|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
ORGANELE LIMFOIDE
Organele limfoide (organa lymphopoietica) produc, stocheaza si "educa" limfocitele, care sunt principalele celulele efectoare ale apararii imune.
Totaliatea organelor limfoide alcatuiesc sistemul imun sau limfoid care cuprinde structurile si celulele din organismul animal care au capacitatea de a distinge selful de nonself si de a distruge sau inactiva substantele sau microorganismele straine ce patrund intr-un organism viu.
Componenta de baza a organelor limfoide este tesutul limfoid Tesutul limfoid este alcatuit din reticul si celule libere (Zarnea-1990). Reticulul este format din fibre reticulare, celule reticulare si macrofage fixe. Celulele libere sunt, in marea lor majoritate, limfocite (aflate in stadii diferite de diferentiere si matuare), plasmotice si macrofage.
Exista 2 tipuri de tesut limofid: - tipul lax, in care predomina celulele reticulare; si - tipul dens, in care predomina limfocitele, care se organizeaza in noduli sau foliculi limfoizi.
Se deosebesc doua tipuri de organe limfoide: - primare (sau centrale) si secundare (sau periferice).
Organele limfoide primare apar primele in filogeneza si in embriogeneza. Sunt reprezentate de: timus si bursa cloacala ( Fabricius) si echivalentii ei la mamifere.
Organele limfoide secundare sunt reprezentate de: ganglioni (noduri) limfatice, splina, placi Peyer, apendice cecal, amigdale, vegetatii adenoide, noduli (foliculi) limfoizi, raspanditi difuzi in diferite mucoase. In filogeneza si in ontogeneza apar mai tarziu, dupa structurarea oragenelor primare. Sunt colonizate de celule diferentiate in organele centrale, celule ce ocupa teritorii specifice, denumite zone timus dependente", populate de limfocite "T" sau zone timus independente", ocupate de limfocitele B".Au in structura lor o retea fina si complicata de vase sangvine si limfatice care creeaza posibilitatea ca limfocitele sa ia contact cu antigenele patrunse in organism sau in circulatie. Celulele sistemului imunitar au posibilitatea sa se recirculeze si sa se reintoarca in teritoriile specifice ( fenomen cunoscut sub denumirea de "HOMING"). In organele limfoide secundare, limfocitele au posibilitatea sa se diferentieze si sa se matureze in continuare, sub influenta stimulilor antigenici. Limfocitele pot sa se transforme in celule efectoare sau in celule cu memorie .
1. Organele limfoide primare sau centrale
Dupa ZARNEA-1990, organele limfoide primare prezinta mai multe caractere generale:
1. apar mai de timpuriu in cursul dezvoltarii embrionare;
2. sunt centre de formare, diferentiere si diseminare in organism a limfocitelor specializate "T" sau "B";
3. in aceste organe, limfopoieza porneste de la celulele STEM multipotente si este independenta de stimularea antigenica. Limfopoieza este intensa inainte de nastere (cand animalul este mai putin expus la antigene), precum si la animalele "germ free";
4. extirparea organelor limfoide primare produce tulburari si deficiente ale raspunsului imun, cu atat mai grave, cu cat se efectueaza mai de timpuriu. Astfel, extirparea inainte de aparitia limfocitelor imunocompetente duce la suprimarea functiilor imunitatii specifice. Extirparea tardiva nu genereaza efecte grave, daca numarul limfocitelor imunocompetente nu este micsorat din alte cauze (iradiere, ser antilimfocitar,varsta inaintata).
5. dupa iradiere, organele limfoide primare pot fi repopulate numai cu celule STEM si nu cu limfocite diferentiate din limfonoduri (ganglioni), timus sau bursa;
6. dupa stimularea cu antigene, organele limfoide primare nu prezinta modificari citologice caracteristice raspunsului imun;
7. celulele produse de organele limfoide primare migreaza in organele limfoide secundare, unde isi desfasoara activitatea la nivelul unor compartimente functionale distincte.
1.1. Timusul
Timusul (timus) este un organ limfoepitelial, ce detine un rol esential, de sef de "orchestra" in realizarea reactiilor de aparare imunitare. La exterior, are o capsula (capsula) de tesut conjunctiv, din care se desprind septuri (septum corticale), care impart parenchimul in mai multi lobuli.
Lobulul timic (lobulus thymi) este unitatea morfofunctionala a timusului. Fiecare lobul are o zona corticala (cortex), periferica, mai inchisa la culoare, si o zona medulara (medulla), centrala, mai deschisa la culoare. Zona corticala apare comuna pentru mai multi lobuli, trecand dintr-un lobul in altul (fig. 1.).
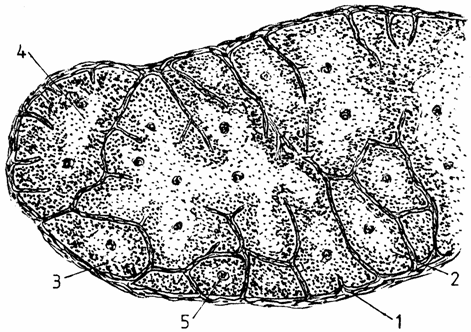
Fig. 1. Histostructura timusului
1 - capsula; 2- lobul; 3 - corticala; 4 - medulara; 5 - corpuscul timic.
Cortexul cuprinde o populatie densa de limfocite T, rare celule epiteliale reticulare dispersate, putine macrofage si limfocite mari. Celulele epiteliale reticulare (epithelioreticulocytus thymi) sunt stelate, cu nuclei ovali, eucromatici. Ele jonctioneaza intre ele prin desmozomi si tonofilamente.
In raport cu structura si localizarea lor, se disting mai multe tipuri de celule epiteliale timice: celule doica, celule epiteliale dendritice din cortexul profund si celule epiteliale din medulara. Ele nu sunt celule limfoide, dar detin un rol cheie in proliferarea si diferenterea limfocitelor timice in curs de dezvoltare.
Celulele doica (thymic nurse cells) sunt celule specializate, localizate in stratul epitelial al cortexului extern. Ele se evidentiaza greu in timusul intact. Se pot izola prin tratament energic cu enzime proteolitice. Apar sub forma unor celule mari, care exprima pe suprafata lor antigene de clasa CMH II si cuprind limfocite corticale incluse in vezicule citoplasmatice. Unii autori considera ca ar fi timocite corticale (fig. 2.).
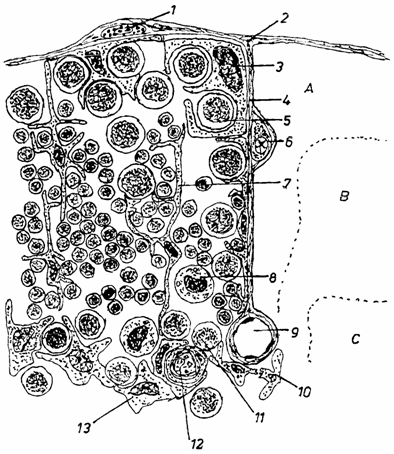
Fig. 2. Schema celulelor din componeneta timusului (dupa Hood-1984)
1 - mastocit extratimic; 2 - capsula; 3 - celula doica; 4 - sept; 5 - limfocit mare din cortex; 6 - plasmocit din sept; 7 - celula epiteliala dendritica corticala; 8 - macrofag; 9 - vas sangvin; 10 - celula dendritica interdigitata; 11 - limfocit mediu din medulara; 12 - corpuscul timic (hassal); 13 - celula epiteliala medulara.
Nu se cunosc inca semnificatia si originea lor, fiind considerate de unii autori ca avand origine epiteliala si rol in limfopoieza timica.
Celulele epiteliale dendritice se gasesc in cortexul profund. Prezinta numeroase prelungiri foarte ramificate (dendritice), care realizeaza o retea si stabilesc interconexiuni complicate in drumul limfocitelor spre regiunea medulara. Ele delimiteaza spatii (lacuri) pline cu limfocite, care se gasesc la limita dintre corticala si medulara. Pe suprafata acestor celule si a prelungirilor lor se gasesc mari cantitati de antigene codificate ale CMH.
Celulele epiteliale din medulara au prelungiri lungi, care formeaza o retea laxa, ce realizeaza un cadru propice in care se asigura maturarea limfocitelor T. Contin pe suprafata lor intreaga gama de antigene, codificata de genele CMH. Unele din aceste celule se grupeaza strans, se dispun concentric si realizeaza corpusculii HASSAL (corpusculum thymicum), relativ sferici, prin hialinizarea citoplasmei si gruparea lor sub forma unor lamele concentrice in jurul unor zone centrale omogene eozinofile.
Rolul corpusculilor HASSAL nu este pe deplin elucidat. Se considera ca pot avea un rol secretor, probabil edocrin, si ca participa la distrugerea limfocitelor nematurate corespunzator, pentru a deveni imunocompetente.
In medulara lobulilor timici se gasesc doua tipuri de celule dendritice nefagocitare: - celulele reticulare denditritice si celule dendritice interdigitate, care contribuie la formarea unei tesaturi laxe. Ele provin din precursori medulari si sunt implicate in prezentarea antigenelor in organele limfoide secundare.
Macrofagele joaca rol de "sentinele", fiind dispuse pe ambele parti ale jonctiunii corticomedulare, in centrul curentului de limfocite.
Celulele limfoide din linia "T" (lymphocytus thymicus) sau timocitele (thymocytus) formeaza populatia celulara dominanta. Ele ocupa reteaua formata de prelungirile ramificate ale celulelor epiteliale. Sunt mult mai numeroase in cortexul timic decat in medulara.
Rolul timusului. La inceputul vietii, timusul este un organ esential pentru maturarea imunologica normala a limfocitelor. La animalele adulte, timusul participa la amorsarea cu elemente a sistemului de aparare imuna de tip celular. Astfel, celulele epiteliale din timus furnizeaza seturile diferentiale ale antigenelor CMH si asigura "educatia" celulelor "T", pentru recunoasterea componentelor "SELF". Alte celule din timus, neidentificate inca, secreta mai multi hormoni peptidici implicati in maturarea limfocitelor T.
1.1.1. Organogeneza timusului
In filogeneza, timusul apare de timpuriu, inca de la pesti. La pasari si mamifere este singurul organ limfatic complet dezvoltat la nastere.
Timusul se dezvolta din epiteliul ventral al celei de a treia (la mamifere) si a patra (la pasari) pungi branhiale, in urma interactiunii dintre ectodermul branhial si endodermul faringian. Astfel, iau nastere doi diverticuli epiteliali ce se evagineaza in mezenchimul adiacent, se izoleaza de formatiunile faringiene, fuzioneaza in plan median , rezultand un conduct, care se orienteaza descendent, ajungand la locul de devoltare a organului ( fig. 3. ).
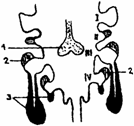
Fig. 3. Organogeneza timusului
1 - Tiroida; 2 - Paratiroidele; 3 - Timusul.
I - IV - Arcuri branhiale.
Prin proliferarea epiteliului, diverticulii si conductul se transforma in muguri plini cu celule epiteliale poliedrice, inconjurati cu tesut mezenchimal. Mezenchimul zonal genereaza capsula, septele interlobare si interlobulare. Totalitatea mugurilor epiteliali formeaza timusul embrionar primitiv epitelial, ce prefigureaza viitorul organ. La soarece, timusul embrionar are origine ecto-endodermica, la cartita este ectodermic, iar la hominide este de natura endodermica. Prin patrunderea de elemente mezenchimale intre celulele epiteliale ale mugurilor timici, rezulta o retea de cordoane epiteliale anastomozate, in ochiurile careia se dispun capilare sangvine si celule mezenchimale. In acest fel, timusul devine un organ cu dubla sau tripla origine embrionara: - endo-mezodermica, la hominide; ecto-mezodermica, la cartita; sau ecto-endo-mezodermica, la soarece. La embrionul de gaina, timusul apare in zilele 7 - 8 de incubatie, iar la soarece in ziua a 11-a de dezvoltare intrauterina, finalizarea organogenezei avand loc postnatal.
Ulterior, in parenchimul timusului se diferentiaza doua zone:- una superficiala, corticala, si alta profunda, medulara. In zona medulara, vasele si nervii patrund mai precoce si sunt mai abundente. In ontogeneza, proportia celor doua zone se modifica o data cu varsta. Astfel, la hominide si alte specii de animale, corticala apare de doua- trei ori mai dezvoltata decat medulara, in momentul parturitiei, pentru ca la pubertate cele doua zone sa aiba o dezvoltare relativ egala. Dupa Botarel si colab. (1980-1981), la foetusii de suine de 60 zile este mai dezvoltata zona corticala; la 90 zile zona medulara se extinde, intensificandu-se morfogeneza si acivitatea corpusculilor Hassal; iar la 113 zile prenatal si la purceii de o saptamana timusul este complet dezvoltat.
De timpuriu, timusul este populat cu celule stem pluripotente, care initial provin din sacul vitelin, apoi din ficat si din maduva osoasa. Stutmann (1978, 1985) considera ficatul foetal ca o statie obligatorie pentru celulele care provin din sacul vitelin. Aceste celule patrund in mezenchim si intre celulele epiteliului timic, alaturi de limfocite. Din celulele epiteliale se dezvolta citoreticulul timic. Corpusculii Hassal apar destul de timpuriu, fiind observati la foetusii suini de 60 zile.
Timusul poate prezenta, ca tulburari de dezvoltare, ectopii, agenezii si hipogenezii. Prin persistenta canalelor timofaringiene, apar traiecte fistuloase, cu lobi timici accesorii.
1.2. Bursa cloacalA (fabricius)
Bursa cloacala sau diverticulul cloacal (diverticulum cloacae) este un organ limfoepitelial specific pasarilor, cu aspect de punga, situat dorsal de proctodeum, cu care comunica printr-un canal bursal. Acest canal poate fi flancat de doua formatiuni bursale accesorii.
Peretele bursei este format din trei tunici: seroasa, musculoasa si mucoasa.
Seroasa este reprezentata de peritoneul local, care acopera diverticulul cloacal pe extremitatea sa cranio- dorsala.
Musculoasa este formata din fibre musculare netede, dispuse in doua straturi, longitudinal extern si circular intern. Mucoasa ocupa cea mai mare parte din grosimea peretelui bursal, avand lamina propria ocupata, apropape in totalitate, de foliculi (noduli) bursali.
Mucoasa apare puternic cutata, formand pliuri groase, al caror centru este ocupat de foliculi, cu contur poliedric, iar suprafata pliurilor este acoperita de un epiteliu simplu sau pseudoprismatic (Bacha-1990, Banks-1992) columnar. In acest epiteliu exista trei tipuri de celule columnare (I, II, III), in functie de granulele pe care le contin (fig. 4.).
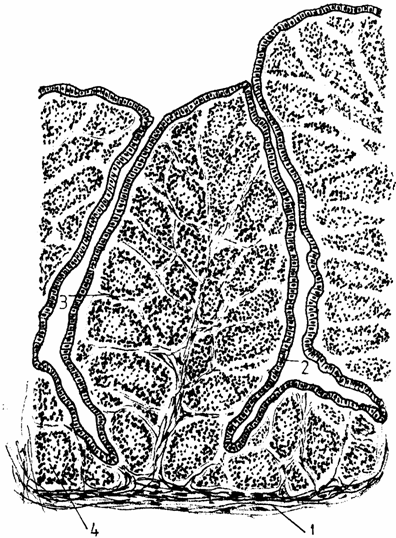
Fig. 4. Histostructura bursei cloacale
1 - peretele bursei; 2 - epiteliul simplu prismatic; 3 - corion; 4 - foliculi (noduli) bursali.
Cararacterul de epiteliu simplu columnar este mai evident atunci cand acopera proeminentele deteminate de proliferarea fiecarui folicul. Acest epiteliu prolifereaza in profunzime, celulele epiteliale amestecandu-se cu celule din foliculi. Foliculii bursali pot avea doua aspecte: un aspect uniform, la foliculii primari, si cu doua zone, corticala si medulara, la foliculii secundari.
Foliculii (nodulii) bursali secundari apar dupa stimulari antigenice. Ei prezinta la periferia medularei un strat de celule epiteliale nediferentiate, cubice, si un strat de capilare. Aceste celule epiteliale cubice sunt implicate in activarea limfocitelor ce trec din corticala in medulara pentru a sintetiza anticorpi.
Corticala este separata de medulara printr-o membrana bazala. Zona corticala apare mai inchisa la culoare, cuprinzand numeroase limfocite, limfoblaste, celule in mitoza si numeroase macrofage. Vascularizatia zonei corticale este asigurata de arteriole, venule si capilare.
Zona medulara apare mai deschisa la culoare si cuprinde celule nediferentiate, limfoblaste si limfocite. Vascularizatia zonei medulare este asigurata de o retea de capilare mari, dispusa imediat sub membrana bazala.
Rolul bursei consta, in principal, in maturarea limfocitelor B, care vor secreta anticorpi, asigurand imunitatea de tip umoral. Initial, bursa are o structura pur epiteliala, fiind colonizata mai tarziu cu celule precursoare, venite din sacul vitelin sau din ficatul embrionar. Desi involueaza dupa ecloziune, isi mentine functiile prin care sunt maturate celulele B imature, aflate in circulatie. Eliminarea experimentala sau patologica a bursei determina aparitia unui deficit in functionarea limfocitelor B, cu atat mai sever, cu cat a trecut mai mult timp de la anihilarea bursei.
Pentru maturarea limfocitelor B, acestea trebuie sa treaca prin epiteliul bursal, care joaca rolul unui factor microambiental. De asemenea, bursa produce un factor hormonal, bursopoietina, care, in vitro, induce diferentierea selectiva a limfocitelor B. Hood (1984) nu exclude posibilitatea ca si alte tesuturi hemopoietice sa participe, la pasari, la diferentirea limfocitelor B.
Limfocitele B parasesc bursa prin vasele sangvine care aprovizioneaza foliculii bursali, ajungand in tesuturile limfoide.
1.2.1. Organogeneza bursei cloacale (Fabricius)
Originea embrionara a bursei cloacale (Fabricius) suscita inca discutii. Primele cercetari considerau ca bursa are origine ectodermica, dezvoltandu-se din peretele superior al proctodeumului. Alti cercetatori sustin originea endodermica, aratand ca dezvoltarea bursei cloacale se datoreste unor celule epiteliale, cu aceeasi origine ca cele din intestinul posterior. Jolly si Boyden, citati de Giurgea - 1982, considera ca bursa este rezultatul unei proliferari a epiteliului endodermal din zona marginala a cloacei. Majoritatea cercetatorilor sustin originea endodermica a bursei, considerand-o ca o evaginare a endodermului pe fata dorsala a extremitatii caudale a intestinului posterior, in vecinatatea membranei uroproctodeale, ce se interpune intre ectodermul proctodeal si endoderm.
Primordiul bursal apare in zilele 4 - 8 de incubare, existand variatii legate de rasa si conditiile de incubare. Are forma unei proliferari, asezata pe fata dorsala a endodermului cloacal. In acest fel, apare o formatiune diverticulara cu suprafata interna neteda, pana in ziua a noua de incubare (Gaboreanu si Magherusan - 1970). Epiteliul endodermal ce captuseste lumenul bursei este format din 2 - 4 straturi de celule poliedrice, cu citoplasma intens bazofila, ceea ce indica o intensa sinteza de proteine. Celulele epiteliale sunt asezate pe o membrana bazala fina, care le desparte de mezenchimul subiacent bogat in celule pironinofile ce produc intens fibre de reticulina, fine si abundent ramificate. Fibrele de reticulina formeaza un strat fibrilar dens, sub membrana bazala. Dupa 7 - 9 zile de incubare, mezenchimul periepitelial si epiteliul bucal prolifereaza spre interior, incat mucoasa bursei sufera un proces de cutare, aparand 5 - 10 pliuri, care capata un aspect lamelar. Fiecare lama prezinta un ax mezenchimatos, bogat vascularizat, acoperit de un epiteliu, in care celulele embrionare incep sa se dispuna intr-un singur strat. In acest mod, din ziua a 9-a de incubare, bursa are o structura asemanatoare intestinului, avand peretele mult mai subtire, datorita reducerii musculoasei.
Din ziua a 10-a, in mezenchimul bursei apare un numar mare de celule acidofile, dispuse in jurul capilarelor si venelor, celule considerate de Ackermann si Knouf (1964) a fi granulocite. In ziua a 11-a de incubare, in epiteliul bursal apar grupe mici de celule mobile, cu origine limfoida, grupe care reprezinta fazele initiale de formare a foliculilor. Astfel, mugurii viitorilor foliculi bursali apar in grosimea epiteliului, iar in corion se declanseaza procesul de granulocitopoieza (Giurgea-1982).
Mugurii epiteliali cresc rapid datorita mitozelor frecvente, astfel incat vor deplasa membrana bazala, infundand-o in corionul adiacent. Celulele epiteliale isi modifica aspectul initial si se diferentiaza in celule limfoide, sub inductia mezenchimului, cu care stabilesc raporturi morfofunctionale stranse.
Mugurii epiteliali cuprind un rand de celule nediferentiate, numite celule de sustinere, cu rol de reinnoire a epiteliului, dispuse pe membrana bazala. Precursorii celulelor limfatice sunt celule stem pluripotente, ce provin din sacul vitelin, ficatul embrionar si maduva hematogena. Ele ajung pe cale sanguina in primordiul bursal, unde gasesc un mediu propice pentru proliferarea si maturarea lor.
In apropierea ecloziunii, pliurile mucoasei bursale sunt bine dezvoltate, acoperite de un epiteliu, cu foliculi tineri in corion, lipsiti de zona corticala.
Momentul aparitiei zonei corticale nu este bine precizat, existand cercetatori dupa care aparitia corticalei are loc preeclozional (in ziua a 17-a), in timp ce dupa altii, corticala apare posteclozional (din ziua a 5-a). Initial, corticala apare sub forma unui capison la un pol al foliculului, pentru ca mai apoi sa inconjoare zona medulara. Celulele zonei corticale sunt slab diferentiate, foarte asemanatoare cu cele ce alcatuiesc epiteliul bursal. La periferia corticalei se gasesc 2 - 3 randuri concentrice de celule mai tinere, ce ajung in contact cu fibrele perifoliculare.
Dupa ecloziune, foliculii bursali prezinta o zona corticala, cu limfocite in diviziune rapida, impachetate dens, si o zona medulara, populata de limfocite dispersate si o retea de celule epiteliale.
Prin proliferarea rapida a limfocitelor apar cateva mii de foliculi limfoizi. In fiecare folicul limfoid sunt cuprinse circa 1 000 de limfocite B, care sunt heterogene sub raportul capacitatii lor de a lega antigenele. Fiecare folicul contine limfocite B provenite dintr-o singura celula stem sau numai din cateva. Este posibil ca o celula stem sa dea nastere mai multor clone diferite de limfocite B (Zola-1985). Limfocitele B parasec bursa intrand in vasele sangvine care iriga foliculii bursali si migreaza in formatiunile limfoide secundare, incepand cu ziua a 16-a de incubare.
Bursa cloacala creste rapid in primele 3-4 saptamani dupa ecloziune, atinge dimensiuni maxime la 5 - 12 saptamani si involueza in perioada maturitatii sexuale, datorita influentelor hormonale.
2. Organele limfoide secundare
Organele limfoide secundare sau periferice sunt reprezentate fie de: - organe anatomic distincte, cu structura complexa, precum: ganglionii (limfonoduri) limfatici, splina, amigdalele, placile Peyer; - structuri limfoide difuze, cum sunt structurile limfoide asociate mucoaselor gastro - intestinale (galt) sau respiratorii (balt).
Dupa Zarnea - 1990, organele limfoide secundare prezinta mai multe caractere generale:
1. Organele limfoide secundare au o aparitie si o evolutie limfoida tardiva filo- si ontogenetic, fiind populate de celule limfoide deja diferentiate, provenite din organele limfoide primare. Proliferarea si diferentierea celulelor limfoide in aceste organe contribuie la realizarea raspunsului imun.
2. Organele limfoide secundare contin compartimente specializate pentru celulele T (timodependente) si pentru celulele B
(timoindependente), fara granite rigide, permitandu-se trecerea de la un compartiment la altul.
3. In organele limfoide secundare, limfopoieza este slaba sau inexistenta inaite de nastere. De asemenea, limfopoieza este foarte redusa la animalele anexice (germ free) chiar dupa introducerea parentala a antigenelor.
4. In organele limfoide secundare, limfocitele pot intra din circuitul sangvin, facand capabila repopularea lor dupa iradiere, in absenta celulelor stem;
5. Este imposibila extirparea in totalitate o organelor limfoide secundare.
6. Datorita structurii lor, organele limfoide secundare permit asigurarea unor posibilitati marite de interactiune a limfocitelor cu celulele care prezinta antigenele, cu circulatia sagvina si limfatica, interactiune absolut necesara pentru declansarea si evolutia raspunsului imun.
7. Organele limfoide secundare contin un numar mare de limfocite periferice, reprezentate de celulele cu memorie, produse sub influenta antigenelor din mediu. Animalele "germ free" (anexice) au tesuturile limfatice secundare atipice, in timp ce organele limfoide primare au mecanisme si structuri normale.
Producerea celor 2 clase principale de limfocite T si B, la nivelul organelor limfoide primare, este urmata de localizarea lor la nivelul organelor secundare, unde limfocitele mor daca nu intalnesc antigenul corespunzator. Sub influenta stimulilor antigenici, limfocitele prolifereaza si isi completeaza diferentierea, devenind celule efectoare sau celule cu memorie.
2.1. Nodulii limfoizi
Nodulii limfoizi (nodulus lymphaticus) sau limphonodulii (lymphonodulus) sunt formatiuni microscopice neregulate, sferice (cu diametrul variabil, de 0,2 - 1 mm). Ei sunt formati din tesut limfoid dens. Sunt prezenti in tesutul conjunctiv al diferitelor organe, in lamina proprie (corionul) a tubului digestiv, a cailor respiratorii, a aparatului urinar etc. Limphonodulii nu sunt structuri permanente, putand sa apara si sa dispara in functie de stimularile antigenice.
Astfel, foliculii limfoizi sunt mai rari la nou-nascuti si la animalele crescute in mediu liber de germeni, prezenta lor fiind determinata de stimulii antigenici locali. Cand prezinta centrii germinativi, se numesc "foliculi (noduli) secundari", fiind diferiti de cei primari, care se remarca prin uniformitatea aspectului lor.
2.2. Limfonodurile sau ganglionii limfatici
Limfonodurile sau ganglionii limfatci sunt organe limfoide secundare, cu aspect reniform sau sferoidal. Se gasesc izolate sau grupate de-a lungul vaselor limfatice. Majoritatea speciilor de pasari nu au noduri limfatice, cu exceptia pasarilor acvatice, unde prezinta o forma primitiva de organizare.
Histostructura limfonodurilor cuprinde: - capsula (capsula), ce detaseaza trabecule (trabecula) conjunctive care divid incomplet parenchimul ganglionar; si - parenchimul ganglionar, care prezinta 3 zone distincte: - corticala (cortex); - paracorticala (paracortex) sau cortexul difuz; si - medulara (medulla).
Capsula si parenchimul ganglionar sunt separate printr-un sinus periferic marginal - sinusul subscapsular (sinus subcapsularis), de la care pornesc sinusurile corticale perinodulare (sinus corticalis perinodularis), care patrund in parenchimul cortical si se continua cu sinusurile medulare (sinus medullaris), separate unele de altele prin cordoane medulare (chorda medullaris). Sinusurile medulare dreneaza limfa in vasele limfatice eferente (vas limphaticum efferens).
Sinusul subscapular nu apare ca o cavitate cu lumen deschis, ci ca o retea densa prin care se filtreaza limfa (fig. 5.).
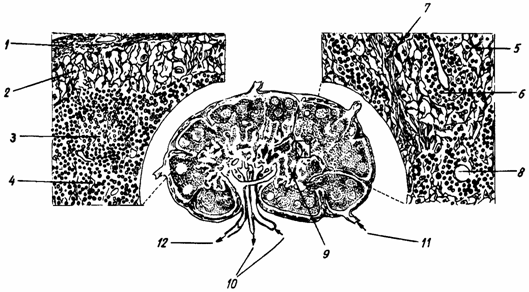
Fig.5. Diagrama structurii unui limfonod (dupa Liebich-1990)
1 - capsula; 2 - Sinus subcapsular; 3 - nodul secundar; 4 - paracortex; 5 - celule reticulare; 6 - limfocite; 7 - trabecule; 8 - vas sangvin; 9 - sinus medular; 10 - artera si vena; 11 - vas limfatic eferent; 12 - vas limfatic eferent.
Parenchimul limfonodului este o retea de celule si fibre reticulare ce umple tot spatiul dintre trabecule si capsula, in ochiurile careia se gasesc limfocite, plasmocite, macrofage, sinusuri limfatice, vase de sange.
Cortexul contine: tesutul reticular, limfocite grupate in formatiuni sferice sau ovale, foliculi sau noduli limfoizi primari (nodulus primarius) (cu un diametru de aprox. 1 mm), ce contin limfocite mici. In cursul raspunsului imun se vor transforma in foliculi sau noduli secundari (nodulus secundarius), caracterizati prin prezenta unei zone de proliferare celulara sau centru germinativ (centrum germinativum), zona timus independenta ,de reactie si de producere a anticorpilor.
Centrii germinativi au o existenta tranzitorie. Ei apar in urma actiunii unui stimul antigenic si marcheaza transformarea nodulilor primari in noduli secundari. Ei sunt formati, in principal, din celule stromale (reticulare) dendritice, care prezinta numeroase plieri ale plasmalemei si permit acumularea unei mari cantitati de antigene.
Parenchimul centrilor germinativi cuprinde 2 zone: - o zona dens populata si alta cu celule rare.
In zona dens populata predomina celulele bogate in ribozomi, pironinofile, care produc si secreta imunoglobuline. Tot in aceasta zona se mai gaseste un numar de macrofage (care fagociteaza celulele moarte si se coloreaza intens, devenind corpi colorabili.
In zona slab populata, celulele sunt rare, dispersate printre celulele reticulare care sunt mai abundente, cu suprafete mari, datorita plierii membranelor. Celulele pot migra dintr-o zona in alta (din compartimentul dens in cel rar populat si, apoi, in masa de celule limfoide inconjuratoare. Zona celulelor dispersate, mai clara, este indreptata spre sursa de stimulare antigenica.
Foliculii secundari apar situati imediat sub capsula. Foliculii secundari, cu centrii germinativi situati mai profund decat acestia, sunt denumiti foliculi tertiari.
Centrii germinativi apar si se evidentiaza in functie de intensitatea stimulului antigenic. Cand dispare stimulul, activitate proliferativa scade, iar foliculul revine la aspectul de folicul primar, care contine macrofage cu incluzii.
Cortexul difuz (profund) sau regiunea paracorticala (paracortex s. zona thymodependens) apare imprecis delimitata, situata intre corticala si medulara. Cuprinde mase neregulate de tesut limfoid dens.
Este o regiune timus-dependenta, populata de limfocite T, in care limfocitele dispar dupa timectomie sau se hipertrofiaza dupa imunizare cu antigene care induc o hipersensibilitate de tip intarziat.
Zona medulara contine tesut limfoid dens, distribuit sub forma de cordoane medulare intre care se afla sinusurile medulare. Cuprinde o populatie celulara mixta, predominant timus-independenta datorita numarului mare de celule B, asociate cu plasmocitele, macrofagele si celulele (limfocite T) pregatite pentu migrare.
Vascularizatia limfonodurilor. In limfonoduri, sangele circula in directie opusa limfei, patrunzand in ganglioni prin hil (pe unde patrund si 90% din limfocite, care vor parasi ganglionul). Limfa patrunde in ganglion prin vase limfatice aferente (vas limphaticum afferens), care strabat capsula si se deschid in sinusul subcapsular. Limfocitele T si B ies din venulele postcapilare trecand prin endoteliu lor (si nu printre celulele endoteliale, cum fac granulocitele si macrofagele) legandu-se in prealabil, cu ajutorul unor glucide de pe suprafata limfocitelor, pe receptorii specifici ai endoteliului.
Dupa extravazare, limfocitele migreaza spre teritoriile specifice, in foliculii primari migreaza limfocitele B, iar in cortexul difuz, limfocitele T. Dupa ce au strabatut teritoriile specifice, limfocitele T si B intra in medulara si ajung, in final, in vasele eferente ce conduc limfa modificata in urma tranzitului prin limfonod. Circulatia prin limfonod permite unui numar mare de limfocite sa treaca prin teritoriile specifice, unde vor intalni substante imunogene, crescand sansa ca un numar de limfocite imunocompetente sa intalneasca antigenul specific si sa reactioneze cu el.
In acest fel, nodurile limfatice apar ca niste structuri dinamice, fiind sediul unui flux continuu de celule, de multiplicare considerabila a limfocitelor si initiere a raspunsului imun.
2.2.1. Formarea nodurilor (ganglionilor) limfatici
Aparitia nodurilor limfatice sau a ganlionilor limfatici este precedata de un proces de multiplicare a vaselor limfatice dintr-un anumit loc din organism, vase care se anastomozeaza, formand un plex. In ochiurile acestui plex apar aglomerari nodulare de celule, ce se diferentiaza din celulele mezenchimale locale. Nodulii cresc, impingand la periferie vasele limfatice, care vor forma sinusul subcapsular. Initial nodulii sunt formati din celule mezenchimale cu mare potential de diferentiere, pentru ca mai apoi, sa fie populati cu limfoblasti si limfocite.
In prima faza de organizare, nodulul limfatic este uniform, dar mai tarziu, apar cordoanele medulare, insotite de sinusuri, diferentiindu-se medulara de corticala.
La hominide, nodurile limfatice apar la sfarsitul lunii a treia de viata intrauterina. La diferite specii de animale, nodurile limfatice apar in cea de a doua jumatate a vietii intrauterine. Morfogeneza nu este incheiata la fatare (Botarel-1981), ci se continua in prima luna de viata extrauterina. Ritmul de organizare difera de la un ganglion la altul, fiind influentat de dezvoltarea fiziologica a zonelor de colectare a limfei. Parenchimul ganglionar se modifica cu varsta, fibrele de colagen si celulele adipoase inlocuind treptat tesutul limfoid.
Ca organe limfoide secundare, nodurile limfatice sufera modificari evidente in cursul imunizarilor si infectiilor, sub raport volumetric si al proportiilor dintre corticala si medulara. De asemenea, se petrec schimbari calitative in populatia celulelor componente (Zarnea -1990).
In cursul raspunsului imun mediat celular, primele modificari se observa dupa 24 de ore, prin aparitia in cortexul periferic a unor celule blastice, care prolifereaza pana in ziua a patra, determinand mariri importante de volum. Celulele endoteliale din venulele postcapilare devin turgescente, iar numeroase celule traverseaza peretele capilar. Celulele blastice dispar in ziua a saptea, dupa ce au format o populatie noua de limfocite mici. Zona corticala si nodulii limfoizi sunt putin modificati. Aceste modificari nu se observa la soarecii timectomizati.
In cursul raspunsului imun mediat umoral, apar modificari semnificative in zonle ocupate de limfocitele B, in regiunea cortexului superficial, timus independenta. Antigenul este atras spre centrul foliculului, unde induce proliferarea limfocitelor, aparitia centrului germinativ si transformarea limfocitelor in plasmocite. Stimularea antigenica atrage macrofagele si alte celule fagocitare, producandu-se si acumulare de plasma. De asemenea, sunt retinute un numar important de limfocite circulante, care se divid intens si care, impreuna cu celulele nelimfoide si cu edemul local, determina o evidenta crestere in volum a nodurilor limfatice.
2.3. Splina
Este cea mai mare acumulare de tesut limfoid din organism. Actioneaza ca un filtru masiv, interpus pe vasele sangvine, avand rol esential in imunitate si in retinerea antigenelor straine, a particulelor straine, a resturilor celulare. Este un sediu al raspunsului imun.
Structura histologica a splinei cuprinde: capsula si parenchimul splenic.
Capsula (capsula) sau tunica fibroasa (tunica fibrosa) este formata din tesut conjunctiv dens neorientat, cu fibre de colagen (fibra collagenosa), fibre elastice (fibra elastica) si celule musculare (myocytus non striatus). Capsula trimite trabecule ce contin artere trabeculare (arteria trabecularis) si delimiteaza compartimente intercomunicante sau loji splenice, in care se gaseste parenchimul alcatuit din pulpa rosie si pulpa alba (fig. 6.).
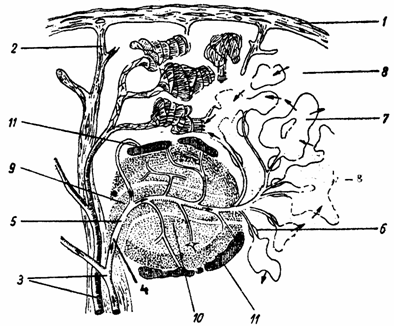
Fig. 6. Structura splinei - schema (dupa Liebich-1990)
1 - capsula; 2 - trabecula; 3 - artera si vena trabeculara; 4 - artera pulpei albe; 5 - artera limfonodulului; 6 - artere penicilate; 7 - capilare sinusoide; 8 - pulpa rosie-cordoane splenice; 9 - teci limfoide periarteriale; 10 - limfonodul secundar; 11 - zona marginala.
Parenchimul splenic sau pulpa splinei (pulpa splenica) este de doua feluri: pulpa rosie (pulpa rubra) si pulpa alba (pulpa alba).
Pulpa rosie este formata din cordoane splenice (chorda splenica) si sinusuri venoase (sinus venularis). Ocupa cea mai mare parte din parenchim.
Cordoanele splenice au o structura de baza (fibre de reticulina, celule reticulare, macrofage) in ochiurile careia se gasesc toate tipurile de elemente albe si un numar foarte mare de hematii.
Sinusurile venoase au diametrul neregulat si sunt situate intre cordoanele splenice. Nomina histologica - 1992 considera ca sinusurile venulare sunt venule postcapilare care leaga vasele capilare terminale de venele pulpei rosii. Sunt captusite de celule endoteliale, ce fagociteaza intens antigenele sub forma de particule. In ele se acumuleaza cea mai mare parte din sangele care patrunde in splina si contine numeroase celule fagocitare si producatoare de anticorpi.
Intre pulpa rosie si pulpa alba se interpune o zona marginala, care delimiteaza pulpa rosie de pulpa alba. Este o zona bogat irigata de numeroase arteriole terminale. In aceasta zona sunt retinute cele mai multe antigene provenite din sange. Contine atat limfocite T, cat si limfocite B, fiind mai bogata in limfocite si plasmocite decat pulpa rosie.
Pulpa alba este formata din limfocite si celule inrudite, dispuse organizat intr-o retea de celule si fibre reticulare, formand teci limfoide periarteriolare (vagina periarterialis lymphatica) si foliculi (noduli) limfatici (lymphonodulus splenicus).
Tecile limfoide periarteriolare sunt mansoane de limfocite organizate in jurul arteriolelor care se detaseaza din arterele trabeculare si patrund in lojile splenice, dand nastere la arterele pulpei albe (arteria pulpae albae). Fiecare teaca are o zona interna, bogata in limfocite T si o zona externa, bogata in limfocite B. Dupa LAMB - 1979, tecile sunt formate exclusiv din celule T, in timp ce foliculii (nodulii) sunt, in cea mai mare parte, formati din celule B.
Foliculii splenici primari au forma sferica sau ovalara, asemanandu-se cu cei din ganglionii limfatici. Sunt raspanditi din loc in loc sau imediat in afara tecilor limfoide periarteriorale, fiind situati frecvent la nivelul bifurcatiilor arterei centrale (arteria lymphonoduli) sau al ramificatiilor ei principale. Ei se transforma in foliculi germinali cu centrii germinativi, in timpul raspunsului imun ;
Splina nu are circulatie limfatica, limfocitele intrand si iesind pe calea circulatiei sangvine (in cca. 6 ore, traverseaza campurile limfoide ale splinei). Artera spelnica se ramifica dupa ce patrunde in oragn in mai multe artere trabeculare (arteria trabecularis), din care se desprind arterele lojilor splenice ce emit arterele pulpei albe (arteria pulpae albae), ce se ramifica in arterele centrale sau ale limfonodulilor (arteria lymphonoduli). Arterele limfonodulilor se ramifica dand arteriole penicilate (arteriola penicilaris). Din arterele penicilate, sangele trece in capilarele poroase, situate la granita dintre tecile limfoide periarteriolare si zona marginala.
2.3.1. Organogeneza splinei
Splina are origine mezodermica. Apare in mezenterul embrionar dorsal, care leaga tubul digestiv de peretele dorsal al embrionului. In jurul unor ramificatii venoase care constiutie primordiul venei porte (vena subintestinala la embrion), mezenchimul adiacent se hiperplaziaza, formand un nodul de tesut mezenchimal mai dens, numit mugure splenic. Celulele mezenchimale se divid rapid, rezultand tesutul reticulat, endoteliul vaselor si parenchimul primitiv. O parte din celulele mezenchimale vor delimita viitoarele sinusuri, iar altele vor participa la formarea unei retele, in ochiurule careia se vor grupa celule de tip sangvin, provenite din celule mezenchimale locale.
Celulele prezente in ochiurile retelei splenice sunt diferite in functie de varsta embrionului sau fatului. Astfel, la hominide, pana in luna a saptea se intalnesc megacariocite, globule albe tinere si globule rosii identice cu cele din maduva hematogena, splina comportandu-se ca un organ mieloid. Din luna a saptea, splina devine organ limfoid, iar la parturitie, in parenchimul splenic se mai gasesc hematii nucleate.
Circulatia arteriala se instaleaza relativ tardiv, sangele arterial fiind drenat in formatiuni venoase dupa ce a strabatut ochiurile retelei. Pulpa rosie si pulpa alba se diferentiaza dupa stabilirea circulatiei arteriale. Dezvoltarea pulpei albe este dependenta de timus.
Splina poate fi afectata de tulburari de dezvoltare, precum: agenezia, fisurarea incompleta sau completa, cu aparitia de spline supranumerare.
2.4. Structurile limfoide ale mucoaselor
Mucoasele contin un numar mare de celule care participa la inducerea si evolutia raspunsului imun, prcum: macrofagele, limfocitele, plasmocitele. Aceste celule pot fi organizate in structuri distincte sau pot fi raspandite difuz in mucoase. Mai bine studiate sunt structurile limfoide
din mucoasele cailor digestive si ale cailor respiratorii, desi in toate mucoasele exista formatiuni limfoide.
Structurile limfoide asociate cailor digestive sunt reprezentate de: amigdale, placi Peyer,diverticulul vitelin, amigdale (placi) cecale si din colon. Formatiunile limfoide asociate aparatului respirator formeaza sistemul limfatic BALT (bronchus associated lymphoid tissue).
Structurile limfoide din intestin formeaza sistemul limfatic GALT (gut associated lymphoid tissue). Aceste structuri nu prezinta capsula si nici vase limfatice aferente. Ele sunt acoperite de un epiteliu diferit de restul epiteliului intestinal, prezentand o «porozitate» destul de mare, care favorizeaza patrunderea antigenelor, incat lichidul care trece spre foliculi prin acest epiteliu inlocuieste limfa aferenta. Materialul antigenic provenit din lumenul organului este, astfel, preluat de macrofage si de foliculii limfoizi.
Placile Peyer sunt prezente la toate mamiferele, intre duoden si illeon, aparand mai dezvoltate in jumatatea distala a intestinului. Numarul lor este diferit de la o specie la alta, existand 12 la soarece, 20 la sobolan si circa 200 la hominide. Ele sunt cantonate in corion si in submucoasa intestinului subtire. Se formeaza prin agregarea mai multor foliculi limfoizi, dar pot aparea si foliculi limfatici izolati. Epiteliul situat deasupra foliculilor formeaza regiunea domului, bogat infiltrata cu limfocite. Epiteliul domului este simplu cubic prismatic si contine o serie de celule modificate, denumite, la soarece, celule asociate cu foliculii (follicle associated cells) sau celule "M" (microfolds cells), la hominide. Celulele M prezinta microvilozitati scurte si neregulate. In citoplasma polului apical se gasesc numerosi tubuli, vezicule si vacuole. Au rolul de a transporta virusurile, microorganismele si particulele straine spre stratul subepitelial
Stratul subepitelial contine numeroase macrofage, celule reticulare dendritice, limfocite T si B, ce pot reactiona cu antigenii. Foliculii (nodulii) limfatici si centrii germinativi contin in special celule B, in timp ce cordoanele interfoliculare contin celule T si venule post-capilare cu endoteliu inalt, ce reprezinta calea de intrare a limfocitelor T si B circulante .
Apendicele cecal, prezent la leporide si hominide, prezinta structuri limfoide cu organizare asemanatoare cu a placilor Peyer. Au domul bine organizat, cu epiteliu foarte poros, iar agregatele de celule B se organizeaza in foliculi (noduli). O zona cu limfocite T este plasata ca un "sandwich" intre 2 foliculi adiacenti.
Amigdalele sunt primele organe limfo-epiteliale care vin in contact cu materialele ingerate la nivelul cavitatii bucale. Au suprafata acoperita cu un epiteliu pluristratificat, infiltrat cu limfocite (fig. 7.).
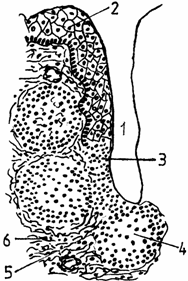
Fig. 7. Amigdala bucala - schema
1 - cripta amigdaliana; 2 - epiteliu stratificat necheratinizat; 3 - infiltratie limfocitara a epiteliului; 4 - noduli amigdalieni; 5 - Infiltratie difuza internodulara; 6 - condensare conjunctiva periamigdaliana
La pasari, diverticulul vitelin (Meckel) devine posteclozional un organ limfo-plasmocito-formator. In corion sunt prezenti noduli limfatici, cu tendinta de a se organiza pe 2 zone, devenind noduli (foliculi) secundari. Este sursa imunogammaglobulinelor imediat in perioada posteclozionala.
Copyright © 2026 - Toate drepturile rezervate