|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
TESUTUL EPITELIAL SECRETOR EXOCRIN
In cursul histogenezei, in diferite puncte ale unui epiteliu de suprafata, prin proliferarea celulelor bazale rezulta un "mugure" celular (o masa celulara) ce deprima membrana bazala si patrunde tot mai adanc in tesuturile subjacente. Progresiv, la extremitatea distala a mugurelui, celulele constituente sufera un proces de diferentiere, caracterizat prin dezvoltarea organitelor si enzimelor implicate in procesul de sinteza. Acest grup de celule diferentiate va forma componenta secretoare a epiteliului exocrin, denumita adenomer, si va dezvolta in centrul sau un spatiu in care produsul de sinteza va fi eliminat. Celulele epiteliale care fac legatura intre adenomer si suprafata epiteliala de origine se vor transforma progresiv dintr-o masa celulara compacta intr-o structura tubulara cu un lumen bine definit, generand astfel componenta de eliminare sau sistemul tubular (canalicular) al epiteliului secretor. Ambele componente - adenomerul si sistemul tubular - raman separate de tesuturile vecine printr-o membrana bazala, in continuitate directa cu membrana bazala a epiteliului de suprafata.
Ca si in cazul tesutului epitelial de tapetare, in structurile tesutului epitelial secretor exocrin se pot deosebi cateva tipuri celulare ce intra in structura adenomerelor si, respectiv, in structura canalelor.
Clasificarea glandelor exocrine se face pe diferite criterii: numarul de celule secretoare, pozitia lor fata de epiteliul de tapetare, histoarhitectonica adenomerelor si a canalelor, tipurile celulare in raport de natura chimica a produsului de secretie, modalitatile de eliminare.
PARTICULARITATI CITOLOGICE LA NIVELUL EPITELIULUI EXOCRIN
Celulele epiteliale care realizeaza structurile epiteliului exocrin prezinta anumite particularitati in raport cu localizarea si, respectiv, functia lor, motiv pentru care necesita o prezentare detaliata. Cu toate ca, din punct de vedere strict citologic, ele pot avea forme variate (cubice, piramidale, cilindrice), exista o serie de caractere comune, legate de procesul de sinteza si secretie a materialului elaborat, care permit incadrarea in patru grupe mari: celule sintetizante de polipeptide-proteine, celule sintetizante de glicoproteine, celule sintetizante de lipide si celule transportoare de ioni. O grupa aparte este constituita de celulele mioepiteliale - deosebite sub raport functional, dar descrise aici, datorita existentei lor in unele tipuri de adenomere.
Celula sintetizanta de polipeptide-proteine
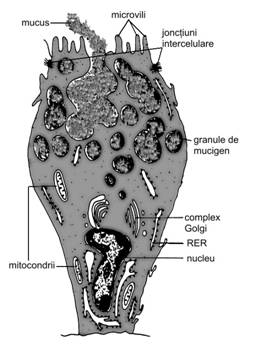
Celula sintetizanta de polipeptide-proteine este denumita si celula zimogena, deoarece produsul elaborat apare sub forma unor granule denumite, in histologia clasica, granule de zimogen. Structura acestui tip de celula este consecinta dezvoltarii importante a organitelor implicate in sinteza si eliminarea de polipeptide sau proteine, cu proprietati enzimatice. Prezenta sa este specifica pentru adenomerele glandelor salivare mixte sau de tip seros (parotida) si ai pancreasului.
In microscopia optica, citoplasma celulara este evident bazofila mai ales in cele doua treimi bazale, cu o zona ceva mai palida, supranucleara. Treimea apicala contine granulele de zimogen, fine, cu un diametru de pana la 1-1,5 μm diametru si cu o tinctorialitate variabila (in raport cu natura chimica a produsului de sinteza); numarul acestora variaza in raport de etapa functionala in care a fost surprinsa celula. Nucleul rotund, cu cromatina in special dispersata si doar cativa grunji de cromatina condensata la nivelul membranei nucleare, este plasat in treimea bazala. In centrul nucleului se gaseste un nucleol proeminent sau in rare cazuri chiar doi nucleoli.
Microscopia electronica releva o
evidenta polarizare, cu un pol apical cu microvilozitati diferit
dezvoltate. Polul bazal nu prezinta un labirint bazal deosebit de
dezvoltat. Se observa o mare cantitate de RER sub forma de cisterne
sau pachete de cisterne, printre care apar numerosi ribozomi liberi
si polizomi. Printre cisternele RER se gasesc mitocondrii in general
alungite, cu creste bine dezvoltate. Supranuclear exista un complex Golgi
bine dezvoltat, organizat cu fata cis spre RER si fata trans
inspre polul apical; cele doua fete sunt unite printr-o retea
tubulara interconectata.
In vecinatatea extremitatilor cisternelor si a fetei
trans se remarca numeroase vezicule de transport, delimitate de o unitate
membranara, cu un continut omogen sau fin granular si o
densitate electronica evidenta. In vecinatatea polului apical se
gasesc complexe jonctionale (ocludente, aderente, desmozomi). De-a
lungul fetelor laterale apar apoi, la oarecare distanta unele de
altele, mai ales jonctiuni aderente.
Din punct de vedere functional, celula produce prin sinteza proteine necesare mentinerii integritatii moleculare a sistemului membranar celular, polipeptide proteine ca produs de secretie ce urmeaza a fi eliminat si proteine care raman stocate in lizozomi. In distributia si destinatia produsilor de sinteza, complexul Golgi joaca un important rol de triaj. Functionalitatea celulei se realizeaza in trei etape importante: (i) captarea precursorilor (aminoacizi) din mediul extracelular, (ii) asamblarea acestora sub forma de polipeptide sau proteine, dupa tiparul sintezei proteice, (iii) stocarea si concentrarea in vacuole golgiene si evacuarea acestora prin polul apical.
Captarea precursorilor
se realizeaza prin polul bazal; printr-un proces activ de punere in
functie a unor pompe cu consum energetic se asigura selectia aminoacizilor necesari. Asamblarea acestora se realizeaza pe fata citoplasmatica
a RER. ARN-ul mesager secundar pentru proteinele secretoare poseda o
secventa particulara de nucleotide ce formeaza codonul
semnal, secventa ce lipseste din structura ARN-ului mesager
secundar pentru proteinele proprii. Odata peptidul semnal asamblat, el se
va continua cu noi aminoacizi atasati, corespunzatori
viitoarelor molecule componente ale secretiei. Peptidul semnal se
leaga cu o particula semnal receptor, care reprezinta adaptorul
ce va permite cuplarea ribozomului si a peptidului semnal la receptorul
ribozomal al membranei RER. Contactul complexului
receptor-semnal - secventa-semnal induce o agregare particulara
a membranei, cu formarea unui por ce va permite translocarea polipeptidului in
interiorul RER. Odata cu trecerea extremitatii carboxiterminale
a polipeptidului prin por, se activeaza o semnal-peptidaza de pe
fata interna, separand peptidul semnal de polipeptidul asamblat;
astfel, acesta devine liber in lumenul RER. In acest moment, prin
detasarea ribozomului, porul se inchide. In
lumenul RER polipeptidul cunoaste modificari posttranslationale,
alunecand in zona REN, unde va fi incorporat in vezicule de transport,
ce vor migra spre fata cis a complexului
Golgi. Din extremitatile saculare ale complexului Golgi si de pe fata trans se desprind
apoi vezicule care, sub actiunea microfilamentelor si microtubulilor,
vor fi dirijate spre polul apical in vederea eliminarii produsului de sinteza. Dupa cum am
mentionat anterior, etapa golgiana pare a juca rolul decisiv in
privinta distributiei materialului sintetizat pentru uz celular sau
pentru export.
Celula sintetizanta de glicoproteine
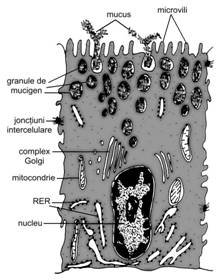
Celula sintetizanta de glicoproteine este cunoscuta, conform histologiei clasice, ca celula mucigena, intrucat produce un material vascos numit, in mod traditional, mucus. Prezenta sa este specifica pentru adenomerele glandelor salivare mixte sau de tip mucos, ai glandelor pilorice si endocervicale.
In fapt, exista trei tipuri distincte de celule mucigene: (i) cu pol mucos deschis (de exemplu, celula caliciforma din epiteliul intestinal); (ii) cu pol mucos inchis (de exemplu, celula din acinii glandelor salivare); (iii) cu pol mucos inchis de tip mucoid (de exemplu, celula epiteliala din epiteliul de tapetare gastric, mentionata anterior in subcapitolul 1 - Tesutul epitelial de tapetare, care are deopotriva rol de protectie si rol secretor sau celula epiteliala din glandele Brunner duodenale). Ne vom referi, in continuare, la celula cu pol mucos inchis.
Caracteristicile morfologice ale acestui tip celular sunt oarecum asemanatoare cu ale celulei zimogenice, existand insa si unele particularitati.
In microscopia optica, prin tehnici de rutina, citoplasma bazofila este cantitativ redusa, restransa in treimea bazala, in jurul nucleului. Cele doua treimi apicale au un aspect foarte palid, pana la incolor, datorita aglomerarii granulelor de mucus; zona aceasta apare intens colorata prin tehnici histochimice (mucicarmin, PAS, albastru alcian). Nucleul, de forma aplatizata, este situat paralel cu polul bazal, in imediata vecinatate a membranei plasmatice; cromatina este mai putin dispersata, iar nucleolul mai putin proeminent, comparativ cu celula zimogena.
Microscopia electronica (fig. I.2.1) evidentiaza tot o celula polarizata, cu aceleasi caracteristici ale polului apical, ale polului bazal si a prezentei si distributiei jonctiunilor. RER este extrem de bine dezvoltat in zona bazala, iar complexul Golgi - supranuclear. Etapele de realizare a produsului de secretie sunt similare cu cele prezentate anterior la celula zimogena, cu diferenta ca aici are loc un intens proces de glicozilare sub actiunea unei glicozil-transferaze.
Procesul de glicozilare incepe in RER prin aditionarea de glucide, mai ales la extremitatile aminolaterale ale asparaginei, si se desavarseste in complexul Golgi, in special in zona trans. In cazul sintezei de glicoproteine sulfatate, adaugarea radicalilor sulfat are loc tot in complexul Golgi, sub actiunea unei sulfo-transferaze. Tot complexul Golgi este implicat si intr-un proces de concentrare, ce poate atinge pana la 25% din masa materialului sintetizat. Ulterior, ca si in cazul celulelor zimogene, se desprind vacuole mici care, prin coalescenta, vor genera vacuole secretoare mari - ce se traduc optic prin prezenta mucusului in cele doua treimi apicale ale celulei mucigene.
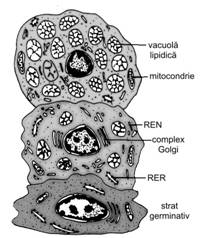
Celula sintetizanta de lipide (sebum)
Celula sintetizanta de lipide este o celula componenta exclusiv a glandei sebacee, anexa glandulara cutanata. Structura sa variaza in raport cu gradul de diferentiere si maturizare, stadii diferite putand fi observate, in microscopie optica si electronica, de la periferie catre centru si zona de eliminare (fig. I.2.2).
Initial, celula putin diferentiata, numita celula bazala, se incarca progresiv cu vacuole lipidice, care cresc in volum, fara insa a fuziona. Urmatoarea etapa este reprezentata de celula intermediara, de forma aproximativ poliedrica, nucleu central si citoplasma acidofila, cu aspect spumos, spongios, datorat incluziilor lipidice. Acest aspect este comparat, pe preparatele de microscopie optica colorate conventional, cu sticla pisata sau cu imaginea unui burete, deoarece picaturile lipidice apar complet incolore, datorita dizolvarii lor de catre solventii organici utilizati in realizarea sectiunilor. Tehnicile speciale pentru lipide permit, insa, conservarea si evidentierea lor. In microscopie electronica, celula intermediara contine toate organitele celulare, in special RER, mitocondrii si mai putin REN. Pe masura acumularii lipidelor, volumul celulei creste, organitele diminua si citoplasma este progresiv inlocuita cu numeroase incluzii lipidice, accentuand aspectul spumos descris anterior. Acest fapt se explica prin existenta si persistenta citoscheletului, format din numeroase tonofilamente, ce impiedica coalescenta incluziilor lipidice.
In stadiile mature
ale celulei, organitele celulare dispar complet, iar nucleii sufera
modificari degenerative (cariopicnoza, cariorexis, carioliza).
O astfel de celula va fi eliminata in totalitatea ei si, prin
distrugerea membranei plasmatice, lipidele vor fi eliminate pe suprafata
tesutului epitelial.
Trebuie mentionat faptul ca nu toate celulele provenite din stratul bazal, germinativ, parcurg aceste etape. Unele dintre ele nu se incarca cu incluziuni lipidice, ci cu keratina, constituind celule de sustinere, dispuse in travee subtiri, cloazonate, intre celulele sintetizante de lipide.
|
|
|
Fig. I.
Celula cu pol mucos inchis - ultrastructura |
|
|
|
Fig. I.2.2. Celula
sintetizanta de lipide - ultrastructura |
Celula transportoare de ioni
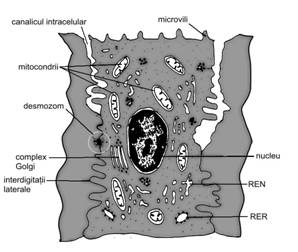
Celula transportoare de ioni este o celula capabila de a efectua un transport activ de ioni contra unui gradient de concentratie. Prezenta sa este specifica in glandele sudoripare, in mucoasa gastrica (celula parietala producatoare de HCl), precum si in unele segmente ale canalelor de excretie a glandelor exocrine. Forma celulei variaza in functie de glanda careia ii apartine. Pentru glandele sudoripare, exista doua tipuri de astfel de celule, incat se diferentiaza doua varietati de glande: ecrine (adevarate, localizate pe toata suprafata corpului) si apocrine (denumire improprie, localizate in axila si in regiunile organelor genitale). Prezentam, in continuare, in detaliu, celula transportoare de ioni, tipica pentru glandele sudoripare ecrine.
In microscopie optica se caracterizeaza printr-o citoplasma acidofila (motiv pentru care este denumita si celula clara), nucleu mai mult sau mai putin central, cu cromatina omogena, fara nucleol vizibil. Adesea este aparent aspectul de striatii al polului bazal.
Microscopia electronica (fig. I.2.3) releva o polarizare manifestata in special printr-un labirint bazal dezvoltat si microvilozitati apicale. Membrana apicala contine enzime (ATP-aza), iar sectorul bazolateral, numeroase pompe ionice. Fetele laterale prezinta numeroase pliuri care intervin in absorbtia apei si electrolitilor. Organitele celulare sunt reprezentate de REN dezvoltat si numeroase mitocondrii.
|
|
|
Fig.
I.2.3. Celula transportoare de ioni -
ultrastructura |
Celula mioepiteliala
Celula mioepiteliala este un tip celular particular, ce apare in structura adenomerelor si a canalelor glandelor exocrine de origine ectodermica - exceptie facand glanda sebacee.
In microscopie optica, pe sectiunile histologice, ea apare ca un nucleu turtit sau bastoniform plasat din loc in loc intre polul bazal al celulelor secretoare si membrana bazala.
Electrono-microscopic s-a constatat ca celula are un corp celular mult aplatizat, din care se desprind prelungiri filiforme fine care se invagineaza printre celulele secretoare, pana la nivelul complexelor jonctionale apicale ale acestora. Astfel, aceste prelungiri "imbratiseaza" celulele secretoare. Corpul celular si prelungirile realizeaza jonctiuni desmozomale cu celulele adiacente. Citoplasma este bogata in filamente contractile (actina si miozina), dispuse asemanator fibrei musculare netede; sunt prezente si filamente intermediare de desmina, specifice tesutului muscular.
Functionalitatea celulei mioepiteliale se gaseste sub control neurovegetativ si hormonal, ceea ce face posibila interventia in procesul de expulzare a granulelor de secretie.
CICLUL SECRETOR
Durata in timp a intregului proces de secretie, de la captarea precursorilor si pana la eliminarea produsului de secretie, formeaza un ciclu secretor.
Acesta este variabil dupa tipul celular si functionalitatea unei glande. De exemplu, celula producatoare de lipide din glanda sebacee realizeaza un singur ciclu secretor, a carui durata este egala cu ciclul celular.
Alte celule secretoare (de exemplu, celula caliciforma) realizeaza doua-trei cicluri secretoare intr-un ciclu celular. In majoritatea glandelor exocrine, numarul de cicluri secretoare este relativ mare, in cadrul unui ciclu celular.
Etapele unui ciclu secretor sunt:
captarea precursorilor, care se face la nivelul polului bazal prin intermediul difuziunii active (canale si pompe), mai rar prin endocitoza mediata de receptori;
sinteza materialului, mecanismul fiind variabil in raport cu natura chimica a produsului de secretie;
desavarsirea compozitiei de secretie, care incepe in complexul Golgi si continua in granulele formate, implicand in special concentrarea si/sau proteoliza unui produs inactiv intr-un produs activ sau a unui precursor in peptide diferite;
transportul
produsilor de secretie, realizat
prin intermediul microfilamentelor si microtubulilor (in cazul granulelor
de secretie) sau prin enzime transportatoare membranare (de exemplu,
ATP-aza);
eliminarea produsului de secretie
In prezent se accepta existenta a doua tipuri de ciclu secretor, care au fost numite tipul secretor constitutiv si tipul secretor reglat.
Tipul secretor constitutiv este reprezentat printr-o eliminare continua, pe masura sintezelor intracelulare. Acest tip apare in special in celule nespecializate in secretie, de exemplu in cele care realizeaza eliminarea de proteoglicani si glicoproteine ale matricei extracelulare.
Tipul secretor reglat, specific celulelor secretoare specializate, permite ca eliminarea sa se realizeze discontinuu, ca urmare a unui semnal molecular (hormon neuromediator) care reprezinta ligandul specific fata de un receptor. Se declanseaza astfel o cascada de procese biochimice, care au drept rezultat cresterea calciului citoplasmatic si a unor substante numite generic fusogene, ce contribuie la eliminare. Celulele secretoare cu tip de secretie reglata au proprietatea de a stoca produsii de sinteza in granule de secretie (vacuole golgiene). Prin fuziunea acestora cu membrana plasmatica se realizeaza, pe de o parte, eliminarea produsului de secretie si, pe de alta parte, un reciclaj al membranei plasmatice, prin adaosul la aceasta a moleculelor constitutive din membranele vacuolelor golgiene.
|
Observatie |
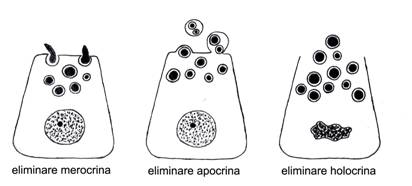
MODALITATI DE ELIMINARE A PRODUSULUI DE SECRETIE
Produsul de secretie elaborat la nivelul glandelor exocrine poate fi eliminat in trei modalitati: merocrina, holocrina si apocrina (holomerocrina) (fig. I.2.4).
Eliminarea merocrina se caracterizeaza prin faptul ca nu afecteaza membrana celulara, mentinand integritatea acesteia (respectiv, integritatea polului apical). S-a presupus ca se realizeaza fie prin difuziune, fie prin traversarea granulelor de secretie integre, la nivelul unor discontinuitati membranare tranzitorii.
Microscopia electronica a demonstrat ca acest tip de eliminare corespunde fenomenului de exocitoza.
|
|
|
Fig. I.2.4.
Modalitati de eliminare a produsului de secretie exocrin |
Eliminarea holocrina consta in expulzarea unei celule glandulare, mai mult sau mai putin modificate, in totalitatea sa. Exemplul ilustrativ clasic pentru aceasta modalitate este cel al glandei sebacee, ce se elimina in totalitate la suprafata epidermului. Prin analogie, considerand spermatozoizii ca celule produse in tubii seminiferi si eliminate de-a lungul unor cai in exteriorul organismului, fenomenul - similar unei secretii exocrine - poate fi considerat de tip holocrin.
Eliminarea apocrina sau holomerocrina a fost considerata initial ca o forma intermediara, in care eliberarea produsului de secretie se realizeaza cu o pierdere a polului apical al celulei. Aceasta modalitate este specifica celulelor epiteliale secretoare de la nivelul glandei mamare.
Microscopia electronica a dovedit ulterior ca substantele proteice si solubile din secretia lactata se elimina prin exocitoza. Pe de alta parte, incluziile lipidice ajunse la polul apical sunt inconjurate de un strat proteic citoplasmatic foarte subtire, care realizeaza fuziunea cu membrana apicala. Acest complex foarte asemanator cu chilomicronii deprima apoi membrana apicala inspre lumen, formandu-se astfel o "inmugurire" care, in final, pierde contactul cu celula. Exista astfel o mica pierdere de citosol si membrana plasmatica, foarte repede refacuta insa de celula secretoare.
TIPURI MORFOLOGICE DE ADENOMERE
Adenomerul reprezinta componenta secretoare propriu-zisa a epiteliului glandular exocrin si este format din celule epiteliale inalt diferentiate pentru secretie. Din punct de vedere morfologic, dupa modalitatile de organizare a celulelor secretoare, se pot deosebi urmatoarele tipuri de adenomere: tubular, acinos/alveolar, sacciform.
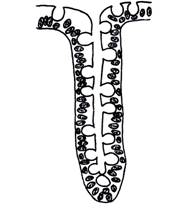
Adenomerul tubular
Adenomerul tubular are structura unui tub, comparabila cu un "deget de manusa". Peretele acestui
adenomer este format din celule de obicei cubice sau
cubico-prismatice, situate pe o membrana bazala. Prin aranjamentul
lor, celulele realizeaza un lumen situat in
centrul adenomerului, care se va continua cu lumenul canalului. Un astfel de adenomer poate avea un traiect aproximativ rectiliniu
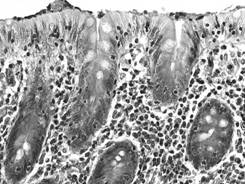
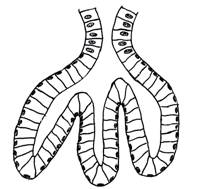
(adenomer tubular drept - de exemplu, glanda intestinala Lieberkühn) (fig.
I.2.5, fig. I.2.6, fig.
I.2.7), un traiect
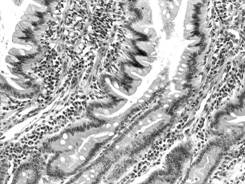
ramificat (adenomer tubular ramificat - glanda pilorica) (fig. I.2.8, fig. I.2.9),
un traiect ondulat,
sinuos (adenomer tubular sinuos -
glanda fundica) sau traiect
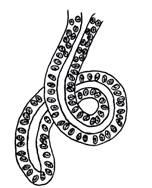
intortocheat, sub forma de ghem (adenomer tubular glomerulat - glanda
sudoripara) (fig. I.2.10, fig. I.2.11).
|
|
|
Fig. I.2.5. Glanda Lieberkühn - schema in MO |
|
|
|
Fig. I.2.6. Glanda
Lieberkühn |
|
|
|
Fig.
I.2.7. Glande Lieberkühn intre |
|
|
|
|
Fig. I.2.8. Glanda tubulo-ramificata - schema in MO |
Fig. I.2.9. Glanda
pilorica (tubulo-ramificata) |
|
|
|
|
Fig. I.2.10. Glanda tubulo-glomerulata - schema in MO |
Fig.
I.2.11. Glanda sudoripara
(tubulo-glomerulata) |
Din punct de vedere citologic, aceste adenomere pot contine fie un singur tip celular, fie doua sau mai multe tipuri celulare.
|
Observatie |
Adenomerul acinos/alveolar
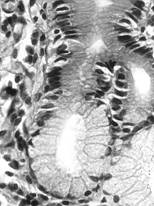
Adenomerul acinos/alveolar este alcatuit din celule care, in ansamblul lor, formeaza o structura sferica (acin) sau alungita elipsoidala (alveola). Celulele, dispuse pe un singur rand aranjat pe o membrana bazala, au forma de trunchi de piramida, cu un pol bazal mai larg si un pol apical ceva mai redus. Ele determina in centrul adenomerului un lumen de dimensiuni diferite care se va continua cu un canal. Datorita acestei structuri, in microscopie optica, pe sectiunile histologice, acinul apare rotund sau alungit.
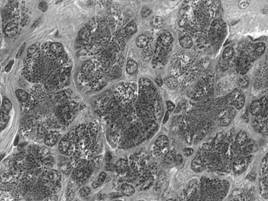
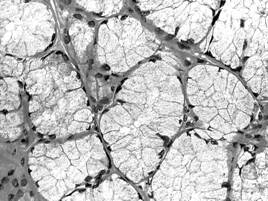
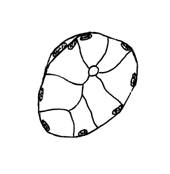
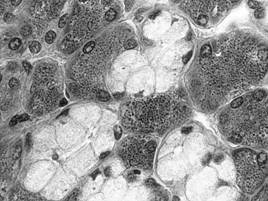
In raport cu natura produsului de secretie, adenomerele acinoase pot fi: seroase, mucoase sau mixte. In acinul seros (fig. I.2.12, fig. I.2.13) celulele secretoare sunt de tip zimogenic, iar lumenul este central, mic si stelat, datorita distantarii polurilor apicale ale celulelor secretoare, deasupra complexelor jonctionale apicale. Acest tip de acin apare in pancreasul exocrin, parotida (glanda salivara seroasa pura) si in glandele salivare mixte.
Acinul mucos (fig. I.2.14, fig. I.2.15) contine in exclusivitate celule de tip mucigen (cu pol inchis), a caror citoplasma apare palida. Lumenul acinului mucos este mai larg comparativ cu acinul seros, si aparent rotund. Acest tip de acin se gaseste in glandele salivare mixte si in glanda palatina (glanda salivara mucoasa pura).
|
|
|
|
Fig. I.2.12. Acini serosi - schema in MO |
Fig. I.2.13. Acini serosi |
|
|
|
|
Fig. I.2.14. Acini mucosi |
Fig. I.2.15. Acin mucos - schema in MO |
|
|
|
|
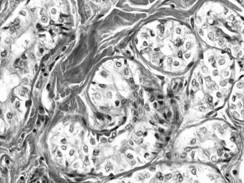
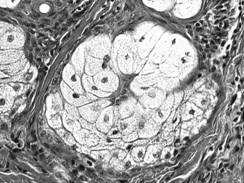
Fig. I.2.16. Acin mixt - schema in MO |
Fig.
I.2.17. Acin mixt |
Acinul mixt (fig. I.2.16, fig. I.2.17) este alcatuit majoritar din celule mucigene (in numar de 4-5) si din cateva celule seroase (in numar de 1-5) ce coafeaza de obicei, sub forma unei semilune - semiluna Gianuzzi, un pol al adenomerului. Acest tip de acin este caracteristic pentru glandele salivare mixte.
|
Observatie |
Adenomerul alveolar reprezinta, in fapt, o varianta morfologica a celui acinos, cu localizare limitata la glanda mamara si la prostata. Din punct de vedere citologic, celulele adenomerului alveolar au o forma cubico-prismatica. Natura produsului de secretie fiind complexa, nu este posibila incadrarea celulelor, ca si tip secretor, in categoria seros sau mucos.
Adenomerul sacciform
Adenomerul sacciform (fig. I.2.18, fig. I.2.19) este specific pentru glanda sebacee situata in derm. Acest adenomer, cu forma de sac, nu are un lumen constituit si este alcatuit - in cea mai mare parte - din celule sintetizante de lipide. La periferie prezinta o membrana bazala, pe care este situat un strat de celule cubico-prismatice cu rol de strat generator - similar stratului bazal din epiteliul stratificat pavimentos. Peste stratul bazal se gasesc suprapuse straturi de celule poliedrice, in stadii progresive de incarcare lipidica si degenerare, astfel incat la portiunea de deschidere a glandei sebacee celulele au o citoplasma de aspect spumos si sunt lipsite de nuclei. In interiorul adenomerului, intre celule, se remarca travee intens colorate, care sugereaza forma unor septuri. Aceste travee reprezinta de fapt coloane de celule keratinizate, provenite prin diferentiere din acelasi strat generator si avand rol de sustinere a celulelor producatoare de sebum.
Nu exista un canal de excretie propriu-zis, adenomerul prezentand o portiune ingustata, (considerata de unii autori gatul glandei), tapetata de un epiteliu stratificat pavimentos in continuitate cu teaca epiteliala externa a foliculului pilos (daca glanda este atasata la folicul) sau cu stratul bazal si spinos al epidermului (daca glanda este independenta de folicul si se deschide direct la suprafata pielii).
|
||||
|
|
||||
|
Fig. I.2.18. Glanda sebacee - schema in MO |
||||
|
|
||||
|
Fig. I.2.19. Glanda sebacee |
||||
TIPURI MORFOLOGICE DE CANALE
Canalele de excretie sunt structuri tubulare epiteliale ce leaga adenomerele de o suprafata epiteliala, permitand eliminarea si scurgerea produsilor de secretie. Canalele sunt delimitate de o membrana bazala pe care se gasesc celule epiteliale cu multiple jonctiuni intercelulare.
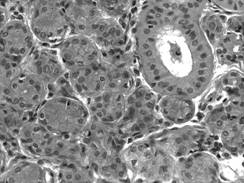
In raport cu diametrul canalelor, epiteliul de tapetare al acestora poate fi: simplu pavimentos, simplu cubic, simplu prismatic (fig. I.2.20), bistratificat cubic, pseudostratificat, stratificat cilindric. In anumite glande, insa, in unele segmente canalele sunt formate din celule transportoare de ioni, cu rol important in desavarsirea si concentrarea produsilor de secretie exocrina (de exemplu, glandele salivare, caile biliare).
Din punct de vedere histoarhitectonic, canalele glandelor exocrine pot fi: (i) simple,
|
|
|
Fig. I.2.20. Canal de excretie - glanda parotida (HE, x 200) |
atunci cand unul sau mai multe adenomere isi varsa continutul intr-un canal unic; (ii) compuse, atunci cand canalele ramificate converg progresiv inspre un canal principal sub forma unei arborizatii, fiecare din multiplele varfuri din ramificatie venind in contact direct cu adenomere.
VARIETATI DE GLANDE EXOCRINE
O clasificare a glandelor exocrine poate porni de la numarul de celule secretoare componente, existand astfel glande unicelulare sau glande multicelulare.
Alte criterii luate in discutie pentru clasificare sunt tipul celular, modalitatea de excretie, localizarea in raport cu epiteliul de tapetare, histoarhitectonica adenomerelor si a canalelor.
Glande unicelulare
Glandele unicelulare reprezinta in fapt celule secretoare dispersate in grosimea unui epiteliu. Exemplul caracteristic este celula caliciforma (celula mucigena cu pol mucos deschis), care apare in grosimea epiteliului respirator si digestiv. Denumirea provine din asemanarea celulei cu calicele unei flori (eng. goblet cell).
In microscopia optica, polul bazal al celulei, fixat pe membrana bazala, contine nucleul (mai mult sau mai putin impins periferic), citoplasma si majoritatea organitelor celulare. Cele doua treimi apicale prezinta o fina patura citoplasmatica periferica, numita teaca (gr. theca) si in interior o multitudine de granule de mucigen. In tehnicile de rutina individualizarea granulelor dispare, datorita tendintei de umflare si coalescenta a acestora. Microscopia electronica (fig. I.2.21) atesta insa originea golgiana a vacuolelor, delimitate de unitati membranare cu continut glicoproteic, eventual sulfatat.
Eliminarea secretiei conduce la diminuarea diametrului transversal al celor doua treimi apicale si revenirea tinctorialitatii, prin expansiunea citoplasmei. Desi secretia de mucus este continua, ea poate fi accelerata prin stimuli parasimpatici sau substante chimice iritative (de exemplu, inhalare de vapori iritanti). Prin pozitia in raport cu epiteliul, celula caliciforma este considerata si o glanda intraepiteliala.
Glande multicelulare
Marea majoritate a glandelor exocrine este constituita din glande multicelulare. In functie de localizarea lor, au fost introduse notiunile de glande intraepiteliale si, respectiv, extraepiteliale.
Faptul ca celulele secretoare se gasesc in insasi grosimea unui epiteliu face ca, intr-o atare situatie, ele sa fie considerate drept glande multicelulare intraepiteliale. Localizarea intraepiteliala este specifica in cazul unor forme simple de glande multicelulare.
|
|
|
Fig.
I.2.21. Celula cu pol mucos deschis -
ultrastructura |
Astfel,
epiteliul simplu cilindric gastric de suprafata si cel al
criptelor gastrice este format dintr-un rand de celule inalte, toate secretand
o substanta mucoida, cu proprietati protectoare
fata de aciditatea sucului gastric. De aceea, se considera
ca epiteliul respectiv are deopotriva rol de tapetare si rol
secretor. Un alt exemplu apare in epiteliul de tip respirator de la nivelul
foselor nazale,
in constitutia caruia exista grupe de cateva celule de tip
caliciform, care formeaza asa-numitele pete secretoare. In
situatia in care structurile de tip adenomer se gasesc in afara
tesutului epitelial de tapetare, conexiunea cu suprafata acestuia
facandu-se printr-un sistem canalicular, glandele sunt incadrate in
categoria glandelor extraepiteliale.
Dupa cum am mentionat anterior, glandele extraepiteliale se
eticheteaza dupa tipul de adenomer, tipul de canal, natura
secretiei si modul de eliminare. Astfel, in definirea si
caracterizarea unei glande exocrine se iau in considerare toate criteriile de
clasificare. Prezentam in continuare cateva exemple de utilizare a nomenclaturii:
glanda parotida: glanda extraepiteliala, multicelulara, tubulo-acinoasa compusa, seroasa (tip celular - celula sintetizanta de proteine/polipeptide), merocrina;
glanda mamara: glanda extraepiteliala, multicelulara, tubulo-alveolara compusa, apocrina;
glanda sebacee: glanda extraepiteliala, multicelulara, sacciforma, tip celular - celula sintetizanta de lipide, holocrina;
glanda intestinala Lieberkühn: glanda extraepiteliala, multicelulara, tubulara simpla dreapta;
glanda
pilorica: glanda extraepiteliala,
multicelulara, tubulo-ramificata, mucoasa
(tip celular - celula sintetizanta
de glicoproteine);
celula caliciforma: glanda intraepiteliala, unicelulara, mucoasa (tip celular - celula sintetizanta de glicoproteine), merocrina.
Copyright © 2025 - Toate drepturile rezervate