|







 |  |
|
| |
 | Alimentatie | Asistenta sociala | Frumusete | Medicina | Medicina veterinara | Retete |
INFILTRATUL INFLAMATOR IN CANCERUL MAMAR
COMPONENTELE INFILTRATULUI INFLAMATOR
Daca cercetarile asupra implicarii celulelor inflamatorii in etapele initiale ale cancerizarii si in special in etapa de promotie sunt recente, observatia asocierii variatelor celule inflamatorii in infiltratul peritumoral dateaza inca din secolul al XIX-lea (1891) prin studiile lui Westphal care raporteaza prezenta unei arii dense de celule inflamatorii la periferia tumorilor maligne[58]
Infiltratul inflamator a fost evidentiat atat la nivelul stromei leziunilor epiteliale premaligne, cat si la nivelul stromei tumorilor maligne, dar multitudinea de tipuri celulare care il compun variaza puternic de la un caz la altul. Reactia de reject al celulelor canceroase transplantate este uzual mediata de raspunsul celular nu prin anticorpi activi, totusi reactia anticorpilor pasivi poate fi realizata in terapia tumorala. Prin coloratii speciale au fost evidentiate o magnitudine de celule inflamatorii nonlimfoide care formeaza o patura cu atat mai densa cu cat tumora este mai agresiva - cresterea rapida a tumorilor maligne cu distructia tesuturilor adiacente conduce la un raspuns inflamator rapid. [136]
Este imposibila realizarea unei distinctii morfologice clare intre celulele inflamatorii care apar ca reactii la distructia realizata de tumora si celulele infiltratului peritumoral care au ca efect promovarea cresterii si invaziei tumorale.
Infiltratul inflamator include o multitudine de componente:
# POPULATII LEUCOCITARE - macrophage
granulocite: neutrofile si eozinofile
limfocite
mastocite
# CELULE NK
# FIBROBLASTII SI FIBROCITELE
# CELULE DENDRITICE[76]
Celule mastocite fibroblasti Capilare si cellule de
sustinere macrofage Celule neoplazice
epiteliale
![]()
![]()
![]()
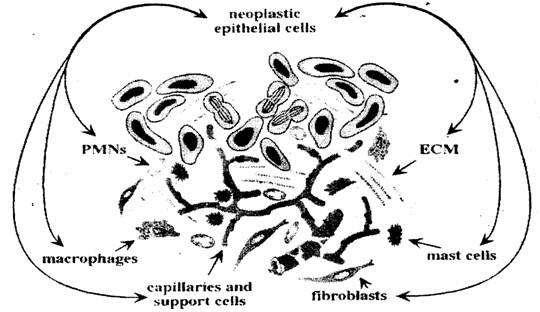
FIG 19 Organizarea tumorii din celule epiteliale maligne si micromediu. ECM - matricea extracelulara contine capilare si suportul lor celular ( pericite, celule musculare netede si fibroblasti ), iar infiltratul inflamator contine macrofage, PMN mastocite si fibroblasti. [76]
1.1. MACROFAGELE
Evaluarea numarului de macrofage in tumora este in mod uzual subestimata din cauza ca aceste celule sunt dificil de recunoscut in sectiunile tisulare prin coloratii uzuale. Macrofagele produc peste 100 de substante biologice dintre care cele mai importante sunt factorii angiogenetici: FGF, VEGF, PDGF, TGFbeta, prostaglandine. De asemenea macrofagele secreta si factori angiogenetici cum ar fi trombospondinul si TNFalfa. [ 136,204]
Macrofagele asociate tumorii reprezinta o componenta proeminenta a populatiei leucocitelor mononucleare in tumorile solide care joaca un rol ambivalent in interrelatia cu celulele tumorale. Recrutarea lor din circulatie se produce prin intermediul chemokinelor tumorale, iar interactiunea cu celulele tumorale are loc la nivelul interfetei tisulare gazda-tumora, regiune adeseori asociata cu o tensiune scazuta a oxigenului. Micromediul tumoral prin citokinele si hipoxia specifica acestuia regleaza captarea, localizarea si functionarea macrofagelor asociate tumorii. [357]
Macrofagele mediaza uciderea celulelor tumorale atat prin citotoxicitate celulara anticorp dependenta (ADCC), cat si prin citotoxicitate directa nonspecifica.[338] Studiile in vitro arata ca citotoxicitatea macrofagelor impotriva celulelor tumorale este mediata prin-o serie de factori solubili dintre care au fost identificati metabolitii oxigenului si nitrogenului [63,338]. In vivo evidentierea acestor mecanisme nu a fost inca realizata pentru carcinoamele umane.Activarea macrofagelor este dependenta de sinteza oxidului nitric(NO), iar inhibitia acestuia duce la depletia potentialului lor citotoxic.Efectul citocid este primar mediat prin NO in virtutea abilitatii lui de a inhiba respiratia mitocondriala prevenind replicarea ADN si denaturand enzimele care contin ionii fier sulf prezente in celulele tinta tumorale[338]
Macrofagele reprezinta prima linie in mecanismul de aparare a organismului care nu necesita reactii de adaptare sau recunoastere imuna.
In cancer macrofagele se comporta ingrat exprimandu-si activitatea atat prin functia obstructiva - prevenirea ancorarii si diseminarii celulelor canceroase, iar simultan exercita si un efect stimulativ asupra cresterii si diseminarii celulelor tumorale.[136,219] Se cunoaste ca tumorile solide sunt constituite din 2 compartimente independente: celulele tumorale si stroma. La nivelul stromei se gasesc vasele de neoformatie, componentele matriciale, celulele responsabile de producerea matricii gel-fibrina si leucocitele inflamatorii. Fagocitele mononucleare reprezinta o componenta importanta constituind pana la jumatate din componentele stromei tumorale. [ 76 ]
De asemenea in micromediul tumoral sunt prezente o multitudine de compusi specifici: care influenteaza activitatea celulelor stromale si in speta a macrofagelor :
Alti produsi antitumorali secretati de macrofage mai nou descoperiti sunt hidrogen peroxidul H2O2 si ligandul FAS(CD95L), esentiali pentru buna functionare a acestora. Aceste componente au impact asupra sistemului imun si influenteaza astfel tropismul tumoral. [ 339]
Cu toate ca relatia intre celulele canceroase si macrofagele care infiltreaza tumora reprezinta inca un subiect controversat, subiectului i se acorda o atentie considerabila. Nivelul macrofagelor recrutat de situsul tumoral este reglat de intensitatea raspunsului imun al gazdei. Procentul macrofagelor asociate tumorii variaza intre 0-80%, cu o medie de 20-30%. Numarul macrofagelor este de cateva ori mai mare la soarecele tumoral fata de soarecele indemn. Tumora maligna induce modificari calitative marcate in subpopulatia TAM si induce adeseori efecte antagoniste functional Influenta locala si sistemica a factorilor derivati tumoral este cruciala in determinarea numarului si naturii TAM. Astfel se explica complexitatea relatiei tumora-macrofage si variabilitatea ei de la o tumora la alta[338]
Macrofagele sunt o sursa de citokine care manipuleaza promptitudinea raspunsului celulelor T. Experimente importante au aratat ca tumorile transplantate la o gazda cu T defective sau celulele NK defective, nu sunt influentate de imunitatea specifica. Astfel, pentru multe neoplazii concluzia a fost ca imunitatea specifica nu joaca rol determinant in infiltrarea tumorii cu macrofage, ci exista un factor derivat din celulele canceroase care pare sa joace un rol major in reglarea nivelului de macrophage la nivelul tumorii [235].
Aceste mecanisme au fost partial elucidate prin descoperirea proteinelor chemotactice monocitare (MCP 1 / CCL 2) care releva chemokinele CC ca factori chemotactici derivati tumoral (TDCF) cu rol major in recrutatrea macrofagelor in situsul tumoral [388]
CCL2 este un membru al familiei chemokinelor CC care leaga receptorul chemokimic CCL2 cu activitate proeminenta asupra monocitelor si limfocitelor T. CCL2 este produs la nivelul situsului tumoral de celulele tumorale, fibroblasti, celulele endoteliale si fagocitele mononucleare. Studiile initiale au aratat ca nivelul expresiei CCL2 este corelat semnificativ cu nivelul de acumulare al macrofagelor in tumora primara atat la soarece cat si la om. Dovezile in vivo, hibridizarea in situ si imunohistochimia au evidentiat productia de CCL2 in melanoame, glioame, cancer ovarian si cancerul mamar. [409]
Studiile in vitro au aratat ca asocierea intre productia de CCL2 si infiltratul macrofagic la nivelul situsului tumoral este corelata cu angiogeneza tumorala atat in melanoame, cat si in cancerul mamar. Pentru carcinoamele mamare s-a aratat ca expresia CCL2 in parenchimul tumoral poate fi corelata cu gradingul histologic al carcinoamelor ductale invasive [388].
Un alt membru al familiei chemokinelor asociat carcinoamelor mamare este CCL5 (RANTES). Studiile au raportat corelatia intre expresia CCL5 in celulele tumorale cu stadii avansate ale bolii sugerand ca CCL5 poate fi implicat in progresia cancerului mamar. Confirmarea acestei ipoteze a fost adusa prin descoperirea recenta a receptorului CCL5 in sectiunile din piesele tumorale ale carcinoamelor mamare operate. Concluzia a fost ca CCL5 in celulele tumorale mamare are ca rezultat migrarea monocitelor in situsul tumoral, activarea protumorigenica, dar si formarea metastazelor, fiind astfel implicat in progresia bolii.[248] Studiile au fost extinse si la cancerul cervical, iar nivelul tisular si plasmatic al CCL5 este corelat cu stadiul bolii atat pentru cancerul mamar, cat si pentru cel cervical.
Macrofagele asociate tumorii se acumuleaza preferential in regiunile precar vascularizate ale tumorii caracterizate printr-o tensiune scazuta a oxigenului. In acord cu aceasta, studiile recente arata ca migrarea macrofagelor este suprimata in conditii de hipoxie, iar macrofagele sunt imobilizate in regiunile avasculare si necrotice ale tumorii datorita hipoxiei caracteristice acesteia. Astfel hipoxia reprezinta un factor de stress care impreuna cu alti parametrii ai micromediului cum ar fi: ph-ul scazut, nivelul scazut al glucozei, nivelul crescut al lactatului, influenteaza profound biologia macrofagelor. In hipoxie macrofagele tumorale sunt nevoite sa-si adapteze metabolismul la caile anaerobiozei si este astfel clarificat cum activitatea proangiogenica a macrofagelor este reglata de hipoxie iar membrii familiei HIF-factorul inductiv al hipoxiei din superfamilia factorilor de transcriptie joaca un rol cheie in expresia genelor care regleaza hipoxia. A fost raportata asocierea intre macrofagele tumorale si expresia factorului HIF-2alfa numai pentru carcinoamele mamare. [374]
O varietate de molecule cu impact posibil asupra angiogenezei s-a aratat ca sunt exprimate de macrofage in conditii de oxigenare scazuta printre acestea amintim: VEGF, TGF-beta, TNF-alfa, MMP-9, bFGF si CXCL8.[429] Mai recent, alti autori au aratat ca hipoxia induce expresia chemokinelor CXCL8 in macrofagele asociate cu cresterea activitatii de legare a ADN-ului la AP1 si HIF1.[177].
Macrofagele asociate tumorii poseda de asemenea capacitatea de a inhiba angiogeneza prin productia de proteine antiangiogenice-inhibitorul activarii plasminogenului tip2 (PAI-2). In timp ce activatorul plasminogenului (uPA) induce factorii angiogenetici implicati in migrarea celulelor endoteliale capilare, expresia inhibitorilor specifici ai plasminogenului (PAI2) este asociata cu un prognostic favorabil. O alta cale prin care macrofagele tumorale inhiba angiogeneza este prin stimularea productiei de trombospondin 1 (TSP1). TSP1 este o proteina a matricei extracelulare produsa de macrofage care inhiba migrarea si proliferarea celulelor endoteliale in vitro si neovascularizatia in vivo prin inducerea PAI [412]
Studiile actuale au aratat ca macrofagele asociate tumorilor mamare reprezinta un factor de prognostic nefavorabil prin favorizarea progresiei si a metastazarii. TAM-urile constituie populatia macrofagica primara cu fenotip M2 polarizata F4/80+/CD206+ care are citotoxicitate redusa pentru celulele tumorale din cauza productiei scazute de NO si citokine proinflamatorii si de asemenea prezinta o capacitate slaba de prezentare a Ag. si supresie efectiva a celulelor T activate [255]
In fapt, aceste macrofage cu fenotip M2 promoveaza proliferarea celulelor tumorale prin secretia de factori de crestere si proangiogenici cum ar fi metaloproteinazele si reglatori ai functiei fibroblastelor din stroma tumorala [237]
S-a descoperit o noua proteina de stress supraexprimata de macrofagele tumorilor mamare care face parte din familia cistein proteazelor denumita LEGUMAIN. Aceasta este o Acid cistein endopeptidaza cu o specificitate restrictiva remarcabila. A fost initial identificata la plante in procesele enzimatice din timpul germinatiei [198,345] si ulterior a fost identificata la mamifere [64,186]. Selectia ei ca tinta pentru terapie are la baza faptul ca genele ce codifica asparagynil endopeptidaza, au fost gasite supraexprimate in tesuturile tumorale, dar au fost gasite absente sau slab exprimate in tesutul normal din care s-a dezvoltat tumora. Acest lucru a fost dovedit pentru tumorile murine si umane [223,263,282].
Supraexpresia LEGUMAIN apare in conditii de stress cum ar fi hipoxia tumorala si este asociata cu stimularea progresiei, angiogenezei si metastazarii. Recent s-a descoperit ca LEGUMAIN este supraexprimata prin TAM-uri in tesutul tumoral mamar utilizand imunohistochimia si profilul expresiei genice. In stroma tumorala in care TAM-urile sunt abundente este gasita si o expresie crescuta a LEGUMAIN. [223]
In contrast, macrofagele cu fenotip M1 ce reprezinta cheia supravegherii imune isi pastreaza functia citotoxica si de prezentare a Ag. nu exprima LEGUMAIN [263]
In consecinta tintirea TAM-urilor ce exprima LEGUMAIN va reduce densitatea lor si va avea ca efect remodelarea micromediului tumoral fara a interfera cu functiile de supraveghere imuna ale macrofagelor M1. Astfel s-ar obtine diminuarea expresiei unor varietati de factori de crestere, proangiogenici si metaloproteinaze eliberate de macrofagele M2 cu efect de stimulare tumorala.[409]
1.2. NEUTROFILE SI ALTE GRANULOCITE
O alta componenta a infiltratului inflamator tumoral este aceea a granulocitelor. Studii in vitro au aratat ca atat macrofagele cat si granulocitele activate de bacterii si produsii lor pot distruge celulele maligne, murine si umane. Neutrofilele care ucid celulele tumorale in vitro sunt sensibilizate prin anticorpi specifici legati de celulele tumorale.[61]
In vivo neutrofilele manifesta activitate antitumorala prin mecanism autoindus de celulele tumorale care in conditii experimentale s-a aratat ca produc nivele crescute de interleukina 4 ( IL4 ) si factorul de stimulare al coloniilor granulocitare ( G-CSF ). Diferentierea fractiunii granulocitare responsabila de inhibitia cresterii celulelor tumorale a fost realizata printr-un studiu la soarece. Tratarea acestuia cu anticorpi care scad numarul granulocitelor duce la blocarea cresterii celulelor tumorale, dar s-a aratat ca neutrofilele nu eozinofilele sunt responsabile de reactia de mediere a inhibitiei cresterii celulelor tumorale in vivo deoarece inhibitia cresterii celulelor tumorale a fost observata si la soarecele cu eozinofilele inactivate dar cu neutrofilele pastrate.[136]
Neutrofilele pot atrage celulele prezentatoare de antigen (prin productia de GM-CSF) in situsul inflamator, ceea ce duce la generarea unui raspuns tumoral specific T mediat al gazdei. Modele experimentale care au studiat efectul depletiei granulocitare au aratat ca acesta duce la esecul dezvoltarii imunitatii specifice antitumorale.
In afara rolului benefic antitumoral s-a aratat ca prezenta granulocitelor poate fi evidentiata si la nivelul infiltartului tumoral in vecinatatea ariilor de proliferare intensa. Astfel, in afara efectului tumoricid si tumoristatic s-a conturat ipoteza participarii granulocitelor si la stimularea cresterii tumorale. Aceasta ipoteza a fost sustinuta de studiile in vivo care au aratat ca depletia granulocitelor prin anticorpi specifici reduce rata cresterii celulelor tumorale transplantate si duce la rejectul tumorii la soarecele eutimic. Pare plauzibil ca efectul neutrofilelor care infiltreaza situsul tumoral sa participe la progresia tumorii si sa nu-si exercite efectul citotoxic si citolitic antitumoral.[81]
Neutrofilele elibereaza si factorul activator plachetar inducand si eliberarea PDGF (factorul de crestere plachetar). Si alte mecanisme sunt implicate in aceste complexe deoarece neutrofilele produc si alte proteine si mediatori. In concluzie neutrofilele in diverse stadii de activare sau prin activitatea lor secretorie au un comportament bivalent atat in sustinerea cresterii tumorale cat si in sustinerea apararii gazdei impotriva tumorilor.[224]
1.3. LIMFOCITELE
Limfocitele fac parte din celulele efectoare implicate in raspunsul imun celular antitumoral.
Rolul limfocitelor T citotoxice
Studierea activitatii celulelor T la pacientii cu cancere primare arata o reactie pozitiva in proportie de aproximativ 70% atat in termenii proliferarii cat si ai lizei. Modelul animal a indicat cu certitudine necesitatea expresiei MHC de clasa I ca triger pentru raspunsul celulelor T prin intermediul receptorului TCR. Rezultatele au fost confirmate si in studiile pe neoplasmele umane. Utilizarea MHC de clasa 1 solubil a dus la identificarea unei populatii de celule T CD8 + specific tumorale la pacientii cu cancer.[338]
MHC-ul tumor specific restrictionat CTL este definit ca efector citotoxic capabil sa lizeze celulele tumorale dar nu si celulele normale autologe.[338]
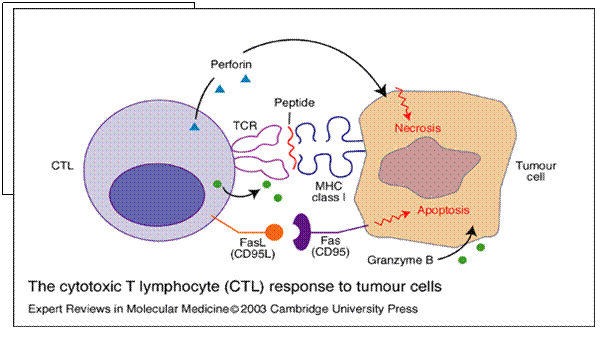
Figura 20. Limfocitul T citotoxic (CTL) si raspunsul antitumoral. [338]
CTL interactioneaza cu celulele tumorale prin intermediul receptorului TCR si complexului major de histocompatibilitate MHC de clasa 1 exprimat pe suprafata celulelor tumorale. Interactiunea implica de asemenea si legarea dintre moleculele de adeziune
-ICAM 1 cu antigenul leucocitar functional asociat (LFA1)
-Fas (CD95) cu Fas ligand (CD95L)
Distructia celulelor tumorale este afectata de eliberarea de perforine, serinesteraze, interferon gama (IFN gama), granzimB si/sau tumor necrozis factor alfa (TNF alfa) din CTL.
LIMFOCITELE Thelper(Th)
Acestea exprima Ag de diferentiere CD4 si joaca un rol crucial in raspunsul antitumoral.
AG tumorale sunt procesate si prezentate celulelor Th prin intermediul moleculelor MHC de clasa 2 ce interactioneaza cu receptorul TCR al celulelor Th. [338] Acestea raspund prin secretia de citokine (IL2,4,10) si activarea altor componente imune.
Th sunt divizate in mai multe categorii in acord cu profilul lor citokinic specific:
a. Th1- produc IFN gama si IL2
b. Th2-produc IL4,5,10
c. Th3-produc IL10 si TGF beta
d. Au mai fost definite Th0 ce produc un spectru mai larg de citokine [338]
Toate aceste subtipuri au rol important in reglarea si controlul initierii si blocarii raspunsului imun.
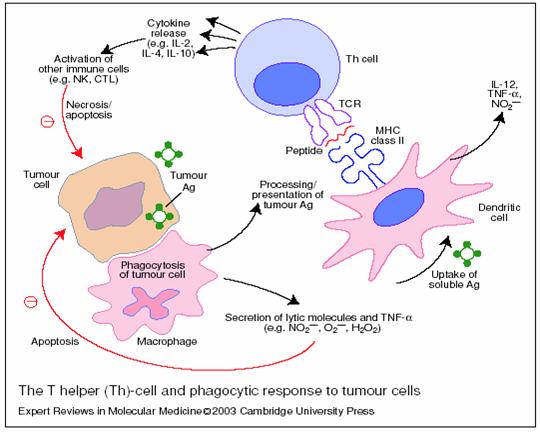
Fig.21 Limfocitele T helper (Th) si raspunsul fagocitic antitoral. [338]
Celulele prezentatoare de antigen (APC) ca macrofagele si celulele dendritice inglobeaza celulele tumorale si produsii lor. Antigenele tumorale sunt procesate si prezentate celulelor Th prin intermediul complexului major de histocompatibilitate (MHC de clasa 2) ce interactioneaza cu celulele Th prin TCR. Celulele Th raspund prin secretia de citokine (IL2, IL4, IL10) ce activeaza alte celule ca NK si TCR. Macrofagele secreta de asemenea molecule litice ca oxidul nitric, superoxid si hidrogen peroxid in aditie cu TNF alfa. Celulele dendritice secreta IL12, TNF alfa si oxid nitric.
Rolul Th in raspunsul imun antitumoral poate fi benefic sau detrimental depinzand de factorii genetici si antigenicitatea specifica fiecarei tumori in parte.
Cercetarile au fost focalizate in special pe raspunsul mediat de CTL deorece majoritatea tumorilor exprima MHC de clasa 1 si pierd expresia MHC de clasa 2. [338]
1. CELULELE NK
Termenul de NK este utilizat pentru a defini celulele citotoxice pentru tintele neoplazice si non-neoplazice in absenta activarii receptorului specific TCR prin antigen.
Celulele NK (natural killer) in particular sunt activate ca celule care ucid celulele canceroase in vitro cat si in vivo. Dovezile experimentale arata ca aceste tipuri de celule elimina celulele canceroase NK sensibilizate injectate intravenos gazdei murine.
La animalele de experienta NK sunt prezente in stadiile precoce de dezvoltare ale tumorilor solide si in stadiile tardive ale metastazarii, dar rolul acestora in diferitele etape ale dezvoltarii cancerului este putin cunoscut [400] La pacientii cu activitate deficienta a NK s-a constatat o rezistenta scazuta la infectii si stimularea procesului de metastazare. Un mecanism prin care NK isi pierde functia citolitica este prin expunere la celulele tinta, acestea induc activarea receptorului CD16. Aceste observatii sugereaza ca CD16 joaca rol activ procesului prin care tintele celulare induc anergia/apoptoza celulelor NK.[338]
NK reprezinta o punte functionala intre raspunsul imun nonspecific precoce si raspunsul subsecvent antigen specific. Activitatea NK este indusa de interferoni: IFN-alfa si IFN-beta, iar citokinele proinflamatorii ( IL 12 ) actioneaza ca factori de crestere stimuland activitatea citotoxica si induc ulterior producerea de interferon gama prin intermediul celulelor NK [338] Putine informatii sunt disponibile pentru recunoasterea structurilor de suprafata a celulelor NK si tintelor celulare. Recent, au fost identificati activatorul receptorului pentru NK in expresia gamadelta si pentru celulele T alfabeta-TCR si liganzii lor corespunzatori MICA si MICB[338] Se cauta pentru tumorile care exprima acesti liganzi mecanismul de control care influenteaza balanta pentru supraexpresie si care favorizeaza toleranta imuna a gazdei pentru tumora maligna.[370,400].
NK ucid celulele tumorale prin urmatoarele mecanisme in care sunt implicate:
In vitro NK pot ucide selectiv celule tumorale care au pierdut antigenele majore de histocompatibilitate de clasa I, iar in vivo a fost observat in timpul progresiei tumorale pierderea alelelor antigenului major de histocompatibilitate de clasa I. Se incearca gasirea modalitatilor de aplicare in practica a functiilor distructive a celulelor NK in asociere cu interleukinele si interferonii.[382]
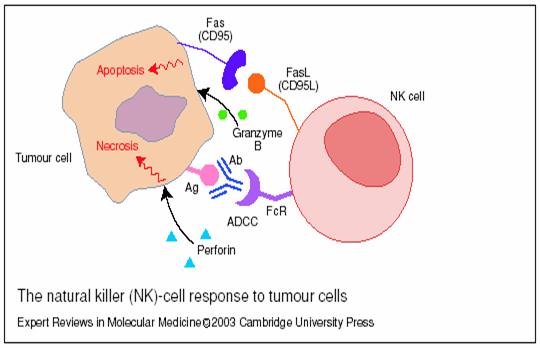
Fig.22 Celulele natural killer(NK) si raspunsul antitumoral [338]
1.5. CELULELE DENDRITICE
Celulele dendritice (DC) functioneaza ca celule prezentatoare de antigen (APC). Prezenta lor in infiltratul inflamator este considerat un factor de prognostic bun, iar interactiunea intre DC si tumora duce la eliberarea de citokine antitumorale (Fig Th) [338,298]
Inhibitia procesului de metastazare tumorala pare a fi dependenta si de intrarea DC in tumora avand ca efect eliberarea oxidului nitric (NO) si a altor citokine inhibitorii. Esecul actiunii DC in tumora este tradus prin abilitatea celulelor tumorale de a produce citokine inhibitorii ale NO [213]
S-a aratat recent ca antigenele ce atrag DC conduc la sensibilizarea directa a celulelor tumorale si stimularea aparitiei raspunsului imun Ag specific. Maturarea si activarea Dc este mediata de interactiunea dintre CD40-DC si CD40L-Th [338]. Studiile aflate in derulare incearca utilizarea potentialului antitumoral al DC activate in terapia cancerului.
1.6. FIBROBLASTELE SI FIBROCITELE
In aria infectiei fibroblastele si fibrocitele reprezinta o sursa semnificativa de cytokine proinflamatorii si imunomodulatorii. Ca raspuns la injurii tisulare aceste celule produc factori de crestere si componente matriciale necesare pentru repararea si remodelarea tesuturilor .
Fibrocitele reprezinta populatia celulelor sentinela care actioneaza in sistemul de aparare precoce pentru repararea tesuturilor si in infectii. De exemplu imediat dupa stimularea prin injurii, fibroblastele raspund rapid prin producerea de IFN-alfa si IFN-beta si de asemenea joaca un rol important in functia de prezentare a antigenului [359]
Fibroblastele si fibrocitele poseda de asemenea un rol critic in controlul cresterii si invaziei celulelor canceroase dovada fiind studiile in vivo care arata ca aceasta subpopulatie reactioneaza prin productia de cytokine si factori de crestere cu actiune asupra tumorii.[194,227]
Ca exemplu, interleukina 6 produsa de fibroblastele dormice este un inhibitor al cresterii celulelor tumorale in stadiile precoce ale dezvoltarii melanoamelor, dar tot aceasta poate induce si o stimulare paracrina a celulelor canceroase in stadii tardive. Un subiect controversat este acela al influentarii fibroblastilor de catre celulele tumorale care pot controla responsivitatea fibroblastilor la factorii de crestere ca si inducerea receptorilor pentru factorii de crestere in fibroblastii stromali.[370]
Familia factorilor de transcriptie REL/NF-kB si reglatorii lor subunitatea I-kB, joaca un rol important in controlul informatiei incluzand cytokine trascriptionale, genele receptorilor citokinelor in fibroblasti si in alte celule ale sistemului imun, dar aceiasi factori si proteine au implicatii majore si in cancer.[39,48,195]
1.7. CITOKINELE
Citokinele sunt reglatori ai activitatii normale si joaca un rol important in raspunsul imun al gazdei impotriva variatelor infectii si a cancerului. Caracterul raspunsului imun antitumoral este semnificativ influentat de activitatea populatiei de celule Thelper si a citokinelor produse de ea. Citokinele majore implicate in raspunsul imun (tabel)
Abilitatea citokinelor de-a produce regresia tumorala este exemplificata in modelul tumoral AK-5 in care IL 2, IL12, IFN gama si TNF alfa au avut un rol important in regresia antitumorala [384,390] La acelasi model animal AK5, s-a aratat ca regresia tumorala a fost mediata primar de celulele NK. NK-urile sunt incapabile sa ucida celulele tumorale pana nu sunt activate de IL2, IL12 si IFN gama.[338]
Nivelul citokinelor in situsul tumoral s-a dovedit a fi de cateva ori mai crescut fata de nivelul lor circulant sugerand astfel rolul lor important in intretinerea limfocitelor infiltrate tumoral si a situsului imun activat antitumoral.
CITOKINELE MAJORE IMPLICATE IN RASPUNSUL IMUN
![]()
![]() Citochine Sursa Tinta Efect in
tinta
Citochine Sursa Tinta Efect in
tinta
TNF alfa Macrofage Macrofage Activare, crescand aderenta
TNF- beta celulele T Granulocite leucocitelor la endoteliu,
Endoteliu producerea proteinelor de
Hypothalamus faza acuta, catabolismul
Celular
IFN-alfa Leucocite Toate celulele Efect antiviral, anti-proliferare
IFN-beta Leucocite inducerea clasei I si II a MHC
Activare
IFN-gama Celulele T Tesut celular Efect antiviral, inducerea
Celulele NK Endoteliu clasei I si II a MHC, activare
M-CSF Macrofage progenitori Stimularea diviziunii si
Endoteliu diferentierii
Fibroblasti
G-CSF Macrofage Progenitori Stimularea diviziunii si
Endoteliu diferentierii
Fibroblasti
Celulele T
GM-CSF Celulele T Progenitori imaturi Crestere
Macrofage Progenitori Diferentiere
Endoteliu Macrofage Activare
MIF Celulele T Macrofage Inhibarea migrarii
Factori Limfocite Leucocite Atractia la locul infectiei
Chemotactici Macrofage sau distrugerii tisulare
Granulocite
TGF-beta Celulele T Celulele T Inhibarea activarii
Macrofage Macrofage Reglarea cresterii
Si altii
![]()
Abrevieri : G-CSF- factorul granulocitar de stimulare a coloniilor; GM-CSF: factorul de stimulare a coloniilor de granulocite-macrofage; IFN: interferon; M-CSF factorul de stimulare a coloniilor de macrofage; MHC: complexul major de histocompatibilitate; MIF: factorul de inhibare a migrarii; NK: distrugatorul natural; TGF-beta: factorul beta de transformare a cresterii; TNF: factorul de necroza tumorala.
![]()
Tabel nr 11 [338]
2 FACTORI REGLATORI ASOCIATI INFLAMATIEI SI CANCERULUI MAMAR
FACTORUL NUCLEAR NF-kB
Factorul nuclear NF-kB face parte din grupa factorilor de transcriptie secventiali specifici fiind cunoscut ca reglator cheie al inflamatiei si initiator al raspunsului imun. Studii recente de inginerie genetica la soarece au indicat clar ca NF-kB are si proprietati organogenetice in cateva tesuturi epiteliale incluzand si glanda mamara. In aditie exista dovezi sugestive asupra implicarii NF-kB in carcinogeneza mamara. Nivele crescute ale NF-kB AND legate au fost detectate atat in liniile celulare ale carcinoamelor mamare in vitro, cat si in vivo in tesutul malign al glandei mamare umane. [48,140]
Factorul de transcriptie NF-kB joaca rol major in procese cheie ale organismului: inflamatie, supravietuirea celulara, transformarea celulara si oncogeneza. La mamifere, familia NF-kB contine 5 variante structurale si functionale:
Rel A ( p 65 )
Rel B
c-Rel
NF-kB1 ( p105, p50 )
NF-kB2 ( p100, p52 )
Toate componentele conserva la nivel inalt domeniul Rel (RHD) si azotul terminal care este responsabil de legarea ADN-ului, dimerizare si asocierea cu proteinele inhibitorii - IkB. Modul de sinteza si proprietatile de transactivare sunt impatite in doua clase
~:prima constituita din Rel A ( p65 ), Rel B si c-Rel care sunt sintetizate in forma lor matura si contin domeniul trasactivarii la nivelul carboxilului terminal;
~ a doua clasa constituita din NF-kB1 si NF-kB2 care sunt sintetizati ca precursori (p105 si respectiv p100), continand N terminal RHD si C terminal in serii repetate[141]
Prin procesele proteolitice este pierdut domeniul C terminal rezultand productia factorilor maturi care leaga ADN-ul - proteina p50 si p52 de la care lipseste domeniul trasactivarii. [60]
Familia proteinelor inhibitorii IkB de asemenea contin 5 membrii distincti la mamifere: IkBalfa, IkBbeta, IkBepsilon ,IkBgama, Bcl-3 si proteinele lor precursoare NF-kB1/p105 si Nf-kB2/p100. Proteinele IkB sunt caracterizate din multiple repetari a domeniului lor C terminal care este asociat cu domeniul terminal RHD si proteinele NF-kB. Aceasta interactiune retine NF-kB la nivel citoplasmatic.[39]
Complexul kinazelor IkB (IKK) a fost identificat ca activator major al NF-kB avand 2 subunitati catalitice: IKKalfa si IKKbeta si o subunitate reglatoare IKKgama cunoscuta si sub denumirea de NEMO.
Ca raspuns la o varietate de stimuli cum ar fi TNFalfa, ligandul CD40 (CD40L) sau LPS (lipopolizaharidele), complexul IKK este activat si proteinele IkB sunt fosforilate in doua serine consevate [195]
Fosforilarea IkB este triggerul care duce la translocarea nucleara a NF-kB si activarea transcriptionala a genelor tinta. Proteinele NF-kB formeaza variat homo si heterodimeri a caror activitate este reglata prin doua cai:
Genele tinta reglate prin NF-kB sunt diverse incluzand: imunoreglatori si gene inflamatorii, gene antiapoptotice, gene de reglare pozitiva a proliferarii celulare si gene care codifica reglatori negativi ai NF-kB. [141]
In forma necuplata, dimerii NF-kB contribuie la tumorigeneza prin cateva mecanisme ca: promotia proliferarii celulare, inhibitia apoptozei, stimularea angiogenezei si cresterea metastazarii. In afara implicarii in raspunsul imun si in inflamatie, factorul NF-kB a fost studiat in glanda mamara aratandu-se ca acesta reprezinta un reglator important in multiplele procese ale dezvoltarii glandei, iar nivelele crescute de NF-kB au fost asociate proceselor moleculare complexe implicate in carcinogeneza glandei mamare [39,48]
Definind mai intai stadiile de dezvoltare ale glandei mamare: embrionic, pubertal al sarcinii, lactatiei si involutiei s-a aratat ca expresia crescuta a NF este asociata stadiilor dinamice cu modificari rapide in glanda mamara [141]
Cele doua subunitati NF-kB Rel A cu p65 si p50 si inhibitorii lor majori IkB sunt exprimate in epiteliul mamar atat la virgine cat si in sarcina si lactatie.
Descoperirile recente care au aratat implicarea NF-kB in dezvoltarea normala a glandei mamare in special prin cresterea proliferarii celulelor epiteliale in timpul sarcinii au asociat si dovezi multiple ale asocierii puternice in tumorigeneza mamara. Studii numeroase au confirmat prezenta nivelelor crescute Nf-kB ADN legat, asociata unei activitati crescute atat in liniile celulare de carcinom mamar in cultura cat si in tesutul cancerului mamar uman primar [48]
Cresterea activitatii kinazice IKK s-a dovedit a fi prezenta atat in liniile celulare de cancer mamar, cat si in tesutul canceros primar la om, iar studiile in vitro au aratat ca inhibitia IKK duce la descresterea activitatii NF-kB in celulele tumorale din cultura.
Mutatiile dominant negative ale IKKbeta blocheaza activarea NF in celulele tumorale epiteliale mamare cu receptori RE negative la animalul de experienta. In acelasi timp reducand potentialul lor tumorigenic prin cresterea sensibilitatii la apoptoza indusa de tratamentul anticanceros [39].
Efectul inhibitor al TGFbeta asupra cresterii celulelor tumorale mamare pare a fi mediat prin inhibitia sau activarea aberanta a activitatii NF-kB, iar supraexpresia Rel A sau c Rel previne acest efect. Luate impreuna aceste studii arata ca activarea NF-kB este asociata cu promovarea cresterii celulare in tumorile mamare iar inhibitia NF-kB deschide o cale posibila in tratarea cancerului mamar.
Nu este clarificat inca care este diferenta intre compozitia dimerilor NF-kB pentru celulele epiteliale mamare normale si celulele mamare carcinomatoase. Pentru celulele epiteliului mamar normal a fost gasita o expresie crescuta a Rel A si p50 si foarte scazuta pentru p52, iar nivele foarte ridicate ale polipeptidelor NF-kB2 (p100 si p52) au fost observate in celulele epiteliale carcinomatoase mamare. Bcl-3 ca unic membru al familiei NF-kB care functioneaza ca si coactivator al transcriptiei este supraexprimat in tesutul tumoral mamar uman fiind asociat cu homodimerul p52 care induce tranzitia de la faza S la G1 prin trasactivarea genei ciclinei D1.
In viitor identificarea tuturor genelor tinta ale NF-kB in celulele epiteliale mamare normale si a cailor de reglare implicate in biologia glandei si cancerul mamar, deschid noi cai pentru terapia moleculara anticanceroasa. Modurile distincte de activare ale NF-kB si prezenta subunitatilor catalitice IKKalfa si IKKbeta sugereaza ca utilizarea inhibitorilor selectivi pentru IKKalfa pot avea un rol de blocare a efectului NF-kB in carcinogeneza mamara fara a cauza imunosupresia sistemica asociata inhibitiei IKKbeta.[39,60]
2.2.FACTORUL DE CRESTERE CSF
CSF este un factor de crestere polipeptidic dimeric actionand la suprafata celulei printr-un receptor tirozikinazic S codificat de protooncogena cfms. Acest complex, receptor-factor de crestere este un important reglator al proliferarii, diferentierii si supravietuirii macrofagelor si progenitorilor sai din maduva osoasa [364]
Expresia CSF 1 este un factor critic al modelului soarecelui transgenic pentru evolutia cancerului mamar. Dovezile experimentale furnizeaza date complete asupra unui subset de celule inflamatorii care sub influenta CSF1 si a micromediului dinamic faciliteaza dezvoltarea tumorilor maligne, cresterea neoplazica si eventual invazia metastatica [76].
Cresterea expresiei CSF1 si cfms a fost asociata unui prognostic nefavorabil in cateva tipuri de cancere epiteliale umane: mamar, ovarian, uterin. De asemenea cresterea expresiei CSF1 a fost corelata cu prezenta unui infiltrat leucocitar in timpul dezvoltarii si progresiei cancerului mamar. Oricum, semnificatia functionala a cresterii CSF1 si infiltratului leucocitar in tesutul neoplazic nu este pe deplin clarificata pana in prezent. Studiile in vivo si in vitro au incercat realizarea unei conexiuni intre expresia CSF1, infiltratul macrofagic si stadiile avansate de evolutie ale cancerului mamar, iar mai recent a fost realizata o corelatie a prezentei acestora cu metastazarea pulmonara in cancerul mamar. Aceste studii au aratat ca celulele inflamatorii potenteaza progresia neoplazica prin intermediul factorilor paracrini fiind la fel de importante in evolutia tumorilor ca si oncogenele sau genele supresoare tumorale. [230]
Prin studiile de inginerie genetica au fost incrucisati soarecii transgenenici care sunt susceptibili sa dezvolte cancer mamar PyMT mice cu soareci avand o mutatie recesiva a genei CSF1 (Csf1 op / Csf1 op mouse). Concluziile studiilor au fost: in absenta CSF1, stadiile precoce ale dezvoltarii neoplazice nu sunt modificate de mutatia genei studiate, dar evolutia spre stadiile avansate si metastazarea pulmonara este semnificativ atenuata prin mutatiile genei CSF1. Diferentele cheie intre soarecele PyMT si soarecele Csf1op/Csf1op nu se manifesta prin capacitatea proliferativa a celulelor epiteliale neoplazice ci prin perturbarea recrutarii macrofagelor mature in tesutul neoplazic in absenta factorului CSF1.[76,364]
Recrutarea macrofagelor peritumoral este intarita prin manifestarea expresiei specifice a CSF1 in epiteliul mamar la soarecele CSF1 nul / PyMT ca si caracteristicile de dezvoltare a tumorii primare si a metastazelor. Studii similare arata ca dezvoltarea insamantarilor pulmonare Lewis este afectata de asemenea la soarecele Csf1op/Csf1op . In ultimul exemplu tumora prezinta si un index mitotic scazut si necroza pronuntata posibil prin diminuarea angiogenezei si afectarea stromei tumorale. Aceste defecte se pot corecta prin tratarea animalului de experienta cu CSF1 recombinant. [60,389]
Impreuna toate aceste experimente genetice prevad o relatie cauzala intre infiltratul inflamator cu macrofage CSF1 dependente si potentialul malign al celulei epiteliale mamare.
MEDIATORI AI INFLAMATIEI PRODUSI DE CELULELE CANCEROASE SI CELULELE INFLAMATORII
.A) SPECII DE OXIGEN REACTIV ( ROS )
In starea de sanatate o mica cantitate de ROS este produsa in timpul proceselor fiziologice celulare, iar aceasta este balansata de antioxidantii celulari. In timpul inflamatiei aceasta balanta este rupta prin cresterea acestor substante.
Leucocitele si celulele tumorale produc intermediari de oxigen reactivi (ROIs), iar la nivelul situsului inflamator exprima cresterea sintezei de oxid nitric (iNOS). Contrabalansarea nivelurilor crescute ale ROS si NO prin produsii antioxidanti nu este suficienta pentru perioade indelungate datorita cresterii productiei endogene sau a secretiei leucocitare a mediatorilor de oxigen reactive. [351]
Este cunoscut ca ROS si NO au un rol dual in carcinogeneza. Concentratiile crescute ale acestora produc peroxidarea lichidelor, fragmentarea ADN-ului si moartea celulei. Nivelele scazute ale ROS si NO care induc semnalizarea primara in celule pot induce si al doilea mesaj intracelular: neovascularizatia si distructia de ADN promutagenice [63].
Intr-un model experimental s-a aratat ca celulele netumorale nemetastatice raspund la inductorii chimici prin cresterea nivelului iNOS si apoptoza, dar celulele netumorale metastatice nu raspund astfel. In cateva studii, nivelele scazute ale productiei NO sunt corelate semnificativ cu gradingul tumoral. Celulele maligne si premaligne care supravietuiesc distructiei NO, vor fi mai rezistente la controlul mediat de gazda si vor avea o crestere mai agresiva. In vivo celulele canceroase transfectate produc nivele crescute de iNOS care ucid celulele transfectate din vecinatate [406]
Cytokine inducerea COX 2 Fibre musculare si tesuturi conexe Specii de oxygen reactive Specii de oxid nitric Lipide reactive si substante metabolice Macrophage Neutrofile Leucocite activate![]()
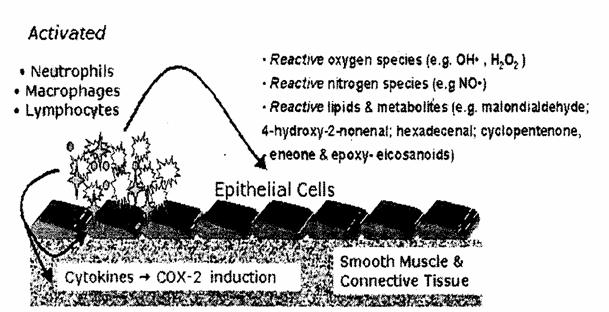
FIG.23 Modelul celular si bazele moleculare ale inflamatiei asociate cancerului.[126]
Aceste studii deschid o noua cale in strategiile terapeutice care vizeaza direct eliminarea tumorii primare si a metastazelor.
B)PRODUSII DE REACTIE AI CICLOOXIGENAZEI
Numeroase investigatii au stabilit importanta medicala a izoenzimelor: ciclooxigenazele 1 si 2 (COX1 si COX2), datele clinice si experimentale arata ca expresia genelor COX2 guverneaza procesele fiziologice si patologice celulare . In neoplazii modificarea expresiei COX 2 poate aparea distict fata de inflamatii.
Cateva tipuri de tesuturi neoplazice sau preneoplazice supraexprima COX2 comparativ cu tesutul adiacent neafectat de cancer. In toate aceste tesuturi expresia COX 2 apare cauzal nu incidental, iar expresia nu este asociata in mod obligatoriu unui proces inflamator preexistent.[136]
In procesele de aparare ale organismului bacteriile si produsii lor cum ar fi lipopolizaharidele (LPS), factorii de crestere si promotorii tumorali chimici, induc ciclooxigenaza 2 (COX2 altfel numita prostaglandin sintetaza) in macrofage si in celulele parenchimatoase. Aceasta enzima este supraexprimata si in variate tumori umane.
![]() Organ / Sistem Expresia COX2 in situs tumoral Nr. Subiecti
Organ / Sistem Expresia COX2 in situs tumoral Nr. Subiecti
![]() Carcinom de colon Proteina COX2 > tesut neafectat 5 / 5
Carcinom de colon Proteina COX2 > tesut neafectat 5 / 5
Adenocarcinom colon Proteina COX2 > tesut neafectat 5 / 7
Tumori mamare Proteina COX2 > tesut neafectat 11/20
Tmori pulmonare Proteina COX2 > tesut neafectat 18/20
Tumori gastrice Proteina COX2 > tesut neafectat 73/104
Carcinom scuamos COX2 mARN de 150 ori > tesut neafectat al
de cap si gat voluntarilor sanatosi
![]() Cancer pancreatic COX2 mARN de 60 de ori > tesut neafectat 9 / 10
Cancer pancreatic COX2 mARN de 60 de ori > tesut neafectat 9 / 10
Tabel nr12 Afectiuni maligne care modifica expresia COX2
Este dovedit ca produsii rectiei COX 2 contribuie la carcinogeneza prin generarea produsilor intermediari care sunt responsabili de mutageneza si activeaza diversi carcinogeni.
In timpul productiei de prostaglandine din acidul arahidonic oxigenul este incorporat in producerea prostaglandinei G2. Aceasta este un intermediar care va fi redus prin activitatea peroxidazica a COX2 la prostaglandina H2. In timpul acestei reactii sunt produsi radicalii liberi oxigeni derivati care cauzeaza reactia oxidativa a ADN-ului. Prostaglandina H2 este convertita in diferite prostaglandine terminale si in aldehida malonica care este mutagen direct al formarii ADN . Chiar daca actiunea radicalilor liberi oxigen derivati este la nivelul lanturilor scurte extreme aceasta sugereaza ca celulele inflamatorii pot cauza distructii ADN oxidative ori mutatii in celulele parenchimatoase de vecinatate [351]
Exista cateva mecanisme aditionale prin care produsii reactiilor COX2 pot contribui la procesele tumorale. Prostaglandinele E si alti produsi ai caii COX2 moduleaza transductia semnalului in variate tipuri celulare incluzand si limfocitele iar PGE poate stimula direct cresterea celulelor epiteliale prin mecanism autocrin ori prin mecanism paracrin (PGE este eliberata preferential in celulele inflamatorii interstitiale).
Supraexpresia COX 2 de asemenea poate descreste apoptoza prin cresterea expresiei BCL2 si reducerea expresiei receptorului TGF beta. Prelungirea supravietuirii celulare rezultata din inhibitia apoptozei creste frecvent aparitia mutatiilor favorizand carcinogeneza [384]
Supraexpresia COX2 duce de asemenea la cresterea productiei metaloproteinazelor accelerand astfel degradarea matricii extracelulare si crescand potentialul invaziv si metastatic al celulelor tumorale. PGE au un rol clar stabilit in activitatea angiogenica si in cresterea eliberarii de eicosanoizi care formeaza suportul neovascularizatiei necesar cresterii invaziei si metastazarii canceroase. [136]
Analiza tuturor studiilor existente arata o frecventa a supraexpresiei COX2 de 43% pentru cancerul mamar invaziv si de 63% pentru carcinomul ductal in situ.
Dovezile epidemiologice, studiile in vitro si in vivo pe modelul animal sugereaza ca expresia COX2 joaca un rol important atat in initierea cancerului mamar cat si in progresie. Prostaglandinele E2 ca mediator major descendent a COX2, promoveaza proliferarea celulara si angiogeneza, induce rezistenta celulelor la apoptoza, creste invazivitatea si moduleaza imunosupresia.
Copyright © 2025 - Toate drepturile rezervate