|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Sunt reprezentate prin eritrocite - care la mamifere si om sunt anucleate si poarta numele de hematii - leucocite si placute sangvine sau trombocite.
Din volumul de sange, cea mai mare parte revine eritrocitelor, 44 %, iar celelalte elemente figurate insumeaza abia 1 %.
Sunt elemente celulare anucleate, avand forma de discuri bi- sau uniconcave (fig. nr. 4.6.) care ofera o suprafata maxima de schimb.
 Eritrocitul are o mare plasticitate, putand sa ia
forme variate, care le permite sa treaca prin porii de 2,3-3,0 m fara sa fie distrus. In mediul
hiperton se retracta, devenind plat, iar in cel hipoton se umfla
capatand aspect aproape sferic, crescand pana atinge valorile
critice ale hemolizei, cand in membrana sa apar pori prin care iese
hemoglobina din celula.
Eritrocitul are o mare plasticitate, putand sa ia
forme variate, care le permite sa treaca prin porii de 2,3-3,0 m fara sa fie distrus. In mediul
hiperton se retracta, devenind plat, iar in cel hipoton se umfla
capatand aspect aproape sferic, crescand pana atinge valorile
critice ale hemolizei, cand in membrana sa apar pori prin care iese
hemoglobina din celula.
 Au
diametrul de 7,5-7,8 mm care scade
odata cu inaintarea in varsta a celulei, insumand o
suprafata totala de peste 2.500 m2, destinata
proceselor de schimb gazos. Rolul esential al hematiilor este transportul de oxigen catre
tesuturi.
Au
diametrul de 7,5-7,8 mm care scade
odata cu inaintarea in varsta a celulei, insumand o
suprafata totala de peste 2.500 m2, destinata
proceselor de schimb gazos. Rolul esential al hematiilor este transportul de oxigen catre
tesuturi.
La microscop au o culoare rosie-portocalie data de hemoglobina, iar numarul de hematii la om este de 5.100.000 ± 500.000/mm3 la barbati si de 4.600.000 ± 500.000/mm3 la femei. Acest numar poate creste atat in conditii fiziologice (la tineri legat de consumul ridicat in procesele de crestere), la altitudine in conditii de presiune atmosferica diminuata; in efort fizic sustinut cand se produce poliglobulia cat si in stari patologice (in bolile insotite de hipoxii tisulare ca insuficienta cardiaca sau tuberculoza pulmonara). In alte cazuri, numarul de hematii poate scadea, tot in conditii fiziologice (ingestia exagerata de apa provoaca o hemodilutie; persoaanele care muncesc in conditii de presiune atmosferica crescuta, deci de hiperbarism, ca minerii sau scafandrii) sau patologice (hemoragii, icter hemolitic, anemii prin deficit de formare).
Pentru medicina clinica o importanta deosebita au doua proprietati ale eritrocitelor: hemoliza si viteza de sedimentare (VSH).
Hemoliza, adica degradarea hematiilor, se produce sub influenta unor factori mecanici (agitare mecanica, ultrasunete), termici (incalzirea la 48°C), fizico-chimici (scaderea presiunii osmotice a solutiei de suspensie), chimici (actiunea substantelor tensioactive si cu efecte de solviti ai lipidelor) si biologici (veninul de sarpe, toxine bacteriene care actioneaza prin mecanisme enzimatice). In laborator, se utilizeaza testul rezistentei hemotilice in diagnosticul anemiilor hemolitice care este consecinta scurtarii vietii hematiilor prin hemoliza. Rezistenta hematiilor la agentii hemolitici depinde de varsta lor, cele batrane sunt mai putin rezistente.
Viteza de sedimentare (VSH) este inversul procesului stabilitatii de suspensie a acestora. Se masoara prin procedeul Westergren, in care, dupa doua ore de la recoltarea sangelui venos, se citeste limita de separare dintre hematiile sedimentate si plasma supraadiacente. Valoarea fiziologica este de 6-8 mm/ora la barbat si de 10-16 mm/ora la femeie. Dupa 24 de ore, limita de separare da valoarea hematocritului, sedimentarea eritrocitelor fiind totala. Sedimentarea este determinata de greutatea specifica a hematiilor (1100), care este superioara plasmei (1027), si variaza in raport cu numarul de hematii, fiind mai incetinita la poliglobuli, din care cauza barbatul are un VSH mai redus. Mai depinde de raportul proteinelor serice (hematiile suspendate intr-o solutie de globuline sedimenteaza mai rapid decat in solutiile de albumine). Ea creste in situatii cand predomina globulinele plasmatice cu molecule mai mari, denumite aglomerine, reprezentate in fapt prin fibrinogen si unele hepatoglobine. De aceea, masurarea VSH in clinica, in cursul bolilor infectioase, constituie un test de orientare asupra reactiei organismului. In bolile acute, cresterea sa denota un prognostic favorabil al reactiei imunologice prezente, iar in bolile cronice, cu componenta imunologica, ca tuberculoza sau reumatismul, viteza crescuta este un indiciu ca procesul este activ. Reducerea VSH ilustreaza stingerea reactiei imunologice. Trebuie insa retinut ca testul VSH nu evidentiaza prezenta infectiei, ci modul in care organismul raspunde la aceasta. Deci, VSH-ul este un test nespecific.
In conditii normale
- hematocrit 40-45 si incarcare normala cu hemoglobina (Hb)
- in 100 ml sange se afla in medie 15 g Hb deoarece 1 g Hb pura se
poate combina cu 1,39 ml oxigen, fiecare
decilitru de sange poate transporta ceva mai mult de 20 ml oxigen in
combinatie cu Hb.
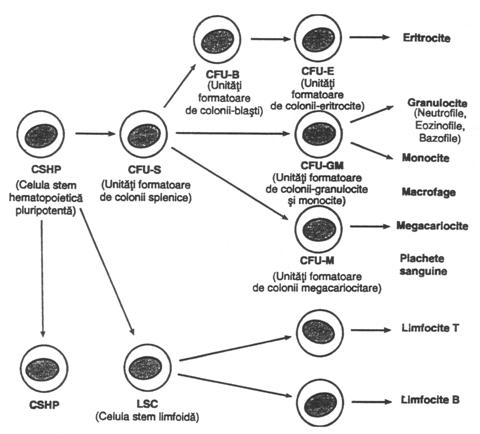
Fig. nr. 4.7. - Formarea diferitelor tipuri de celule sangvine periferice din celula stem hematopoietica (CSHP) aflata in maduva osoasa
Hematopoieza. In maduva oaselor se afla celule denumite celule stem pluripotente hematopoietice (CSPH), din care provin toate celulele sangvine, aspect ilustrat in fig. nr. 4.7., care prezinta diviziunile succesive ale acestor celule ce duc in final la formarea diferitelor elemente sangvine periferice.
Celulele stem se reproduc pe toata durata vietii individului si o buna parte dintre ele sunt retinute la nivelul maduvei osoase, unde constituie un fond celular care asigura continuitatea hematopoiezei. Dar, partea cea mai numeroasa a celulelor stem se diferentiaza, formand alte celule, mentionate in dreapta figurii 4.7. O parte dintre celule nu pot fi inca identificate ca tipuri celulare sangvine distincte desi sunt deja orientate spre un tip de celule sangvine. Acestea se numesc celule stem sau orientate. Cresterea si multiplicarea celulei stem este controlata de numeroase substante de natura proteica numite inductori de crestere. Sunt cunoscuti patru inductori principali, dintre care interleukina - 3 stimuleaza cresterea si reproducerea tuturor tipurilor de celule stem, iar celelalte induc numai cresterea anumitor tipuri de celule stem. S-a mai descris, de asemenea, si un alt set de substante proteice denumite inductori de diferentiere, care stimuleaza diferentierea celulara, prin determinarea mai multor diviziuni succesive, dand o celula sangvina adulta. Acesti factori de crestere si de diferentiere se afla sub controlul unor factori extramodulari.
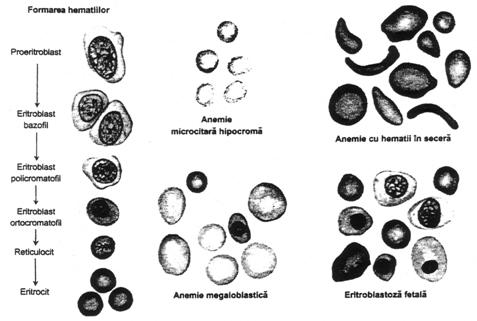
Se disting mai multe etape ale diferentierii eritrocitelor.
In conditii de stimulare corespunzatoare, din celulele stem se
formeaza un mare numar de protoblasti
(fig. nr. 4.8.) care se divid de mai
multe ori, formand numeroase eritrocite mature. In lungul acestor diviziuni,
celulele se incarca progresiv cu Hb pana la concentratia de 34
%. Nucleul se micsoreaza, iar in final va fi eliminat din
celula. In final, celule trec, prin diapedeza, in sangele capilar.
Fig. nr. 4.8. - Formarea si aspectul hematiilor in diferite tipuri de anemii
Eritropoieza este reglata de mai multi factori. Masa eritrocitelor din sange este mentinuta in limite foarte stricte ca sa nu impiedice circulatia, dar care sa asigure oxigenarea tesuturilor printr-un mecanism de control, redat in fig. nr. 4.9.
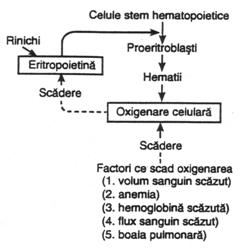

In acest mecanism, scaderea cantitatii de oxigen transportat catre tesuturi (anemie produsa de hemoragie, prezenta la altitudini mari etc.) creste productia de eritrocite (maduva hematogena incepe imediat sa produca cantitati mari de eritrocite). Deci oxigenarea tisulara este factorul primar al reglarii productiei de eritrocite. Rezulta ca nu numarul de eritrocite din sange este acela ce controleaza rata productiei de eritrocite, ci capacitatea functionala a acestora de a transporta oxigenul in tesuturi in functie de nevoile acestora pentru oxigen.
Ca raspuns la hipoxie, se formeaza un hormon (glicoproteina) circulant denumit eritropoietina, in lipsa caruia eritropoieza nu se produce, iar in prezenta sa hipoxia va creste productia de eritrocite. Eritropoietina se formeaza in proportie de 80-90 % in rinichi (iar diferenta in ficat), dar inca nu stim in care portiune a sa se formeaza.
Maturarea eritrocitelor este conditionata de prezenta vitaminei B12 si a acidului folic, care asigura formarea trifosfatului esential pentru sinteza ADN. In absenta lor, prolifereaza eritrocite de mari dimensiuni, numite megaloblasti, care au membrane subtiri. Acestia sunt capabili sa transporte normal O2, dar sunt foarte fragili, avand o viata scurta, 1/3-1/2 din normal.
Se produce un deficit de maturatie pe linia eritropoiezei, care are ca si cauza absorbtia insuficienta a vitaminei B12 in tractul gastro-intestinal. Ea apare in atrofia mucoasei gastrice. Consecinta este instalarea anemiei pernicioasa. In mod normal, glandele gastrice produc o glucoproteina, factorul intrinsec, care se combina cu vitamina B12 din alimente si astfel faciliteaza absorbtia sa in intestin.
Sinteza hemoglobinei. Incepe in proeritroblast, unde continua cateva zile, dupa care celulele parasesc maduva si trec in circulatia sangvina. Treptele principale ale formarii hemoglobinei sunt prezentate in fig. nr. 4.10.
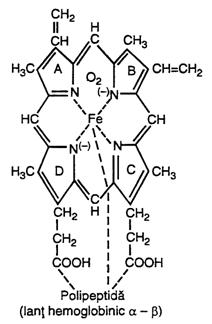 Principalele
etape ale formarii hemoglobinei sunt: (a) sinteza hemului care are un atom de fier; (b) hemul se combina cu un lant polipeptidic lung, denumit globina, formand o subunitate a Hb
care este lantul de hemoglobina
(redat in fig. nr. 4.11.), cu
greutate moleculara de 16.000; (c) se leaga 4 lanturi intre ele
formand o molecula mare de
hemoglobina.
Principalele
etape ale formarii hemoglobinei sunt: (a) sinteza hemului care are un atom de fier; (b) hemul se combina cu un lant polipeptidic lung, denumit globina, formand o subunitate a Hb
care este lantul de hemoglobina
(redat in fig. nr. 4.11.), cu
greutate moleculara de 16.000; (c) se leaga 4 lanturi intre ele
formand o molecula mare de
hemoglobina.
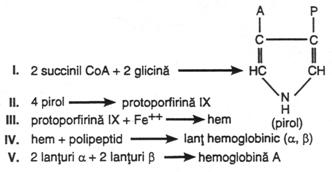

Fig. nr. 4.10. - Formarea hemoglobinei
Deoarece fiecare molecula de Hb are 4 atomi de fier, care pot lega fiecare o molecula de O2, rezulta ca o molecula de Hb poate transporta 4 molecule de O2 (8 atomi).
Cea mai importanta proprietate a Hb este capacitatea sa de a se combina slab si reversibil cu oxigenul. Se combina cu oxigenul in plamani, pe care il elibereaza (sub forma moleculara, nu atomica) in capilarele tisulare, unde presiunea partiala a oxigenului (pO2) este mai mica decat in plamani.
Metabolismul fierului. In organism exista aproximativ 4 g fier, din care 65 % intra in Hb, 4 % in mioglobina, 1 % in alti compusi hemici care realizeaza oxidarea intracelulara, 0,1 % este combinat cu transferina din plasma, iar 15-30 % este stocat sub forma de feritina, in principal in ficat.
Transportul, stocarea si metabolismul fierului sunt ilustrate in fig. nr. 4.12.
Fierul
se absoarbe in intestinul subtire, unde se combina imediat cu apotransferina, formand transferina, care va fi
transportata prin plasma, putand fi furnizat catre orice
celula din tesuturi, dar mai ales in celulele reticulo-enditeliale
sau hepatocite, unde se combina cu o apoferitina,
formand feritina, care
reprezinta rezerva de fier a
organismului. Cand scade cantitatea de fier din plasma, fierul este
usor eliberat din feritina, fiind transportat de transferina
plasmatica ori unde este nevoie in organism, dar ea se fixeaza
puternic pe receptorii membranei
eritroblastilor din maduva rosie hematogena, de unde
este ingerat in eritroblasti prin endocitoza. In eritroblasti,
transferina elibereaza fierul direct
mitocondriilor, deci acolo unde se produce sinteza hemului. La persoanele
care nu au cantitati adecvate de transferina, transferul
fierului catre eritroblasti este deficitar si se instaleaza
anemia hipocroma, in care
eritrocitele contin cantitati foarte mici de Hb. Cand hematiile
isi incheie ciclul de viata si sunt distruse, Hb se
elibereaza, fiind colectata de celulele sistemului
monocito-macrofagic, care ii elibereaza pentru a fi stocati de
catre feritina sau pentru a fi
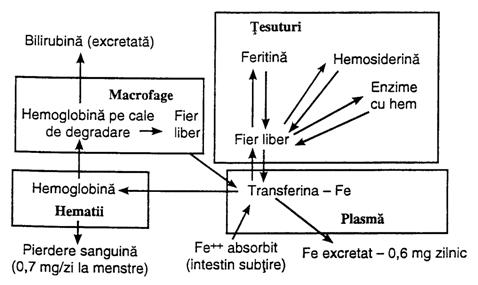
reutilizati la sinteza Hb.
Fig. nr. 4.12. - Transportul si metabolismul fierului
Zilnic, se pierd, in conditii normale, cca. 1 mg fier prin fecale, iar la femei se pot pierde 2 mg/zi prin menstruatie. Cantitati mai mari se pierd prin sangerare.
Controlul cantitatii de fier din organism se face de catre rata de absorbtie a acestuia. Cand organismul este saturat de fier, rata de absorbtie a acestuia in tractul gastrointestinal este mica, deoarece aproape toata apoferitina din rezerva este combinata cu fier, neputand accepta nici cea mai mica cantitate de fier din mucoasa intestinala.
Eritroliza. Durata medie de viata a eritrocitelor este de cca. 120 zile. La aceasta durata sistemul metabolic al hematiilor devine tot mai putin activ, iar celulele tot mai fragile, astfel ca la trecerea prin locuri mai stramte ale circulatiei se produc rupturi celulare. Fragmentele rezultate se aglomereaza in splina, printre trabecule. Aici Hb eliberata prin eritroliza este fagocitata imediat de microfagele din organism, mai ales in ficat (celulele Kupffer), splina si maduva osoasa. Dupa cateva ore, microfagelele elibereaza fierul din Hb inapoi in sange. Fierul este transportat catre transferina fie in maduva pentru a forma noi eritrocite, fie in ficat sub forma de feritina. Portiunea porfirinica a Hb este transformata de catre macrofage in bilirubina, ce ajunge in sange si apoi este secretata de ficat in bila.
Anemiile. Consta in pierderea prea rapida sau productia prea lenta a eritrocitelor.
S-au stabilit mai multe tipuri de anemie: posthemoragica (dupa o hemoragie rapida cand se pierd multe eritrocite); aplastica (consecinta aplaziei maduvei osoase cu scaderea capacitatii sale functionale de a produce eritrocite); megaloblastica (productie slaba de eritrocite cauzata de carenta in vitamina B12 si acid folic).
Efectele anemiilor sunt anumite modificari a compozitiei si proprietatilor sangelui (vascozitatea se reduce la jumatate), vasele tisulare se dilata, iar inima cunoaste o crestere marcanta. Aspectul este si mai sever in conditii de efort fizic. Inima nu mai poate pompa cantitati mult mai mari de sange decat in repaus. Drept urmare, apare hipoxia tisulara extrema si adeseori insuficienta cardiaca acuta.
In fine, leucocitele sangvine si celulele tisulare rezultate din ele intervin in combaterea unor agenti toxici si infectiosi fie prin distrugerea acestora prin fagocitoza, fie prin formarea de anticorpi.
Reprezinta unitati mobile din sange care fac parte din sistemul de aparare al organismului.
Sunt formate in parte de maduva rosie hematogena (granulocite, monocite), dar si de tesutul limfatic (limfocite si plasmocite).
Rolul lor este ca cele mai multe sunt transportate prin sange la zonele de inflamatie, fiind un aparator rapid si puternic impotriva oricarui agent patogen, cautand si distrugand orice agresor.
Caracteristicile leucocitelor. In sange se afla 6 tipuri diferite de leucocite: polimorfonucleare, neutrofile, polimorfonucleare eozinofile, polimorfonucleare bazofile, monocite, limfocite si ocazional plasmocite. La acestea, se adauga megacariocitul, ale carui fragmente devin plachete sangvine (trombocite). Polimorfonuclearele au aspect granular (fig. nr. 4.13.), din care motiv se numesc si granulocite sau polinucleare.
Granulocitele si monocitele au rolul de a proteja organismul impotriva microbilor prin ingestia lor (fagocitoza). Unele limfocite fixeaza agresorii specifici si ii distruge ca si granulocitele. Limfocitele si plasmocitele functioneaza in tandem cu sistemul imun. Plachetele au functia principala de a activa mecanismul coagularii sangvine.
Individul adult are aproximativ 7000 leucocite/mm3 de sange, in urmatoarele proportii: polimorfonuclearele neutrofile 62 %, limfocitele 30 %, monocitele 5,3 %, polimorfonuclearele eozonofile 2,3 %, polimorfonuclearele bazofile 0,4 %. Plachetele sangvine, care sunt numai fragmente de celule, 300.000/mm3.
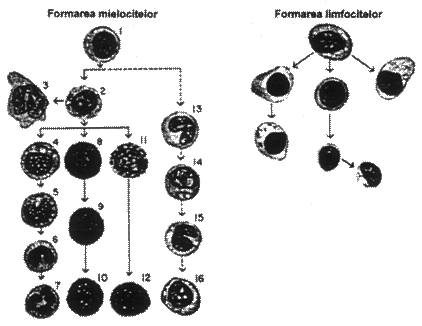

Leucopoieza. leucocitele provin din doua linii majore: linia mielopoietica si linia limfopoietica. Cele doua linii sunt prezentate in fig. nr. 4.13. prima in dreapta, a doua in stanga figurii. Prima linie incepe cu mieloblastul, iar a doua cu limfoblastul.
Locul de formare, de asemenea, este diferit: granulocitele si monocitele numai in maduva osoasa, celelalte in organele limfopoietice (ganglioni limfatici, splina, timus, amigdale si unele formatiuni limfoide din maduva oaselor si intestin).
Leucocitele formate in maduva sunt stocate aici pana cand sunt solicitate de sistemul sangvin, cand, datorita unor factori de eliberare, solicitarea este onorata. Sunt stocate in maduva de 3 ori mai multe granulocite decat se afla in circulatie (productia pe 6 zile).
Odata eliberate, granulocitele stau in circulatie 4-8 ore, iar in tesuturi 4-5 zile. In infectii durata lor este si mai mica (cateva ore), deoarece se indreapta catre zona infectata, unde sunt distruse in procesul protector respectiv. Monocitele au un timp de tranzit prin sange de 10-20 ore, dupa care trec in tesuturi, unde cresc in dimensiuni, devenind macrofage tisulare, in care faza traiesc luni sau chiar ani fara a fi distruse, indeplinind aici functii fagocitare. Limfocitele intra in circulatie odata cu drenajul limfei din ganglionii limfatici si dupa cateva ore trec in tesuturi, dupa care, prin intermediul limfei reintra in sange. Ele pot trai luni sau ani daca nu sunt solicitate de tesuturi.
Functia de aparare a neutrofilelor si a sistemului monocito-macrofagic. Agentii patogeni (bacteriile, virusurile) sunt atacate si distruse de neutrofile (celule mature cu capacitate de atac mare chiar in sange) si de monocite (celule imature, cu capacitate mica de actiune, dar numai in tesuturi, unde ajunse cresc mai mult de 5 ori, putand fi observate chiar si cu ochiul liber). Aceste ultime celule, mult dezvoltate, sunt numite macrofage si au capacitate foarte mare de a distruge agentii patogeni.
Neutrofilele si macrofagele se deplaseaza in tesuturi prin miscari amiboidale, uneori destul de rapide, 40 mm/minut. Ele pot fi atrase de numeroase substante eliberate in tesuturi - fenomenul fiind numit chemotaxie - care pot fi (1) unele toxine bacteriene, (2) produsi de degenerescenta eliberati de tesutul inflamat si (3) numerosi produsi de reactie.
Fagocitoza. Este functia cea mai importanta a fagocitelor, neutrofilelor si macrofagelor. Fagocitele au insa o actiune selectiva pentru a nu distruge celulele tisulare normale. Selectarea lor are la baza unele strategii: a) sunt atrase de particule cu suprafata rugoasa, ca a microbilor; b) prefera particulele fara invelisuri proteice protectoare, cum sunt cele ale tesuturilor moarte si a particulelor straine; c) recunosc particulele straine fata de cele ale propriului organism.
Fata de corpii straini selectati sintetizeaza anticorpi, care adera la membrana bacteriei, crescand sansa acesteia de a fi fagocitata.
Neutrofilele. Ajunse mature in tesuturi pot incepe imediat fagocitoza. Se apropie de particulele de fagocitat, lipindu-se de ele, trimitand o retea de pseudopode in jurul acestora. In interiorul citoplasmei neutrofilelor se formeaza o vacuola care a inglobat corpul strain, numita vezicula fagocitara, ce inainteaza in interiorul citoplasmei. Fiecare neutrofila poate fagocita 5-20 bacterii, dupa care sunt inactivate si mor.
Macrofagele. Odata activate de sistemul imun, macrofagele devin mai puternice decat neutrofilele, putand fagocita chiar 100 de bacterii. Pot sa inghita particule mai mari (neutrofilele numai cat o bacterie), pe care le fagociteaza, fara ca ele sa fie distruse dupa fagocitoza, ca neutrofilele, eliminand la exterior materialul fagocitat.
Particula fagocitata este contactata de unele granule citoplasmatice, mai ales de lizozomi, fuzionand cu vezicula fagocitara, care este inundata de o seama de enzime digestive. Ca urmare, vezicula fagocitara devine vezicula digestiva si imediat particula este digerata.
Sistemul reticulo-endotelial. Este constituit din celule diferite: monocite, macrofage mobile si macrofage fixe in tesuturi (nesolicitate in fagocitoza), unele celule endoteliale specializate din maduva osoasa) splina si noduli limfatici. Iata rolul acestora in fagocitoza.
Sinusurile medulare ale ganglionilor limfatici sunt captusiti de numeroase macrofage astfel incat orice particula care trece pe aici este fagocitata, impiedicand diseminarea particulelor in tot corpul.
In structura alveolelor pulmonare se afla, de asemenea, un mare numar de microfage tisulare care fagociteaza particulele ajunse in alveole, in special bacteriile din aerul inspirat.
Sinusurile hepatice sunt captusite cu macrofagele tisulare, numite celulele Kupffer, care este un sistem foarte eficient de filtrare a particulelor, astfel ca bacteriile din sangele port nu pot trece in circulatia sistemica.
Daca totusi vreo particula a scapat in circulatia generala actioneaza o alta linie de aparare, cea a macrofagelor din splina si maduva osoasa. Astfel, trabeculele pulpei rosii a splinei sunt captusite cu numeroase macrofage care au ca scop fagocitoza unor resturi din sange, in special eritrocitele imbatranite sau anormale.
Inflamatia. Este un complex de transformari tisulare consecutive afectiunii acestora de un atac bacterian, traumatic, chimic, caloric etc. In urma respectivului atac se elibereaza substante care produc transformari tisulare.
Un prim rezultat al inflamatiei este de a delimita zona inflamata de tesuturile din jur, prin cheaguri de fibrinogen, care impiedica raspandirea bacteriilor sau a produsilor toxici. Viteza de delimitare a zonei inflamate depinde de gradul de agresiune a bacteriilor. De exemplu, infectia stafilococica (care poate fi letala) se delimiteaza extrem de rapid fata de infectia streptococica, la care procesul de delimitare se desfasoara incet deoarece chiar daca bacteriile se raspandesc nu provoaca distructii tisulare atat de grave.
Macrofagele si neutrofilele isi manifesta imediat raspunsul la infectie. Prima linie de aparare o formeaza macrofagele. Cele mobile incep fagocitoza corpilor infectiosi in cateva minute, iar cele fixe se activeaza si devin mobile in cca. o ora.
A doua linie de aparare este constituita din neutrofile, care in timp de o ora trec din sange in zona infectata, datorita atragerii lor (chemotaxie) de catre anumiti produsi din focarul inflamator.
Paralel, se produce si un proces de neutrofilie, adica de crestere de 4-5 ori a numarului de neutrofile din sange (15.000-25.000 microlitru) pentru a fi puse la dispozitia tesuturilor. Cresterea se datoreste unor produsi de inflamatie care patrund in sange si care sunt transportati in maduva osoasa, care va elibera rezervele de neutrofile.
A treia linie de aparare este formata din monocite-macrofite. Numarul lor este mic in sange, cat si in maduva osoasa. De aceea, ele patrund mai incet in zona inflamata, dar dupa cateva zile devine tipul de celule dominant.
In sfarsit, a patra linie de aparare consta in cresterea productiei de granulocite si de monocite de catre maduva osoasa, care se produce insa abia dupa 3-4 zile de la declansarea infectiei. Cresterea poate fi chiar de 50 de ori mai mare decat in mod normal.
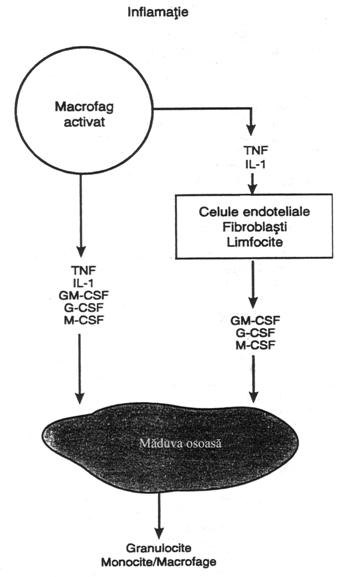 In controlul raspunsului, prin
feedback, al macrofagelor si neutrofilelor in infectie intervin vreo
25 factori, dintre care 5 au un rol predominant, prezentate in fig. nr. 4.14.
In controlul raspunsului, prin
feedback, al macrofagelor si neutrofilelor in infectie intervin vreo
25 factori, dintre care 5 au un rol predominant, prezentate in fig. nr. 4.14.
Acesti factori sunt: (1) factorul de necroza tumorala (TNF); (2) interleukina (IL-1); (3) factorul de stimulare a coloniilor de granulocite-monocite (GM-CSF); (4) factorul de stimulare a coloniilor de granulocite (G-CSF) ori (5) factorul de stimulare a coloniilor de monocite (M-CSF). Ei formeaza un mecanism de feed-back foarte puternic ce actioneaza initial in tesutul inflamat, continuand cu formarea celulelor albe de aparare si terminandu-se cu indepartarea cauzei inflamatiei ca urmare a activarii sistemului monocite-macrofage.
 Eosinofilele. Sunt fagocite slabe cu
manifestare chemotaxica si cu actiune incerta in
infectie. In cazul persoanelor cu infectie parazitara
numarul lor este mare, insa parazitii fiind prea mari nu pot fi
fagocitati. Totusi, ei se ataseaza de corpul
parazitilor, eliberand substante care ii omoara pe multi
dintre acestia.
Eosinofilele. Sunt fagocite slabe cu
manifestare chemotaxica si cu actiune incerta in
infectie. In cazul persoanelor cu infectie parazitara
numarul lor este mare, insa parazitii fiind prea mari nu pot fi
fagocitati. Totusi, ei se ataseaza de corpul
parazitilor, eliberand substante care ii omoara pe multi
dintre acestia.
S-a mai constatat ca eozinofinele se aglomereaza in zonele unde au aparut reactii alergice (tesuturile peribronhice de la asmatici, alergiile cutanate). Se crede ca ele detoxifica zona de unele substante toxice produse de alergie, prevenind raspandirea procesului.
Bazofilele. Elibereaza heparina - previne coagularea si creste viteza de indepartare din sange a grasimilor dupa un pranz gras - ca si mastocitele din apropierea capilarelor. Joaca un rol deosebit de important in unele tipuri de reactii alergice. Tipul de anticorpi IgE, care determina reactiile alergice, are tendinta predilecta de a se atasa de bazofile. Dupa ce antigenul a reactionat cu anticorpul, bazofilele se rup si elibereaza cantitati mari de histamina, tradikinina, heparina, care, la randul lor, produc reactii locale vasculare si tisulare, care reprezinta cauza menifestarilor alergice.
Leucopenia. Consta in incetarea productiei de leucocite, lasand corpul neprotejat de agentii invadatori. Astfel de cazuri apar la iradierea corpului cu raze gamma, la expunere la medicamente sau substante care contin benzi ori produsi antranici. In aceste cazuri cresc populatiile de bacterii din corp peste normal. In organismul nostru traiesc in simbioza multe bacterii (in gura - spinochete, pneumococi, streptococi, prezenti in numar mic si pe tractul respirator; in tractul gastro-intestinal - in special colibacili; se afla peste tot, chiar si in ochi). Scaderea de leucocite circulante permite invazia tesuturilor de bacterii, care dupa doua zile provoaca ulceratii bucale si in colon sau o infectie respiratorie severa. Bacteriile din ulceratii invadeaza tesuturile inconjuratoare si sangele. Fara tratament 3-6 zile pacientul poate muri.
Leucemiile. Sunt de doua feluri: limfoide si mieloide. Leucemiile limfoide sunt cauzate de o productie necontrolata, neoplazica de celule limfoide, initiata in tesuturile limfoide, mai ales in ganglionii limfatici. Leucemiile mieloide incep cu productia neoplazica de celule tinere mieloide din maduva osoasa. Dupa initiere, in ambele cazuri, celulele neoplazice se raspandesc in tot organismul. Acestea fiind celule nefunctionale, nu asigura o protectie normala impotriva infectiilor.
Primul efect al leucemiei este cresterea metastazica a leucocitelor in zone atipice organismului; invadand tesutul osos, respectiv splina, ganglionii limfatici, ficatul etc. si apoi tesuturile inconjuratoare. Alt efect principal este aparitia infectiilor, anemie severa, tendinte de sangerare in urma reducerii trombocitelor. Dar, cel mai drastic efect este utilizarea excesiva a materialului metabolic de catre celulele canceroase in crestere.
Sunt cele mai mici elemente figurate si nu au o structura celulara propriu-zisa, reprezentand fragmente citoplasmatice de 2-5 m
In granulatiile mai mari sunt prezenti: factorul F3 plachetar, serotonina cu rol vasoconstrictor, ADP, ATP, avand o membrana lipoproteica, putand prezenta cativa microtubi cu rol contractil.
Se formeaza in maduva rosie hematogena din celula susa numita hemocitoblast, care sub actiunea unui factor tumoral, numit trombopoietina, trece in forma de megacorioblast (celula giganta), care, prin maturare, rezulta megacariocite de gradul I, II si III. Ultimele, prin fragmentarea citoplasmei elibereaza placutele sangvine - circa 250.000 - 400.000 - prin mecanisme neuro-umorale.
Placutele sangvine au capacitatea de a forma agregate, sub influenta Adp-ului si trombinei. In contact cu suprafetele rugoase si umectabile emit o serie de prelungiri subtiri si lungi; aderand la acestea, dezagregandu-le. In anumite conditii, ca in hemoragii, trombocitele adera cu usurinta pe suprafete de colagen denundate, fiind un factor de reparare a endoteliului capilar. Scaderea lor sub 60.000 perturba procesul de hemostaza, iar sub 20.000 determina hemoragii spontane automate sau mucoase, care se combat prin transfuzii de sange. Persista in circulatie circa 4-15 zile dupa care sunt distruse.
Hemostaza. Este procesul fiziologic prin care organismul intervine in oprirea hemoragiei, produsa prin leziune vasculara, prevenindu-se pierderea sangelui din vasele lezate.
In hemostaza au rol important: vasul sangvin, placutele si procesul de coagulare. Hemostaza are trei faze.
Faza vasculara. Consta in modificari vasculare rapide si constau in spasmul musculaturii netede a vasului lezat, cu reducerea consecutiva a lumenului. Inchiderea vasului lezat se realizeaza prin mecanisme reflexe si umorale. (1) Reflex, lezarea vasului stimuleaza terminatiile nervoase libere care conduc impulsurile nervoase pana in coarnele laterale ale maduvei spinarii, in centrii nervosi simpatici. Pe calea simpatica se produce dupa 1 minut constrictiile vasului lezat, care dureaza cateva minute. (2) Umoral, se produce constrictia vasului prin serotonina eliberata din trombocitele dezagregate, cu efecte de mai lunga durata. In vasele mici (capilare) inchiderea se realizeaza prin retractia elastica a vasului lezat, mecanism insa insuficient pentru vasele mari.
Faza plachetara. Trombocitele adera la suprafata vasului lezat, organizandu-se in mai multe straturi succesive, formand un dop de cheag trombocitar, care include si alte elemente figurate. Din trombocite se elibereaza factorul F3 trombocitar, care impreuna cu tromboplastina tisulara, initiaza coagularea, proces favorizat de ADP si Ca2+ din placute si tesuturi.
Faza de coagulare. Are loc dupa 15-20 s si este totala la 1-3 minute, cand se formeaza un dop ce ocluzioneaza vasul dupa 3-6 minute. Este urmata de retractia cheagului la 30-60 minute prin scurtarea prelungirilor emise de placute, care este un proces activ, cu consum de energie, furnizata de ATP. Prin retractia cheagului, se separa partea lichida (serul) de cheagul propriu-zis, iar buzele plagii se apropie. In momentul urmator, din sange intra si alte elemente figurate (celule mobile) neutrofile, limfocite, care se transforma in monocite macrofage. Se produce organizarea fibroasa a cheagului. Pe filamentele de fibra se absoarbe plasminogen, care, sub influenta unor activatori locali, se transforma in plasmina, ce produce liza fibrinei. In decurs de cateva zile apare o cicatrice fibroasa reparatorie si se reface continuitatea vasului.
Procesul de hematoza poate deveni nociv pentru peretii vaselor pentru ca apar leziuni care ii fac rugosi si umectabili (leziuni de ateroscleroza), cu depuneri de lipide pe pereti. Aceste zone initiaza adeziunea si aglomerarea de trombocite in zona lezata, putand duce la formarea de trombi, cu rol obliterant. Trombii mobilizati in circulatie reprezinta embolii. Daca acestia ajung in plamani pe calea circulatiei venoase, declanseaza o serie de reflexe si tulburari mecanice circulatorii de ocluzie arteriala pulmonara, care dau tabloul emboliei pulmonare, a carei gravitate poate duce la moarte.
Copyright © 2026 - Toate drepturile rezervate