite acaricide a devenit o problema in ultimul timp si limiteaza eficienta acestor substante.
Rezistenta este definita de WHO (World Health Organization) ca «dezvoltarea capacitatii unei tulpini a unui organism de a tolera doze ale unei substante toxice care sunt letale pentru majoritatea indivizilor unei populatii normale ale aceleiasi specii».
Controlul biologic
Bacteriile. S-a constatat ca bacteriile Cedecea lapagei (Enterobacteriacea) de origine braziliana infecteaza in mod natural pana la 40% din femelele hranite de Boophilus microplus. Aceste bacterii infecteaza si distrug epiteliul vaginal al capuselor. Injectarea a 1 μl de suspensie bacteriana 2×109 sau imersionarea femelelor hranite intr-o asemenea solutie rezulta in 100 si respectiv 39,1% mortalitate in randul femelelor. Descoperirea unor izolate mai virulente sau producerea unor specii mai virulente prin selectie si/sau bioinginerie, combinata cu o formulare optima, poate conduce la obtinerea unor acaricide puternice.
Femelele de Argas persicus hranite, la sprayerea cu toxina rezultata din Bacillus thurengiensis var. kurstaki (1,250 μg/ml), au inregistrat o mortalitate de 100 %. Femelele nehranite sau ouale acestei specii, precum si femelele si ouale de Hyalomma dromedari par sa fie mai putin susceptibile (Samish 2000).
Fungii. Cateva ciuperci entomopatogene sunt asociate in mod natural capuselor iar unele au fost demonstrate a fi extrem de virulente pentru capuse in conditii de laborator. Cele mai intens studiate ciuperci entomopatogene sunt Metarhizium si Beauveria (Deuteromycotina). Ele pot penetra cuticula si sunt adesea virulente pentru toate stadiile de dezvoltare ale capuselor. Cateva experimente in teren au conferit rezultate promitatoare in acest sens. Ciupercile necesita umiditate relativ crescuta pentru germinare si, in aceste conditii, capusele sunt omorate in numai cateva zile.
Nematodele. Nematodele entomopatogene apartinand familiilor Steinernematidae si Heterorhabditidae au fost identificate ca virulente pentru 13 specii de ixodide si doua specii de argaside; o singura specie din cele testate a fost gasita rezistenta la tulpinile respective de nematode. Capusele adulte par sa fie mai susceptibile la nematode decat stadiile preimaginale. Nematodele sunt deja comercializate pentru controlul unor daunatori ai plantelor; totusi, susceptibilitatea lor la umiditate si temperaturi scazute le poate limita utilizarea la anumite nise ecologice. Introducerea de noi tulpini de nematode si formulari ar trebui sa creasca diversitatea niselor ecologice adecvate
Viespile parazite. In prezent, se cunosc sapte specii parazitoide din genul Ixodiphagus dar numai Ixodiphagus hookeri si Ixodiphagus texanus au fost studiate amanuntit in conditii de laborator. De obicei, ele depun ponta in larvele sau nimfele de capuse iar viespile adulte emerjeaza din nimfe la 1-2 luni dupa ce acestea s-au desprins de gazda. In ultima parte a secolului al douazecilea incercarile de introducere a I. hookeri in cateva noi situri fie au esuat fie, daca au reusit, nu au avut nici un efect semnificativ asupra populatiei de capuse. Astfel, au fost descurajate initiativele ulterioare in aceasta directie de biocontrol al capuselor. Este inca neclar de ce viespile au fost ineficiente, daca augmentarea populatiei de parazitoizi ar putea aduce vreo schimbare sau in ce masura ar putea fi mai eficiente alte specii de viespi introduse in noi ecosisteme. Intr-un singur experiment, in Kenya, eliberarea viespilor la intervale de un an in apropierea vitelor infestate cu Amblyomma variegatum, a condus la reducerea populatiei de nimfe atasate de gazde cu pana la 95% in decurs de patru luni de la prima eliberare si la mentinerea acesteia la un nivel scazut timp de cel putin doi ani.
Pasarile. Din aproximativ 50 de specii cunoscute a se hrani cu capuse, cele mai importante sunt Buphagus africanus (yellow billed oxpecker, YBO) si Buphagus erythrorhinchus (red billed oxpecker, RBO) care traiesc in regiuni extinse din Africa. Aceste pasari se hranesc aproape exclusiv cu artropodele parazite ale mamiferelor si in special cu Ixodidæ; de aceea par sa fie foarte importante in biocontrolul capuselor. Buphagus africanus poate consuma zilnic pana la 13.600 nimfe sau 109 femele (Boophilus decoloratus), in timp ce Buphagus erythrorhinchus poate consuma 12.500 nimfe sau 98 femele din aceeasi specie de capuse. Totusi, aceste pasari pot sa mentina deschise cicatricile sau ranile mamiferelor respective, hranindu-se cu bucati de piele sau sange din acestea; astfel, cresc sansele aparitiei unor infectii secundare, inclusiv miaze.
In Brazilia, la investigarea canalului alimentar al unui mic soim (Milvago chimango) au fost gasite in medie 122 capuse apartinand speciei Boophilus microplus, cu un maxim de 289 capuse per pasare. Pe baza acestor date s-a calculat ca o pasare ar putea reduce populatia de capuse de pe o vaca infestata masiv cu aproximativ 13% zilnic. Imbunatatirea habitatului acestor pasari pentru a spori propagarea lor sau introducerea lor in noi situri se poate realiza numai dupa determinarea potentialei lor influente asupra altor animale.
S-a descoperit ca gainile domestice se hranesc intens cu capuse, dupa o perioada de adaptare. Atunci cand gainile au fost lasate sa se hraneasca intr-o crescatorie de vite, 75, 79, si respectiv 48 capuse/ pasare/ ora au fost ingerate dupa una, doua si respectiv trei ore. Pasarile au colectat atat capusele de pe vite, cat si cele libere. Astfel, ar fi eficient sa fie lasate gainile sa se hraneasca in curti in dimineata premergatoare eliberarii vitelor la pascut, pentru reducerea populatiei de capuse.
Controlul imunologic
Ixodidele sunt cei mai importanti vectori ai agentilor patogeni atat la animalele domestice cat si cele salbatice. In ceea ce priveste rolul lor vectorial pentru agentii patogeni umani, ixodidele se situeaza pe locul al doilea dupa culicide (Balashov 1972). In continuare sunt descoperite noi boli transmise de ixodide. Incidenta borreliozei Lyme este in continua crestere. Importanta infectiilor umane si animale cu specii de Ehrlichia este recunoscuta pretutindeni. Mai nou, infectia cu rickettsia Rochalimea henselae este considerata a fi transmisa de ixodide (Lucy et al. 1992).
Microorganismele desfasoara cicluri de dezvoltare foarte complexe in interiorul ixodidelor (Kocan 1986). Dezvoltarea agentilor patogeni, biologia vectorilor si raspunsul gazdelor la infectie si infestare sunt interconectate. Examinarea unuia dintre aceste aspecte fara a le lua in considerare pe celelalte conduce la crearea unei imagini incomplete asupra dinamicii interactiunii gazda-vector-patogen.
Cunostintele referitoare la activarea si dezvoltarea agentilor patogeni in artropodele vectoare, la transmiterea lor, la modularea raspunsului imun al gazdei la agentul patogen si la impactul acestui raspuns imun asupra transmiterii agentului patogen de catre vectori, sunt limitate.
Citokinele, anticorpii si raspunsul imun celular constituie elementele critice in stabilirea unei infectii, evolutia unei boli si in apararea gazdei. In ultimul timp a devenit foarte importanta utilizarea imunitatii induse a gazdei ca strategie de control pentru vectorii hematofagi. Cunoasterea amanuntita a raspunsului imun al gazdei in cazul unei infestari este esentiala pentru obtinerea unor vaccinuri impotriva vectorilor.
Artropodele stimuleaza diferite tipuri de raspuns imun. Studiile initiale asupra imunitatii au vizat reactiile gazdei la nivelul tegumentului unde se hranesc vectorii. Aceste studii au relevat diferite aspecte ale evenimentelor celulare ce se petrec la locul intepaturilor artropodelor hematofage. Investigatiile ulterioare au vizat relatia dintre substantele continute de saliva artropodelor, raspunsul imun al gazdei la acestea si impactul acestor interactiuni asupra transmiterii agentilor patogeni.
In dezvoltarea unui vaccin impotriva ixodidelor trebuie luati in considerare mai multi factori:
cunoasterea detaliata a raspunsului imun al gazdei in cazul infestarii;
utilizarea raspunsului imun umoral si celular al gazdelor rezistente pentru identificarea imunogenelor ce declanseaza raspunsul imun;
caracterizarea procesului de modulare a raspunsului imun al gazdei de catre vector si utilizarea ca tinta a vaccinurilor a acelor molecule responsabile de imunosupresarea gazdei.
Mecanismele imune ale gazdei supresate de catre ectoparaziti sunt cele care au rol in respingerea acestora. Astfel, supresia acelor mecanisme imune faciliteaza supravietuirea ixodidelor. Moleculele imuno-supresive introduse de catre vector ar putea fi considerate ca tinte pentru vaccinuri. Ribeiro si Spielman (1986) au descris procesul de inactivare a analfilatoxinei de catre saliva speciei Ixodes scapularis. Inactivarea anafilatoxinei blocheaza raspunsul imun mediat de complement.
Punctul de plecare pentru crearea unui vaccin impotriva ixodidelor a fost reprezentat de utilizarea omogenatului total de ixodide sau a extractului din glandele salivare ale acestora, insa in ultimul timp atentia s-a concentrat asupra unor imunogene bine definite (Wikel et al 1992). Trager (1939) a indus rezistenta la infestarea cu Dermacentor variabilis prin imunizarea cobailor cu extract total de larve; a fost astfel indusa rezistenta la infestarea ulterioara cu aceeasi specie de ixodide, fiind respinse 38-68% din larvele infestante.
S-a constatat ca cele mai comune elemente tinta ale reactivitatii imunitare a gazdei sunt derivate din tractul digestiv al ixodidelor; alte tinte sunt reprezentate de hormoni, elementele sistemului nervos, cuticula, muschi si proteinele din hemolimfa. Descoperirea unor molecule care sa confere imunitate incrucisata este foarte importanta, un singur vaccin putand oferi protectie impotriva mai multor specii de ixodide.
Protectia individuala
Vestminte protectoare pot oferi protectie la muscaturile diferitelor artropode in cazul in care au o grosime si o textura greu de strabatut de aparatul bucal al acestora.
Culorile deschise atrag, in general, mai putine insecte decat cele inchise. In cazul persoanelor ce desfasoara activitati in natura, hainele ar trebui sa acopere o parte cat mai mare din corp iar pe zonele expuse trebuie aplicati repelenti.
De asemenea si hainele pot fi tratate cu repelenti sau cu insecticide cu actiune rapida din categoria piretroizilor (cum este permetrinul). Aceste insecticide irita artropodele sau le omoara inca inainte de a se hrani. Aplicarea repelentilor si a insecticidelor pe haine este de preferat deoarece reduce riscul reactiilor alergice. In plus, aderenta puternica la fibrele textile face posibila utilizarea unor doze mai mari de repelenti sau insecticide.
Pentru tratarea hainelor sunt preferati piretroizii sintetici fata de repelentii volatili, din mai multe motive:
actioneaza rapid si resping sau indeparteaza artropodele;
au durata de actiune mai lunga si sunt mai rezistenti la conditiile variabile de mediu (soare, ploaie, s.a.);
utilizarea lor este mai placuta, intrucat au un miros discret sau imperceptibil, nu au culoare si nu au consistenta uleioasa;
folosirea lor nu implica riscuri si nu irita pielea, in cazul aplicarii unor doze corecte;
nu afecteaza produsele plastice;
sunt mai ieftini decat repelentii si nu necesita decat aplicari rare si in cantitati mici.
Pe de alta parte, datorita fenomenului de evaporare, hainele tratate cu repelenti volatili confera o mai buna protectie a zonelor expuse (neacoperite). Impregnarea sosetelor si a pantalonilor este eficienta in protectia contra ixodidelor. Pentru tratarea hainelor, vestelor, camasilor cu maneca lunga sau a pantalonilor este recomandata o doza de 1,25 g/m2, iar pentru tratarea camasilor cu maneca scurta o doza de 0,8 g/m2 in cazul permetrinului. In cazul folosirii unui spray pe baza de DEET este recomandata o doza de 20 g/m2, sau aproximativ 70 g de ingredient activ pentru un obiect vestimentar. DEET-ul folosit pentru tratarea hainelor prin inmuiere este disponibil sub forma de mixtura (30% si 95%) cu alcool.
Concentratiile de acaricide folosite in diferite metode de aplicare sunt trecute in Tabelul 13.
Argasidele care se hranesc pe oameni in interiorul casei pot fi controlate cu ajutorul plaselor de pat pentru tantari, impregnate cu piretroizi. Plasele au de obicei un diametru al ochiurilor de 1,2-1,5 mm si sunt confectionate fie din materiale traditionale (in, rafie sau canepa) fie din bumbac sau materiale sintetice (nylon, poliester, polietilena).
Tabel 13 Concentratiile de acaricide folosite in diferite metode de aplicare.
|
Metoda de aplicare |
Concentratia de acaricid |
|
Inmuiere, spalare sau stropire |
Malathion (5%), diclorvos (0,1%), carbaryl (1%), dioxation (0,1%), naled (0,2%), coumafos (1%) |
|
Acaricid sub forma de pudra |
Carbaril (5%), coumaphos (0,5%), malathion (3-5%), triclorfon (1%) |
|
Spray rezidual aplicat pe podele |
Solutii uleioase sau emulsii de DDT (5%), lindan (0,5%), propoxur (1%), bendiocarb (0,25-0,48%), metil pirimifos (1%), diazinon (0,5%), malathion (2%), carbaril (5%), clorpirifos (0,5%) |
|
Stropirea suprafetelor extinse |
Insecticide organofosforice, carbamati, piretroizi |
|
Zgarzi pentru caini si pisici, impregnate cu acaricide |
Diclorvos (20%), propoxur (10%), propetamfos (10%), permetrin (11%) |
Anderson J. F., Mintz E. D., Gadbaw J. J., Magnarelli L. A. Babesia microti, Human Babesiosis, and Borrelia burgdorferi in Connecticut. Journal of Clinical Microbiology, 29: 2779-2783.
Baumgarten B. U., Röllinghoff M., Bogdan C. Prevalence of Borrelia burgdorferi and Granulocytic and Monocytic Ehrlichiae in Ixodes ricinus Ticks from Southern Germany. Journal of Clinical Microbiology, 37: 3448-3451.
Bozeman F. M., Masiello S. A., Williams M. S., Elisberg B. L. Epidemic typhus rickettsiae isolated from flying squirrels. Nature 255(5509): 545-547.
Caruntu F., Angelescu C., Caruntu V., Streinu-Cercel A., Bocarnea C., Panoiu L., Cracea E., Constantinescu S., Balaci L., Vizitiu O. Aspecte clinice si de laborator ale infectiei cu Borrelia burgdorferi in Romania. Viata Medicala 35: 353-357.
Coipan E. C., Vladimirescu Al. - 2003. Ticks (Acari: Ixodidae) and Lyme Disease in Romania. Romanian Journal of Parasitic Diseases 2 (1-2): 39-54.
Cracea E., Constantinescu S., Balaci L., Vizitiu O., Caruntu F., Angelescu C., Caruntu V., Streinu-Cercel A., Bocarnea C., Panoiu L. Lyme borreliosis in Romania. Archives Roumaines de Pathologie Experimentale et Microbiologie 47: 17-21.
Feider Z. Fauna Republicii Populare Romane - Acaromorpha, Suprafamilia Ixodoidea (Capuse). Editura Academiei Republicii Populare Romane, Bucuresti.
Feider, Z., Rauchbach C., Mironescu I. Die Zecken der Rumanischen Volksrepublik. Cesk. Parasit. 2: 71-87.
Fishbein D. B., Dawson J. E. Ehrlichiae. In: Balows A, Hausler WJ, Herrmann KL, Isenberg HD, Shadomy HJ, eds. Manual of Clinical Microbiology. 5th ed. Washington, DC: American Society for Microbiology 1054-1058.
Hoogstral H, Aeschlimann A. Tick-host specificity. Bull Soc Entomol Suisse 55:5-32.
Humair P.F., Péter O., Wallich R., Gern L. Strain variation of Lyme disease spirochetes isolated from Ixodes ricinus ticks and rodents collected in two endemic areas in Switzerland. Journal of Medical Entomology 32: 433-438.
Jenkins A., Kristiansen B-E., Allum A-G., Aakre R. K., Strand L., Kleveland E. J., van de Pol I., Schouls L. Borrelia burgdorferi Sensu Lato and Ehrlichia spp. In Ixodes Ticks from Southern Norway. Journal of Clinical Microbiology 39: 3666-3671.
Kelly P.J., Mason P.R. Serological typing of spotted fever group Rickettsia isolates from Zimbabwe. Journal of Clinical Microbiology (10): 2302-2304.
McDade J.E., Newhouse V.F. Natural History of Rickettsia rickettsii. Annual Reviews of Microbiology 40: 287-309.
Nakao M., Miyamoto K., Fukunaga M. Lyme disease spirochetes in Japan: enzootic transmission cycles in birds, rodents, and Ixodes persulcatus ticks. Journal of Infectious Diseases 170: 878-882
Opdebeeck J. P. 1994. Vaccines against blood-sucking arthropods. Veterinary Parasitolology 54: 205-222
Oprescu A.C. Raspandirea ixodidelor pe teritoriul R.P.R. Analele Academiei R.P.R. - seria stiinte medicale 2: 1-20.
Pirvulescu V. Contribution a l'etude de la distribution geographique des Ixodides en Roumanie. Bulletin de l'Academie de Medecine de Roumanie 9-10: 357-361.
Pop O., Dutschak H., Rosiu N., Pop-Nowak W. Un caz de boala Lyme detectat in Romania. Bacteriologia, Virusologia, Parazitologia, Epidemiologia 40: 245-247.
Ribeiro J. M. C. Vector saliva and its role in parasite transmission. Experimental Parasitology 69: 104-106.
Ribeiro J. M. C. 1989. Role of saliva in tick/host interactions. Experimental and Applied Acarology 7:15-20
Ribeiro J. M. C., Spielman A. Ixodes dammini: salivary anaphylatoxin inactivating activity. Experimental Parasitology 62: 292-297.
Samish M Biocontrol of ticks. Annals of the New York Academy of Sciences 916: 172-178.
Schouls L. M., Van De Pol I., Rijpkema S. G. T., Schot C. S. Detection and Identification of Ehrlichia, Borrelia burgdorferi Sensu Lato, and Bartonella Species in Dutch Ixodes ricinus Ticks. Journal of Clinical Microbiology 37(7): 2215-2222.
Teodorescu I., Toma D. Boli parazitare. Editura Constelatii, Bucuresti.
Trager W. Acquired immunity to ticks. Journal of Parasitology 25:57-78.
PLANSA 56.
Stadiile de dezvoltare ale ciclului de viata la Argasidae
Tabel 14. Speciile mai importante ale genului Argas
|
Speciile |
Lungimea in (mm) a adultilor nehraniti |
Gazda principala |
Boala (patogenul) |
Tipul patogenului transmis |
|
Argas persicus |
f 5,5-11 m 5,5-8 |
gaini |
Spirochetoza pasarilor de curte (Borrelia anserina) |
S |
|
Argas reflexus |
porumbei |
Borrelia anserina |
S |
Capusele "moi" argaside Argas spp. au nevoie de circa 3-36 luni pentru a se maturiza (in functie de temperatura). Exceptie fac larvele, ce sug sangele 3-10 zile, celelalte stadii se hranesc de mai multe ori, scurte perioade de timp (adultii cateva minute, numai noaptea).
PLANSA 57.
Anatomia capuselor
Mascul de Ixodidae: vedere dorsala (A); vedere ventrala (B).
Femela de Ixodidae: vedere dorsala (C); vedere ventrala (D).
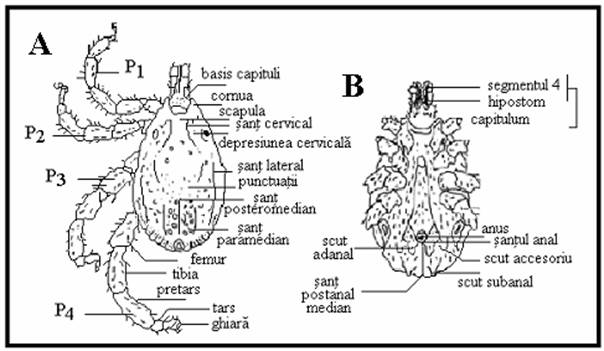
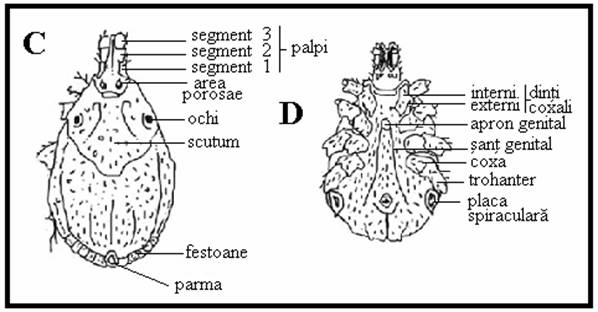
PLANSA 58.
Detalii ale morfologiei externe la capuse I.

(Sursa Internet)
Morfologia externa a capuselor ixodide (A, C, D) si argaside (B) (scanare cu electroni). A, C. Rhipicephalus sanguineus - femela, vedere dorsala (A × 20, C × 75). B. Argas sp. - adult, vedere ventrala; de remarcat ca piesele bucale nu ajung la marginea anterioara a corpului (× 10). D. Ixodes ricinus, tarsul primului picior (× 75).
PLANSA 59.
Detalii ale morfologiei externe la capuse II.
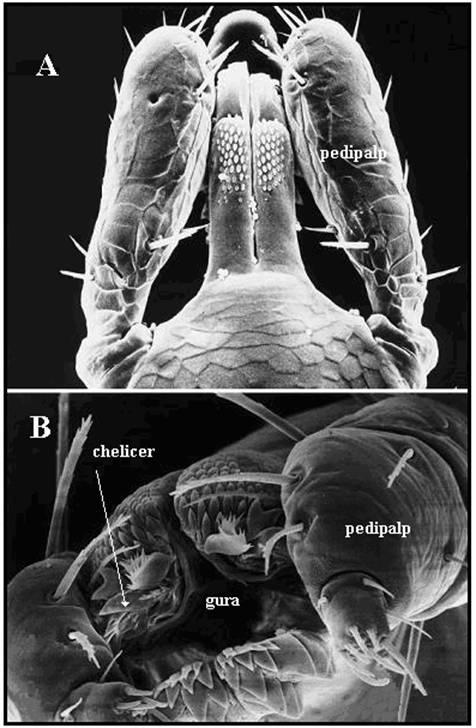
(Sursa Internet)
Piesele bucale
ale capuselor ixodide vazute la microscopul electronic.
A. Ixodes ricinus - vedere dorsala a unei larve (× 150). B.
Amblyomma variegatum - imaginea gurii (× 400).
PLANSA 60.
Specii de capuse intalnite si in Romania

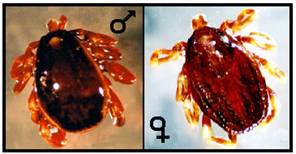
Dermacentor reticulatus Hyalomma marginatum

Ixodes ricinus


Haemaphysalis sp. Rhipicephalus sp.
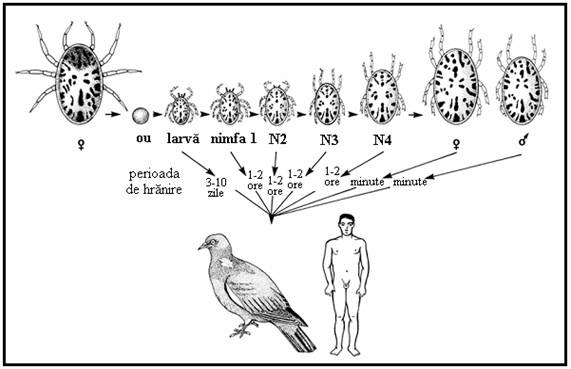
PLANSA 61.
Ciclurile de viata ale capuselor
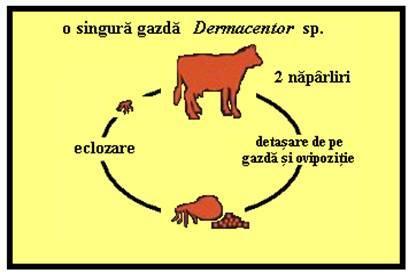
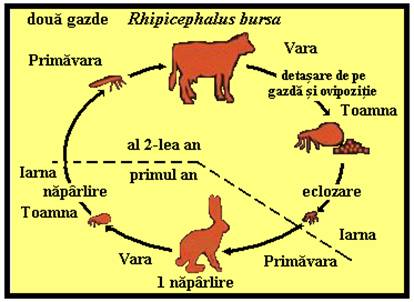
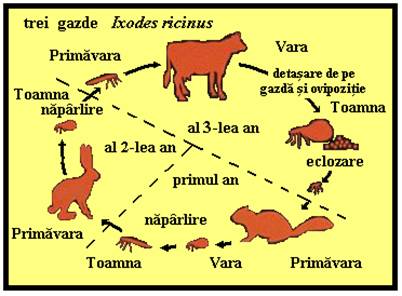
PLANSA 62.
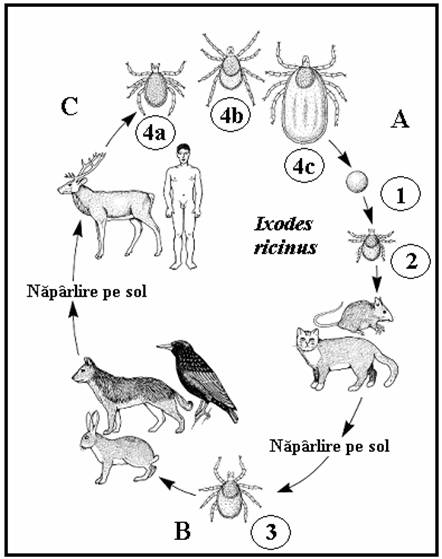
Ciclul de viata la Ixodes ricinus
(Sursa Internet)
Exemplu de capusa cu trei gazde, A-C). 1. Femelele gravide (4c) ating o lungime de circa 1.5 cm, cad pe sol si timp de o luna depun pana la 2000 de oua sferice sau ovoide ce sunt lipite intre ele formand o aglomerare pe sol. 2. Larvele cu sase picioare eclozeaza din oua dupa 3-36 de saptamani (in functie de temperatura) si urca pe frunzele de iarba de unde ataca gazdele aflate in trecere (mici mamifere dar si pasari sau oameni; (A). 3. Dupa hranire larvele cad pe sol si in 5-7 saptamani (uneori 5 luni) se transforma in nimfe cu opt picioare ce nu au inca o deschidere genitala. Nimfele ataca mamiferele mai mari precum si alte gazde (B), sug sange 4-7 zile si cad pe sol, unde dupa 2-8 luni se transforma in adulti maturi sexual (4a = mascul, 4b = femela negravida). Primavara, adultii ataca mamiferele mai mari inclusiv oamenii (C), femelele in special se hranesc 5-14 zile. Intreaga dezvoltare este dependenta de temperatura si in Europa dureaza circa 2-3 ani.
PLANSA 63.
Aspecte din viata capuselor

Stadiile de dezvoltare ale capuselor: de la larva (dreapta) pana la adult (stanga).

Capusa adulta Intepatura capusei
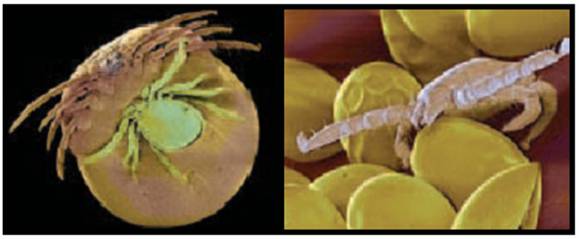
Copulatia Ecloziunea oualor
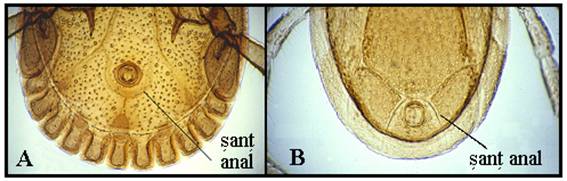
Comparatie intre capusele cu festoane (A) si cele fara festoane (B): genurile non-Ixodes au festoane, in general, si un sant anal posterior fata de anus; membrii genului Ixodes sunt lipsiti de festoane, iar santul anal este anterior fata de anus.
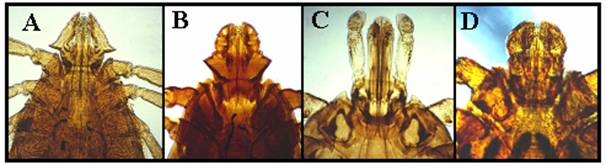
Comparatie intre patru genuri de capuse: Haemaphysalis (A), Portiunea posterioara a segmentelor secundare ale palpilor dilatata lateral; Rhipicephalus (B), structura hexagonala (ascutita lateral) la baza capitulum; Amblyomma (C), segmentele secundare ale palpilor sunt alungite si Dermacentor (D). Toate cele trei segmente ale palpilor par butucanoase.
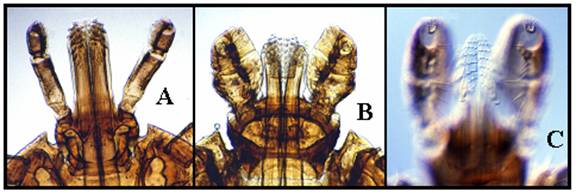
Hipostomul (si dentitia) la trei specii de capuse: A - Amblyomma americanum mascul), B - Dermacentor variabilis (femela), C - Ixodes scapularis mascul
Copyright © 2026 - Toate drepturile rezervate