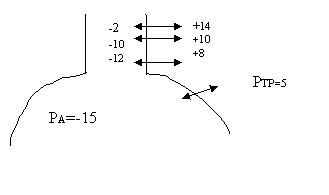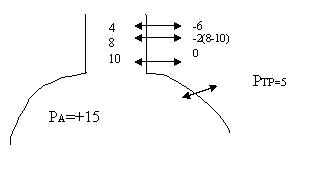|







 |  |
|
| |
 | Alimentatie | Asistenta sociala | Frumusete | Medicina | Medicina veterinara | Retete |
FIZIOLOGIA APARATULUI RESPIRATOR
Prin respiratie se intelege schimbul de oxigen si bioxid de carbon intre organism si aerul atmosferic.
Consumul de oxigen si eliberarea de bioxid de carbon au loc in cadrul proceselor oxido-reducatoare de la nivel celular. Intre celule si mediul extern se interpun doua sisteme:
Sangele din aparatul cardio-vascular, care transporta gazele respiratorii spre si de la plamani;
Membrana alveo-capilara din structura plamanului, prin care are loc schimbul acestor gaze intre sange si aerul atmosferic.
Aerul din apropierea membranei alveo-capilare este permanent preschimbat prin procesul ventilatiei, iar sangele este pompat de ventriculul drept si este adus pe calea circulatiei pulmonare.
Studierea respiratiei se face in trei etape:
1.Respiratia interna reprezentata de totalitatea proceselor oxidative de la nivel celular. Ele sunt studiate la biochimmie.
2.Transportul oxigenului si bioxidului de carbon prin sange;
3.Respiratia externa sau pulmonara in cadrul careia se face schimbul celor doua gaze intre sange si aer.
Respiratia pulmonara are loc la nivelul celor doi plamani, situati in cutia toracica si formati din lobi: 3 in plamanul drept si 2 in plamanul stang. Plamanii sunt inveliti de o seroasa (pleura) formata din doua foite:
a) parietala, ce acopera si adera de suprafata interna a cutiei toracice;
b) viscerala, ce acopera si adera de plaman, patrunzand in scizurile dintre lobi.
Intre cele doua foite exista un strat subtire de lichid (cca 10 ml), secretat de vasele foitei parietale. El permite alunecarea plamanilor in timpul actului respiratiei, dar realizeaza si aderarea dintre cele doua foite, deci a plamanului de cutia toracica.
Toracele si plamanul, sunt doua structuri elastice cu tendinte opuse:
Plamanul de a se retracta, el fiind tractionat de cutia toracica (deschiderea cutiei toracice - pneumtorax - determina colabarea plamanului, dar si marirea de volum a cutiei toracice);
Cutia toracica de a-si mari volumul, fiind tractionata de plaman ( in emfizem, cand elasticitatea plamanului scade, cutia toracica isi mareste volumul).
Cele doua structuri elastice se gasesc in echilibru la sfarsitul unei expiratii de repaos, dar fiecare avand tendinta de retractie, creeaza in spatiul pleural o presiune mai mica decat cea atmosferica, numita presiune negativa, cu o valoare de 2,5mm Hg (4-5 cm H2O). Dovada acestei presiuni negative este ca, in momentul introducerii unui ac in spatiul pleural, aerul atmosferic patrunde in acest spatiu (pneumotorax).
Aceasta presiune negative este mentinuta de absorbtia continua a lichidului pleural de vasele limfatice din pleura viscerala si, dupa cum vom vedea, se modifica cu miscarile respiratiei si cu pozitia corpului.
Componenta respiratorie a plamanilor este reprezentata de caile respiratorii. Ele incep in fosele nazale si au o componenta extrapulmonara, reprezentata de fosele nazale, faringe, laringe, trahee si cele doua bronhii principale. Traheea si bronhiile prezinta cartilagii in potcoava ce impiedica colabarea. In portiunile libere de cartilagii se gaseste musculatura neteda, a carei contractie creste viteza de expulzie a aerului in timpul expectoratiei din tuse.
Bronhiile principale patrund in plamani la nivelul hilului si se divid in doua, apoi iar in doua,etc. prezentand cca 23 de diviziuni successive numite generatii ( generatia 0 - traheea, generatia 1- cele 2 bronhii principale, etc.) In primele 10 generatii, bronhiile prezinta cartilagii in pereti. Din generatia a 11-a cartilagiile dispar si capata denumirea de bronhiole, cu un diametru sub 1mm.
Stratul muscular devine tot mai bine reprezentat. Bronhiolele sunt mentinute deschise prin ancorarea de structurile din jur si prin existenta in jurul lor a unei presiuni negative. Ele sunt susceptibile colabarii in timpul expiratiei. Aproximativ generatia a 16-a este reprezentata de bronhiole terminale.
Intreg acest segment (inclusive bronhiolele terminale) al arborelui bronsic joaca rol de conducte. Are un volum de cca 100-150 ml si se numeste spatiu mort, pentru ca nu participa la schimburi respiratorii. Epiteliul acestui segment este simplu cubic, cu numerosi cili, ce bat spre faringe. In epiteliu se gasesc celule care secreta mucus. Acesta e secretat si de glandele submucoase si acopera intreaga suprafata a mucoasei. In acest segment aerul inspirat este incalzit, umidificat si filtrat deoarece diferitele particule sunt fixate in mucus, iar cilii imping spre faringe mucusul cu particule inglobate in el.
Aproximativ la generatia 17 incep sa apara alveole in peretii bronhiolelor, ele capatand denumirea de bronhiole respiratorii, densitatea alveolelor crescand spre generatia 19. Generatiile 20-22 se numesc ducte alveolare, au peretii complet acoperiti de alveole. Ductele se termina in sacii alveolari (generatia 23).
Structurile ce deriva dintr-o bronhiola terminala formeaza unitatea respiratorie terminala sau acinul pulmonar (cca 60.000 in cei 2 plamani) si datorita prezentei alveolelor reprezinta regiunea in care se fac schimburile respiratorii. Ele sunt umplute cu aerul alveolar, care corespunde CRF (capacitatea reziduala functionala), avand un volum de cca 2.300ml in timpul respiratiei de repaos.
Suprafata de sectiune a traheei este de 2,5cm2, ea ajunge la 180 cm2 la nivelul bronhiolelor terminale. Este cauza scaderii vitezei de deplasare a aerului. Pana la nivelul generatiei 16 deplasarea aerului se face prin convectie si viteza scade treptat pana la nivelul acinului, unde difuziunea gazelor devine mai rapida decat convectia.
Schimbul de gaze intre sange si aer are loc la nivelul alveolelor - structuri emisferice, cu un diametru de 75-300 µm. Numarul lor este de 300 mil in cei 2 plamani. Au o suprafata de 50-100 m2 si un volum de 5-6 l (alveolele maresc foarte mult suprafata - o sfera cu un volum de 5,5l si are o suprafata de 1,6 mp. Peretele alveolei este format dintr-un strat de celule epiteliale - pneumocite de tip I si de tip II. Cele de tip I sunt mai plate, cele de tip II sunt cuboidale si sintetizeaza surfactantul, depozitat in corpii lamelari. Pneumocitul de tip I acopera 90-95% din suprafata si reprezinta calea cea mai scurta de difuziune a gazelor respiratorii. In caz de lezare a pneumocitului de tip I, cele de tip II prolifereaza restabilind continuitatea epiteliului.
Alveolele sunt inconjurate de o retea de capilare. Membrana alveo-capilara e formata din epiteliul alveolar, cele 2 membrane bazale si endoteliul capilar, iar in cazul pneumocitului de tip I are o grosime de 0,15-0,30 µm. Capilarele din jurul alveolelor provin din circulatia pulmonara. Au un diametru de 8 µm si un eritrocit strabate un capilar in repaos in 0,75 sec. In peretele alveolar se mai gasesc macrofage alveolare, cu rol de aparare. Ele capteaza si degradeaza particulele ajunse la acest nivel. Cand particulele sunt in numar mare, macrofagele elibereaza enzime producand inflamatie si distructii tisulare. La fumatori, prin eliberarea elastazei din macrofage, este distrus tesutul elastic, ceea ce duce la emfizem. In peretele alveolar se mai intalnesc si limfocite, plasmocite si mastocite. Acestea din urma contin heparina, histamine, polipeptide ce participa la reactiile alergice.
Fortele ce participa in timpul acestor miscari sunt studiate de mecanica respiratiei.
Respiratia externa sau pulmonara presupune:
ventilatia pulmonara;
perfuzia pulmonara;
difuziunea O2 si CO2 prin membrane alveo-capilara.
Ventilatia pulmonara consta in preschimbarea aerului alveolar. Se realizeaza prin miscarile respiratorii de inspiratie si expiratie, miscari ce se desfasoara fara pauza, raportul dintre expiratie si inspiratie fiind de 1,3.
Volumul total al cailor respiratorii este de 5-6 l. Fiziologii au stabilit o serie de volume, care nu corespund unei anumite regiuni anatomice, dar determinarea lor aduce informatii utile pentru aprecierea starii si evolutiei unei afectiuni respiratorii. Aceste volume sunt:
a.Volumul respirator curent (VC) - reprezinta aerul care intra si iese din plamani in timpul unei respiratii de repaos. El este de aproximativ 500 ml sau 15% din capacitatea vitala (CV);
b.Volumul inspirator de rezerva (VIR) - reprezinta aerul care patrunde in plaman intr-o inspiratie maxima, dupa o inspiratie de repaos; reprezinta cca 60% din CV;
c.Volumul expirator de rezerva (VER) - reprezinta aerul care se elimina din plaman intr-o expiratie fortata, dupa o expiratie de repaos, precedata de o inspiratie de repaos. Reprezinta cca 25% din CV.
d.Volumul residual (VR) - aerul care ramane in plaman dupa o expiratie maxima.
Combinatia volumelor amintite poarta denumirea de "capacitati". Acestea sunt:
Capacitatea inspiratorie : VC+VIR;
Capacitatea expiratorie: VC+VER;
Capacitatea reziduala functionala (CRF): VR+VER;
Capacitatea vitala: VC+VIR+VER.
Capacitatea totala: VC+VIR+VER+VR
In mod current se determina CV, pentru a urmari evolutia unei afectiuni pulmonare. Un alt parametru care se determina este Volumul expirator maxim pe secunda (VEMS), detrminat printr-o inspiratie maxima urmata de o expiratie cu maxim de efort. Volumul expirat in prima secunda se numeste VEMS si reprezinta 70-80% din CV. Acest procent se numeste indice de permeabilitate bronsica (IPB). Determinarea volumelor si capacitatilor amintite se face in sala de lucrari.
Ventilatia este realizata prin miscari de inspiratie si expiratie.
Inspiratia este partea activa a respiratiei, deoarece este realizata prin contractia muschilor inspiratori. In inspiratia de repaos se contracta:
m. diafragm;
m. intercostali externi.
Muschiul diafragm are forma unei emisfere cu convexitatea orientata in sus. El se insera pe un centru tendinos si pe coaste. Are o suprafata de cca 270 cm2 .Prin contractie coboara, marind diametrul longitudinal al cutiei toracice cu cca 1,5cm, ce corespunde unui volum de cca 400ml.Contractia lui e comandata prin nervii frenici ce deriva din coarnele anterioare C3-C5.
Muschii intercostali externi ridica si rotesc coastele impingand sternul inainte si in sus. In acest fel maresc diametrul antero-posterior si transversal.
Intr-o inspiratie fortata intervin muschii inspiratori accesori - scaleni, sternocleidomastoidieni, pectorali, dintati, care, in mod normal au insertia fixa pe cutia toracica si cea mobila pe oasele capului, gatului si coloanei vertebrale. Contractia lor determina miscari ale capului si gatului. In inspiratia fortata ei isi schimba insertiile: cea fixa pe oasele capului si gatului si cea mobila pe coaste, marind volumul cutiei toracice. In acelasi timp deplasarea diafragmului se poate face pe o distanta de pana la 15 cm, m.diafragm fiind principalul muschi inspirator.
Expiratia este componenta pasiva a respiratiei. In expiratia de repaos muschii inspiratori se relaxeaza si toate structurile elastice tensionate in inspiratie isi revin. In expiratia fortata intervin muschii accesori, principalii fiind muschii abdominali. Prin contractia lor presiunea din abdomen creste si diafragmul este impins in sus de viscerele abdominale.
Fortele care participa in timpul acestor miscari sunt studiate la mecanica respiratiei.
Presiunea din cutia toracica si din alveole
Cutia toracica e considerata spatiul dintre plaman si cutia toracica si se refera la spatiul intrapleural, spatiul dintre plamani si diafragm si la mediastin si spatial interstitial din plaman, in afara cailor respiratorii, a vaselor sanghine si limfatice.
La sfarsitul unei expiratii de repaos, cele 2 structuri elastice plamanul si cutia toracica, unite prin spatial intrapleural, sunt in echilibru:
Ele au tendinte opuse:
plamanul de a se retracta;
cutia toracica de a expansiona.
Din aceasta cauza in spatiul intrapleural se creaza o presiune negative cu o valoare de -5 cm H2O. Aceeasi presiune negativa se gaseste si:
in spatiul dintre plaman si diafragm;
in mediastin;
in spatiul interstitial din jurul cailor respiratorii, a vaselor de sange si limfatice.
Ea se numeste presiune intratoracica (PIT) si poate fi masurata prin introducerea in spatiul intrapleural a unui ac conectat cu un manometru sau prin introducerea unui balonas in partea mediastinala a esofagului, balonasul fiind conectat cu un manometru. Presiunea intratoracica este influentata de pozitia corpului fata de actiunea fortei gravitationale. In ortostatism, forta gravitationala atrage plamanul in jos, PIT la varful plamanului scade la sfarsitul expiratiei la -10 cm H2O, iar la baza plamanului creste la -2,5cm H2O (pentru simplificare se va tine seama de aceste modificari). PIT este influentata si de miscarile respiratorii. In inspiratia de repaos scade la -7,5 cm H2O ( se introduce 500 ml aer VRC), in inspiratia fortata are poate ajunge la -10..-15 cm H2O. In expiratia fortata PIT are valori positive de +10..+15 cm H2O.
Importanta este si presiunea transpulmonara (PTP), adica presiunea rezultata din insumarea celor 2 forte ce se exercita pe membrane alveolara: PIP si PA (presiunea alveolara). Deoarece au tendinte opuse PTP = PA - PIP. Valoarea pozitiva indica distensia alveolara, valoarea negative indica colabarea.
In conditii statice, adica oprirea respiratiei cu glota deschisa presiunea din toate caile aeriene, dar si in alveole este cea atmosferica, adica zero.In aceste conditii PTP = 0 - PIP , deci PTP = -PIP . De exemplu la sfarsitul unei expiratii de repaus PIP = -5, PTP = -(-5), deci 5 cm coloana de apa. PTP este forta care invinge reculul elastic al plamanului. Aceasta se poate demonstra introducand un ac in spatiul intrapleural. Aerul patrunde in acest spatiu unde presiunea este negativa. In acest timp, desi de cele 2 parti ale peretelui alveolar presiunea este aceeasi (cea atmosferica), alveolele colabeaza.
Fortele care se opun distensiei pulmonare si care tind sa colabeze alveolele sunt:
tesutul elastic din structura plamanului (fibre de elastina si colagen);
tensiunea superficiala care se exercita la interfata dintre lichidul ce abduce suprafata interna a alveolelor si aerul din alveole.
2. Rezistenta pulmonara este determinata de;
- rezistenta opusa de caile respiratorii la trecerea aerului;
- rezistenta opusa de structurile din plaman care se freaca intre ele in timpul miscarilor respiratorii. Cele ce au valoarea cea mai mare sunt fortele elastice, tensiunea superficiala reprezentand 2/3 din valoarea lor.
Tensiunea superficiala este determinata de atractia intre moleculele de apa de pe suprafata lichidului care abduce alveolele. Aceasta atractie reprezinta o forta care actioneaza paralel cu suprafata lichidului, tinzand sa colabeze alveolele. Ea are o valoare de 50 dyne/cm (in cazul apa-aer = 70 dyne/cm).
Tensiunea superficiala este redusa de surfactant .
Surfactantul este secretat de pneumocite de tip II si de celulele Clara din mucoasa bronsiolelor. Este un amestec format din lipide (90%), proteine (10%) si Ca++ . Jumatate din lipide sunt reprezentate de dipalmitoil lecitina, exista si fosfatidilcolina. Proteinele sunt 4 apoproteine: SPA, SPB , SPC si SPD . SPB si SPC favorizeaza patrunderea surfactantului la suprfata lichidului si raspandirea lui pe suprafata. Componenta lipidica patrunde din sange, cea proteica e sintetizata de celule.
Surfactantul se dispune pe suprafata lichidului cu gruparile hidrofile spre lichid, cele hidrofobe spre aer interpunandu-se intre moleculele de apa. In acest fel scade tensiunea superficiala de la 50 la 5-30 dyne/cm si deci scade forta necesara distensiei alveolare. Cu cat densitatea surfactantului de pe suprafata lichidului este mai mare cu atat scadrea tensiuii superficiale este mai mare. Surfactantul se secreta in timpul vietii intrauterine, iar secretia lui creste mult inainte de nastere, sub actiunea cresterii glucocorticoizilor din sangele matern.
La copiii care se nasc prematur, surfactantul poate fi in cantitati mici si prezinta dificultati in distensia pulmonara.
Prezenta surfactantului are urmatoarele consecinte:
Conform legii Laplace P= 2T/R, rezulta ca pe masura scaderii razei unei alveole, presiunea necesara mentinerii deschise a alveolei trebuie sa creasca. In plamani presiunea in toate alveolele este aceeasi, deci ar urma ca alveolele mici sa colabeze, cele mari sa creasca in volum.
Surfactantul din alveolele mari se distribuie pe suprafata lichidului, lasand spatii de mai multe molecule de apa, deci scade mai putin tensiunea superficiala, in cele mici stratul de surfactant este aproape continuu, scazand mai mult tensiunea superficiala.
Aceasta distributie a surfactantului permite mentinerea tuturor alveolelor deschise deci stabilizeaza alveolele. Surfactantul are si rolul de a uniformiza dimensiunea alveolelor deoarece:
daca in timpul inspiratiei o alveola este mai bine ventilate deci patrunde mai mult aer si se destind mai mult, surfactantul intinzandu-se pe o suprafat mai mare scade viteza de distensie a alveolei;
in alveolele in care patrunde aerul mai incet, deci care se destind mai putin stratul de surfactant este mai mare ceea ce scade mai mult tensiunea superficiala favorizand distensia.
Un alt rol al surfactantului este cel de scadere a filtrarii la nivelul capilareor (scade presiunea negativa, opunandu-se instalarii edemului pulmonar din interstitiu).
PTP este forta care determina valoarea volumului pulmonar.
La PTP = 5 cmH2O (sfarsitul expiratiei de repaos) volumul plamanului este egal cu CRF (capacitatea reziduala functionala) adica 2500ml.
La PTP = 15 cm H2O (inspiratie fortata) volumul pulmonar ajunge la CT (capacitatea totala) adica 5000-5500 ml.
Relatia dintre PTP si volumul pulmonar ne da indicatii asupra distensibilitatii pulmonare.
In acest scop se determina complianata pulmonara, adica variatia de volum a plamanului (ΔV) pentru o variatie de PTP de 1cm H2O (ΔP=1cm H2O). Variatia de volum se obtine inspirand dintr-un spirometru o cantitate cunoscuta de aer (de ex. 500 ml). Variatia de PTP se determina introducand un balonas in partea mediastinala a esofagului, care determina PIP .
Sa presupunem ca inainte de introducerea aerului PIP =-5, iar dupa introducerea celor 500 ml = -7,5. Cum determinarea acestor presiuni se face in conditii statice cand PA =0, PTP va fi de -(-5) adica 5 la inceputul inspiratiei si -(-7,5) adica 7,5 la sfarsit, deci pentru ΔV =500 ml, ΔP =2,5cm.
ΔV/ ΔP = 500/2,5 = 200/1
Complianta in acest caz este 200 ml, si aceasta este valoarea normala a compliantei.
|
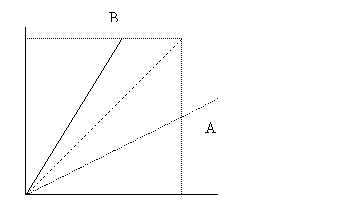
Scaderea compliantei (A) inseamna o distensibilitate redusa (fibroza pulmonara), cresterea compliantei (B) - distensbilitate crescuta (emfizem pulmonar).
P
IIi iIn conditii dinamice, adica in timpul miscarilor respiratorii, intervine rezistenta pulmonara, care se opune distensiei, si care fiziologic are o valoare mica, 80% din ea fiind determinata de rezistenta opusa de caile respiratorii. Si in cazul cailor respiratorii, ca si in cazul vaselor de sange, se aplica legea lui Poissaulle: R=8lD/πr4 , adica rezistenta este invers proportionala cu raza la puterea a 4-a, rezistenta fiind mult influentata de calibrul cailor aeriene. In caile respiratorii superioare calibrul nu se modifica din cauza prezentei tesutului cartilaginos in pereti. In bronhiolele in care in pereti exista musculatura neteda, calibrul este variabil, fiind influentat activ sau pasiv de mai multi factori. Interconectarea intre compartimentele pulmonare determina in timpul maririi de volum a cutiei toracice, distensia inclusive a bronhiolelor.
In aceste procese intervin si factori chimici:
acetilcolina (mediator parasimpatic) contracta bronhiolele
noradrenalina (mediator simpatic) bronhiolodilatator
adrenalina, bronhiolodilatator
histamina (secretata de mastocite in stari alergice) contracta ductele alveolare.
Un alt factor care influenteaza calibrul bronhiolelor este volumul de aer existent in plamani in timpul miscarilor respiratorii.
Un criteriu de apreciere a rezistentei cailor aeriene este determinarea debitului de aer (fluxului) prin aceste cai. In afara de calibrul cailor aeriene, sensul si marimea fluxului depind de valoarea PA, aceasta fiind determinata de:
PIP ;
Reculul elastic, direct proportional cu distensia alveolara;
Cand volumul de aer din plamani este mare, in timpul unei expiratii fortate, ambii factori au valori mari, deci PA este mare si fluxul e ridicat.
Cand volumul scade si cei 2 factori scad, scade PA si fluxul scade.
Aceasta relatie intre flux si volumul pulmonar se stabileste prin determinarea buclei flux-volum. In acest scop se inspira maxim deci in plaman se introduce un volum de aer corespunzator CV (capacitatii vitale), apoi se expira cu forta maxima. Se observa la inceput o crestere pronuntata a fluxului, apoi pe masura scaderii volumului pulmonului fluxul scade.
|
V |
c Calibrul bronsiolelor depinde de fazele respiratorii. In inspiratie bronsiolele se dilate, in expiratie au tendinta de colabare. C |
75%cv 50%cv 25%cv 0cvC I
I,m
IIn acest sens se poate folosi urmatorul exemplu
|
PIP=-20 |
In inspir profund PA scade la -15, PIP=-20, PTP=5 In caile respiratorii presiunea creste ajungand la 0 la nivelul buzelor. Calculand presiunea exercitata pe peretele bronsic (de ex. -12 - (-20)=+8 se constata valori positive care indica dilatarea bronsiolelor. |
INSPIR
IIn expir fortat PA =+15, PIP =+10, PTP =+5
Presiunea din caile respiratorii scade treptat ajungand la 0 la nivelul buzelor
|
PIP=+10 |
Calculand presiunea exercitata pe peretele bronsic se obtin valori negative, deci colabare. Valorile negative apar dupa depasirea punctului de egalare al presiunilor. |
In mod normal la volumul de 5l, in timpul expirului fortat punctual de egalare al presiunilor se situeaza in caile respiratorii superioare (trahee), necompresibile. Daca volumul de aer din plaman este scazut acest punct se deplaseaza spre alveole si deci ajunge la nivelul bronsiolelor, depasirea lui produce colabare deci imposibilitate de expirare. Deplasarea punctului de egalare spre alveole are loc si in cazul spasmului bronsiolelor (spasm bronsic) . In acest caz scaderea razei duce la o crestere foarte mare a rezistentei (de 12 ori) ceea ce scade mult si repede presiunea din bronsiole apropiind punctual de egalare al alveolelor, ceea ce duce in timpul unui expir fortat la colabarea bronsiolelor.
Daca ne referim la bucla flux-volum, colabarea este marcata de o scadere brusca a fluxului.
Ventilatia alveolara consta in improspatarea aerului alveolelor cu aerul atmosferic in timpul miscarilor respiratorii. Cu fiecare respiratie de repaos se introduc in plaman 500 ml aer (VRC) din care cca 150 ml raman pe caile respiratorii, in spatiul care nu participa la schimburi, denumit spatiul mort anatomic. In acini ajung cca 350ml de aer atmosferic, improspatand cei 2500ml de aer alveolar cu fiecare inspiratie. Daca frecventa respiratorie este de 12x350=cca 4,2l/min. Cu cat frecventa respiratorie este mai mare cu atat este ventilat mai mult spatiul mort si mai putin aerul alveolar.
PERFUZIA PULMONARA
Irigatia plamanului este nutritiva si functionala.
Irigatia nutritiva este asigurata de arterele bronsice, ramuri ale aortei descendente, deci apartin circulatiei sistemice. Ele se ramifica in paralel cu bronsiile si iriga bronsiile, septurile, pleura parietala. Se capilarizeaza si apoi formeaza doua plexuri:
peribronsic;
submucos.
Plexul peribronsic, prin vena Azygos, se varsa in venele circulatiei mari, cel submucos se varsa in venele pulmonare si de aici in atriul stang, impurificand cu bioxid de carbon sangele oxigenat venit din plaman (resprezinta cca 1-2% din debitul cardiac). Circulatia functionala este asigurata de artera pulmonara, care pleaca din ventriculul drept aducand sange venos bogat in CO2 .
Patul vascular pulmonar se caracterizeaza printr-o rezistenta scazuta deoarece:
vasele sunt scurte;
peretii arterelor sunt subtiri, cu putin tesut elastic in peretii arteriolelor, tesutul muscular este slab reprezentat, de aceea presiunea in circulatia pulmonara nu scade brusc la nivelul arteriolelor;
capilarele nu au sfinctere precapilare;
venele sunt foarte distensibile.
Rezistenta la curgerea sangelui fiind scazuta si presiunea in artera pulmonara este mica:
Pmax = 25 mm Hg;
Pmin = 8-10 mm Hg;
Pmedie = 15 mm Hg;
In capilare: P = 7mm Hg; in vene 2-4 mm Hg, apropiata de cea din atriul stang. Debitul este acelasi ca in circulatia sistemica de 5,5l/min in repaus. Volumul sanguin de 450-500 ml (9% din volumul total). Volumul in capilare de cca 100 ml, creste la 200 ml in clinostatism (dispnee clinostatica).
Creterea presiunii in circulatia pulmonara nu se datoreaza cresterii rezistentei ci mai ales unui obstacol la curgerea sangelui (stenoza mitrala, insuficienta ventriculara stanga).
Controlul circulatiei
Influenta sistemului nervos vegetative este redusa. SNS are rol de mobilizare a sangelui din circulatia pulmonara (daca se penseaza experimental artera pulmonara, sangele mobilizat din plaman mai permite 2 contractii ventriculare.
Controlul circulatiei este in primul rand automat, adaptand perfuzia la ventilatie. Scaderea presiunii oxigenului din alveole la 70 mmHg (efect max. la 35 mmHg) determina vasoconstrictie in teritoriul respectiv, cu dirijarea sangelui spre zonele bine ventilate (se presupune ca are loc eliberarea din celulele alveolare a unor substante vasoconstrictoare). Scaderea pH, cresterea CO2 determina deasemenea vasoconstrictie.
Efectul gravitatiei
In ortostatism distanta de la varf la baza este de 30 cm, deci diferenta de presiune de 23 mm Hg - presiunea deasupra inimii scade la varf cu 15 mmHg, iar dedesuptul inimii creste la baza cu 8 mmHg (deci in loc de 25 mmHg avem 10 la varf si 33 la baza-25+8). Capilarele sunt destinse de presiunea sangelui, dar colabate de presiunea din alvole. In plamani se poate vorbi de 3 zone:
Zona I - presiunea din alveole o depaseste pe cea din capilare si astfel avem o lipsa de debit. Se poate intalni in ortostatism la varf, cand se respira la presiuni crescute sau cand TA e scazuta.
Zona II - debit intermitent, se intalneste la varf in ortostatism deoarece in sistola TA = 25-15 = 10 mm Hg, in diastola 8-15 = -7, deci mai mica decat presiunea alveolara, astfel incat sangele circula doar in sistola.
Zona III - debit continuu, apare in intreg plamanul in clinostatism sau in efort fizic
Schimbul de gaze la nivelul membranei alveolo-capilare
In timpul ventilatiei alveolare (a celor 60.000 de acini) viteza curentului de aer - a convectiei aerului - scade pe masura divizarii arborelui bronsic, suprafata de sectiune creste de la 2,5cm2 - suprafata traheei la 180cm2 - suprafata de sectiune la nivelul bronhiolelor terminale. Ca urmare, la nivelul acinilor, deci a bronhiolelor respiratorii, canalelor si sacilor alveolari, viteza de convectie a aerului devine mai mica decat difuziunea gazelor din compozitia sa (proces pasiv prin care gazul se deplaseaza de la concentratie mai mare la concentratie mai mica).
Intre sange si aerul alveolar se interpune mambrana alveolar-capilara formata din: epiteliul alveolar, membrana bazala a alveolei si a capilarului, endoteliul capilar care vine in contact cu membrane hematiei. Traversarea acestei membrane de gazele respiratorii se face prin difuziune. Gazele fiind liposolubile traverseaza cu usurinta membranele celulare. Difuziunea prin lichidul celular si extracelular presupune dizolvarea gazelor in acest lichid.
Conform legii lui Henry cantitatea de gaz dizolvata e direct proportionala cu presiunea partiala a gazului si cu solubilitatea acestui gaz. Coeficientul de solubilitate al CO2 este de 0,57 al O2 este de 0,024, deci CO2 este de 24 ori mai solubil decat O2 .
Viteza de difuziune a celor 2 gaze prin membrana alveolo-capilara este direct poportionala cu diferenta de presiune ΔP, cu suprafata membranei (A) si cu solubilitatea gazului (S) si invers proportionala cu distanta (d) deci grosimea membranei si cu greutatea moleculara a gazului. Cele mentionate pot fi exprimate prin urmatoarea formula :
Viteza de difuziune = ΔP.A.S/d√Mw
Factorul ce depinde de gaz este s/√Mw si reprezinta coeficientul de difuziune care este de 1 pentru O2 si de 20,3 pentru CO2 , deci CO2 este de 20 de ori mai difuzabil decat O2 .
Dozarea gazelor din aerul alveolar se exprima in volume, difuziunea se face in functie de presiunile lor partiale. Pentru a transforma volumele in presiuni partiale se tine seama de legea lui Dalton conform careia presiunea partiala a unui gaz dintr-un amestec este egala cu presiunea pe care ar avea gazul daca ar ocupa singur volumul amestecului. Tinand seama de aceasta lege calculul presiunii partiale se face astfel:
In aerul alveolar concentratia O2 = 13,6%, al CO2 5,3%.
Presiunea totala a gazelor din aerul alveolar este presiunea atmosferica din care se scade presiunea vaporilor de apa, aerul alveolar fiind saturat cu vapori de apa si la temperatura de 37 °C si are o presiune de 47mm Hg, deci : 760-47=713.
In cazul O2 :
|
100vol | |||||
|
X.... | |||||
|
X |
x |
104 mm Hg |
|||
In cazul CO2 :
|
100vol | |||||
|
X ... | |||||
|
X |
|
x |
40 mm Hg |
||
In sangele venos din capilarele pulmonare
P O2 = 40 mm Hg, P CO2 = 46 mm Hg
In concluzie O2 trece din alveola (P = 100mmHg) in sange (P = 40 mmHg), iar CO2 din sange (P=46 mmHg) in alveola (P=40 mmHg).
Suprafata de schimb, este suprafata unde alveolele ventilate vin in contact cu capilarele perfuzate. Ea are o valoare de cca 70 m2 . Cand se reduce la 1 -¼ schimbul de gaze e afectat si in repaos (fenomen intalnit in emfizem, cand se distrug septurile dintre alveole, sau in ocluzii ale arterei pulmonare).
Grosimea membranei alveolo-capilare: normal distanta este de 0,2 - 0,5µm. Ea creste in edem pulmonar, in fibroza. Cand grosimea membranei creste de 2-3 ori, schimbul de gaze e afectat, mai ales cel de O2 , iar fenomenul poarta denumirea de "bloc alveolo-capilar".
Aprecierea difuziunii celor doua gaze se face determinand capacitatea de difuziune, care este volumul de gaz ce difuzeaza pe minut pentru o diferenta de presiune de 1mm Hg. Ea este:
pentru O2 in repaos de 21 ml si creste in efort la 65ml (prin dilatarea capilarelor si deschidere de capilare noi deci prin cresterea suprafetei).
pentru CO2 se deduce prin calcul tinand seama de coeficientul de difuziune de 20 de ori mai mare - deci 400 ml in repaos, 1200-1300ml in efort (determinarea e dificila din cauza vitezei foarte mari de difuziune a CO2 ).
In urma difuziunii celor 2 gaze prin membrana alveolo-capilara, presiunea lor partiala din sange devine egala cu cea din aerul alveolar. Egalarea presiunilor, pentru CO2 se face instantaneu, pentru O2 - in 0,3 secunde, ori capilarul e strabatut de sange in repaos in 0,7 sec.
P O2 = 104 mm Hg -alveola
P O2 = 40 P O2 = 104 - capilar
![]()
![]() 100
100
![]()
![]()
![]() 0,75
0,75
40 0,3
Aceasta difuziune rapida a O2 face posibila oxigenarea sangelui in efort cand capilarul e strabatut de sange in 0,3 sec.
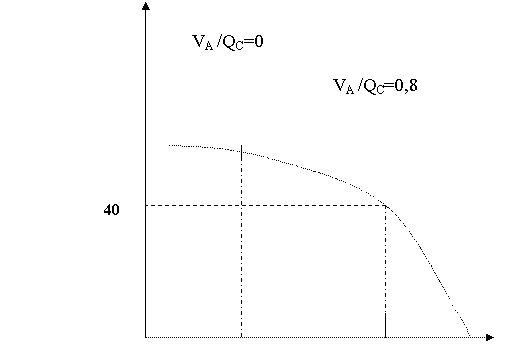
Oxigenarea sangelui, deci mentinerea unei presiuni constante de O2 si CO2 in alveole depinde de raportul dintre ventilatie si perfuzie (VA /QC ). Normal acest raport este de 4/5 = 0,8, dar situatie ideala nu exista. In ortostatism la baza plamanului creste perfuzia deci raportul scade, la varful plamanului ventilatia e mai mare deci raportul creste. In medie, in majoritatea alveolelor se mentine la 0,8.
Efectele modificarii raportului - sa luam situatia extremelor:
P CO2
VA /QC≥
P0
 100 PO
100 PO
In cazul ventilatie 0, perfuzie normala aerul din alveole se va apropia de presiunile partiale ale gazelor din sangele venos. Aceasta situatie se compara cu un sunt arterio-venos si duce la hipoxemie (scaderea PO2 din sange).
In cazul ventilatiei normale, dar a perfuziei 0 compozitia aerului alveolar se apropie de cea a aerului inspirat, nu au loc schimburi, e situatia spatiului mort functional. Si acest caz se insoteste de hipoxemie.
Cazurile cele mai frecvente de scaderi ale oxigenarii sangelui nu sunt hipoventilatia, nici tulburarile de difuziune, nici alternarea debitului sanguine ci o neuniformitate a raportului VA /QC.
Cauzele ventilatiei neuniforme:
cresterea rezistentei la nivelul cailor aeriene (spasm bronsic);
scaderea compliantei pulmonare;
Cauzele perfuziei neuniforme:
actiunea gravitatiei;
embolii, tromboze in circulatia pulmonara;
ateroame care ocluzioneaza arterele;
obliterari fibroase ale arterelor;
hipotensiune severa ce duce la inchiderea unor vase pulmonare.
SSSSchimbul la nivelul capilarelor
Presiunea sangelui este de 7 mm Hg, presiunea interstitiala e de 8 mm Hg, iar presiunea coloid osmotica din interstitiu 14 mm Hg. Aceste 3 forte sunt profiltrare si insumate dau 29 mm Hg.
Presiunea coloid osmotica a sangelui are valoarea de 28 mm Hg si reprezinta forta proreabsorbtie.
In concluzie presiunea efectiva de filtrare este de 1mm Hg.
Lichidul filtrat este preluat rapid de limfaticele plamanul, fiind organul cu circulatia limfatica cea mai dezvoltata.
Cand presiunea sangelui in capilare creste, se intensifica filtrarea ce duce la acumulare de lichid in interstitiu, apoi in alveole, adica la instalarea edemului pulmonar. Prevenirea edemului se face prin intensificarea circulatiei limfatice:
daca presiunea crescuta se mentine, circulatia limfatica se dezvolta;
cand presiunea creste la 35-45 mm Hg se instaleaza edemul.
TRANSPORTUL GAZELOR RESPIRATORII
Oxigenul (O2) este transportat :
dizolvat in sange;
fixat de hemoglobina;
Oxigenul dizolvat: dizolvarea este direct proportionala cu presiunea oxigenului. La PO2 de 100 mm Hg (alveola, sange arterial) se dizolva 0,3 ml O2%. La PO2 de 95 mm Hg (din cauza impurificarii sangelui arterial cu sange venos) se dizolva 0,29 %.
La nivelul tesuturilor, la PO2 de 40 mmHg, in sange raman 0,12 ml O2%, deci se consuma 0,17 ml O2%, o cantitate foarte mica. Forma dizolvata prezinta importanta deoarece transferal prin membrane (alveolo-capilare, capilare) se face sub aceasta forma.
Oxigenul fixat pe hemoglobina (Hb): Hb este o componenta formata dintr-o parte proteica - globina (96%) si o parte neproteica - hem (4%). Hemul este un inel tetrapirolic si are in centrul lui Fe++ . Fiecare Fe++ fixeaza o molecula de oxigen. Cum Hb este un tetramer, fiecare monomer fiind format dintr-un lant al globinei (2α, 2β) si un inel tetrapirolic cu Fe++ , o molecula de Hb fixeaza 4 molecule de O2 (proces de oxigenare nu de oxidare).
Structura quaternara a Hb prezinta alosterie, adica se modifica dupa fixarea unor liganti cum ar fi O2, deoarece legaturile dintre lanturile α1- β1 si α2- β2 sunt fixe, in schimb intre α1- β2 si α2- β1, sunt mobile si intervin in schimbarile de configuratie.
Din punct de vedere al configuratiei spatiale, Hb se gaseste sub 2 forme:
Forma relaxata sau oxigenata cu lanturile β mai apropiate, favorizand fixarea oxigenului.
Forma tensionata sau redusa in care fixarea oxigenului e slaba.
Fixarea sau eliberarea oxigenului pe sau de pe cei patru atomi de Fe bivalent ai Hb se face succesiv. Fixarea pe primul Fe, prin efect alosteric creste afinitatea pentru O2 a Fe II, a Fe II o creste pentru Fe III etc, a.i. afinitatea Fe IV pentru O2 creste cu 125 de ori fata de a Fe I. La o PO2 de 100 mmHg, reactia are loc in 0,01 secunde.
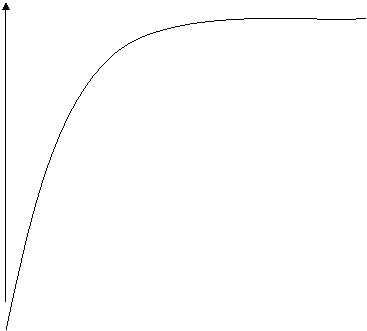
Relatia dintre PO2 si formarea O2Hb (saturarea Hb cu O2) se inscrie sub forma unei curbe stabilite de Barcroft, curba cu forma literei S.
|
. - - - - - - - - - 20 40 60 80 100 P oxigen |
100 mm Hg - 97,5 80 mm Hg - 94,5 60 mm Hg - 89 40 mm Hg - 75 20 mm Hg - 35 10 mm Hg - 13,5 |
Acest aspect al curbei de disociere a O2Hb are doua avantaje:
1. La PO2 60-100 mm Hg saturarea cu O2 este peste 90%; deci doar cu 7 mm Hg mai mica decat saturatia normala. In cazul presiunilor mai mari (ex. 500 mm Hg) saturatia nu poate fi mai mare decat 100%.
2. La nivelul tesuturilor pierderea O2 este rapida si cand PO2 scade la 15-20 mm Hg (efort fizic), desprinderea O2 se face si mai repede.
In acest fel, Hb mentine PO2 la nivelul tesuturilor relativ constanta, deci o stabilizeaza.
Hb saturata 100% cu oxigen fixeaza per gram 1,34 ml O2, astfel la nivelul plamanului se fixeaza pe cele 15 gr Hb - cca 20 ml O2. La nivelul tesuturilor, la PO2 = 40 mmHg si saturatia Hb de 75%, raman in sange cca 15 vol O2%. Deci din fiecare 100 ml sange se consuma 5 ml O2 (25%).
Acesta este coeficientul de utilizare al O2. El creste in efortul fizic, deoarece disocierea O2Hb este favorizata de:
cresterea concentratiei CO2 (efect Bohr);
cresterea concentratiei ionilor de H;
cresterea temperaturii;
Tinand cont ca debitul sanguin este de 5,5 l/min, consumul de O2, in repaos este de 250 ml/min. Rezervele de O2 ale organismului sunt reprezentate de:
1000 ml O2 in circulatie;
900 ml O2 in plamani;
150-200 ml O2 in mioglobina.
Ele pot asigura necesitatile organismului O2 in caz de apnee, pentru maximum 4 minute, dup care PO2 din alveole ajunge la 30 mm Hg iar in sangele arterial la 20 mm Hg.
Transportul CO2 se face sub forma de:
CO2 dizolvat;
HCO3 - ;
Carbamati.
CO2 se formeaza in celule ca urmare a proceselor de oxidare. PCO2 in tesuturi creste la 50-70 mm Hg. El difuzeaza in sange prin membrane celulara si prin peretele capilar (in sangele arterial PCO2 = 40 mmHg, in sangele venos PCO2 = 46 mm Hg). In plasma raman 8% din care 5% se leaga de gruparile aminice ale proteinelor si 3% se dizolva. 92% trec in hematii, 82% formeaza HCO3 - (fenomen de membrana Hamburger), 10% se fixeaza pe gruparile aminice ale globinei (mai ales din Hb deoxigenata) formand carbamatii de Hb. (la PCO2 = 46mm Hg se adauga cca 4ml CO2 %).
La nivel pulmonar reactiile se inverseaza, eliminandu-se CO2. Din cei cca 5l, ai debitului sanguin/min, se elimina 200 ml CO2/min. La nivelul tesuturilor CO2 dizloca O2 de pe Hb (efect Bohr). La nivel pulmonar O2 dizloca CO2 de pe Hb (efect Haldane) deoarece la PO2 =100 mm Hg, Hb fixeaza O2 , ca urmare:
O2Hb fixeaza mai putin CO2 sub forma de carbamati, deci se elimina CO2 din carbamati;
O2Hb este un acid mai puternic de 70 de ori decat Hb, deci se disociaza mai mult, cedeaza mai mult H+, care unindu-se cu HCO3- formeaza H2CO3; acesta se descompune in CO2 si H2O, iar CO2 se elimina.
Conform acestui efect la nivel pulmonar O2 dizloca CO2 , la nivel tisular, scaderea O2 favorizeaza fixarea CO2 .
REGLAREA RESPIRATIEI
Reglarea respiratiei consta in adaptarea ventilatiei pulmonare la necesitatile metabolice ale organismului, cu mentinerea in sangele arterial a presiunii O2 la 104 mm Hg si a CO2 la 40 mm Hg.
Respiratia de repaos se datoreste unor impulsuri nervoase din trunchiul cerebral spre neuronii motori ai muschilor ce determina inspiratia. Ca dovada, sectionarea MS sub bulb duce la oprirea respiratiei, iar sectionarea deasupra puntii nu influenteaza respiratia de baza. Centrii din trunchiul cerebral se gasesc in formatiunea reticulata bulbo-pontina.
Neuronii din bulb sunt grupati in:
1. Grupul dorsal se afla la nivelul Nc. tractului solitar. Aici se termina fibrele sensitive ale nervilor glosofaringian si vag. Din neuronii acestui grup pleaca impulsuri spre Nc. nervului frenic, si de asemenea spre grupul ventral. Neuronii acestui grup emit spontan si ritmic impulsuri (chiar daca se sectioneaza toate aferentele) ca si cum ar avea automatism. PA elaborat e slab la inceput, creste in intensitate, in panta, timp de cca 2 secunde, apoi se opreste pentru alte 3 secunde si ciclul reincepe de 12-15 ori pe minut. Aceasta crestere treptata a intensitatii impulsului face ca inspiratia sa nu aiba caracter brusc, spastic.
2. Grupul ventral are:
- o componenta craniala (in N. ambiguu);
- o componenta caudala (in Nc.retroambiguu.)
Neuronii acestui grup nu prezinta automatism. Ei primesc impulsuri de la grupul dorsal si sunt in legatura atat cu neuronii inspiratori cat si cu cei expiratori, mai ales cu cei care intervin in expiratia fortata (ai muschilor abdominali).
De remarcat ca la nivelul neuronilor motorii din maduva exista o inervatie reciproca in sensul ca excitarea neuronilor spre muschii inspiratori ii inhiba pe cei spre muscii expiratori si invers.
Centrii din punte
Sectionarea SN la mijlocul puntii duce la oprirea respiratiei in inspiratie (apneuzis). Se datoereaza faptului ca in punte se gasesc 2 centrii:
1.Centrul pneumotaxic actioneaza asupra neuronilor din grupul dorsal, scurtand inspiratia, ceea ce duce la cresterea frecventei respiratorii (un stimul duce la 30-40 respiratii/min).
2.Centrul apneustic, contestat de unii - stimuleaza neuronii inspiratori. Din aceasta cauza, in cazul sectiunii descrise, neuronii inspiratori sunt lipsiti de actiunea pneumotaxicului si sub influenta n. apneustic inspiratia se prelungeste si se opreste in inspiratie. Un efect asemanator centrului pneumotaxic il au nervii vagi, stimulati de distensia bronsica. Acest reflex a fost numit al lui Hering-Brener.
Deci intermitenta de descarcare a neuronilor inspiratori se datoreaza inhibarii lor de n. pneumotaxic si de n. vag. La om, nervii vagi intervin cand volumul respirator current (VRC) ajunge la 1500 ml, deci acest reflex ar avea mai mult un rol de protectie impotriva unei distensii pulmonare prea mari. Daca se face o sectiune sub punte respiratiile raman ritmice (indiferent de sectiunea nn vagi) dar sunt neregulate ca amplitudine si sacadate, deci aparent, centrii pontini ar determina egalitatea descarcarilor ritmice bulbare si trecerea lina de la inspiratie la expiratie.
Reglarea activitatii centrilor respiratori, adica adaptarea la necesitatile metabolice, se face prin factori metabolici: cresterea pCO2 , scaderea pO2 , scaderea pH, deci printr-un control chimic. Acest control se face prin chemoreceptori - terminatii nervoase sau celule care sesizeaza modificarile amintite, adaptand debitul respirator la necesitatile metabolice.
A. Factorul principal este pCO2 , (hormon respirator).
Chemoreceptorii sunt:
1.Chemoreceptori centrali;
2.Chemoreceptori periferici.
1.Chemoreceptorii centrali se gasesc pe partea ventrala a bulbului ( la 200-400 µm de suprafata ventrala), separati de centrul respirator ventral. Ei sunt stimulati de cresterea concentratiei ionilor de H din mediu (LCR sau lichid interstitial), dar si de cresterea ionilor de H din sange, deoarece acesta nu traverseaza bariera hematoencefalica. Cresterea concentratiei de CO2 din sange ii stimuleaza indirect. El traverseaza bariera hematoencefalica cu usurinta (e usor difuzabil) si se uneste cu H2O dand H2CO3 . Acesta disociaza in HCO3- si H+ , iar H+ stimuleaza chemoreceptorii. Stimulul e transmis grupului dorsal intensificand inspiratia.
2.Chemoreceptorii periferici se gasesc in corpusculii carotidieni (cate unul la bifurcatia carotidei primitive) si corpusculii aortici ( doi sau mai multi in apropierea arcului aortic). Prima descriere a fost facuta de Heymans (1938) care a primit cu aceasta ocazie premiul Nobel. Mai bine studiati sunt cei carotidieni - ceva mai mari si mai usor accesibili. Au o greutate de 2 mg fiecare si sunt foarte puternic irigati - cel mai mare debit din organism - de cca 40 de ori mai mare ca al creierului si au un metabolism foarte crescut (de 2-3 ori mai mare ca al creierului) dar debitul fiind foarte mare PO2, PCO2 si pH-ul in capilare sunt la fel ca in sangele arterial. Celulele chemosensibile sunt celule de tipI, cu diametru de 10µm, sferice dispuse in insule, comunica intre ele prin jonctiuni GAP (de origine neuroectodermica) - prezinta caracteristici comune cu neuronii si cu celulele cromofine adrenale. Astfel:
unele sunt inervate de fibre pregangl. simpatice;
au canale ionice dependente de voltaj;
depolarizarea duce la aparitia PA;
au numeroase vezicule intracelulare ce contin o varietate de mediatori.
Stimularea acestor celule determina eliberarea de acetilcolina, NA, subst.P, si met-encefalina. Eliberarea acestor substante controleaza activitatea terminatiilor nervoase. In jurul acestor celule sunt terminatiile sensitive ale nervului sinusului carotidian (rr ale n.IX). Mediatorii chimici eliberati de celulele chemoreceptoare declanseaza PA care ajunge in grupul neuronal dorsal (component al Nc.Tractului solitar). Si terminatiile nervoase din corpusculi poseda vezicule deci sinapsa cu celulele poate fi bidirectionala (functia nu e cunoscuta).
In jurul insulelor de celule tipI sunt celule de tipII - celule de sustinere (asemenea celulelor gliale). In stransa vecinatate exista o retea de densa de capilare fenestrate.
Corpusculul carotidian este inervat simpatic si parasimpatic - terminatiile preganglionare fac sinapsa cu celulele de tipI, dar si cu vasele sanguine, simpaticul producand vasoconstrictie - scade debitul sanguin si astfel PO2 , stimuland receptorii.
Chemoreceptorii descrisi sunt stimulati in principal de scaderea PO2 ,dar si de cresterea PCO2 si a concentratiei ionilor de H. Toti acesti factori actioneaza prin inhibarea canalelor de K+ .Ca urmare are loc depolarizarea membranei, urmata de patrunderea Ca++ in celula si de eliberarea mediatorului chimic (cei 3 actioneaza asupra canalelor de K pe cai diferite). Stimulii de la chemoreceptori sunt transmisi Nc.dorsal prin nn glosofaringieni IX (din corpusculul carotidian) si prin n.vag X (de la corpusculii aortici).
Dupa denervarea corpusculului carotidian raspunsurile la scaderea PO2 si la cresterea H+ , sunt abolite, iar la cresterea PCO2 e diminuat doar cu 20%, ceea ce demonstreaza importanta chemoreceptorilor centrali in cazul cresterii PCO2 .
Viteza raspunsului e determinata de chemoreceptorii periferici, intensitatea de cei centrali.
Raspunsul la cresterea PCO2 :
intensificarea activitatii centrilor respiratorii;
in hipercapnie (CO2 in alveole este peste 7%) este deprimata activitatea intregului SNC (narcoza cu CO2), ajungandu-se la confuzie si coma.
Raspunsul la lipsa O2 : La PO2 peste 60 mmHg e slab. Sub 60 mm Hg se intensifica ventilatia (deoarece Hb fixeaza H+, cresterea ventilatiei scade CO2, ambele reduc ventilatia). La sub 60 mm Hg efectul stimulator il depaseste pe cel inhibitor.
Asupra centrilor respiratorii actioneaza si influente de la alti centrii nervosi.
Relatii intercentrale:
deglutitia, voma, opresc respiratia;
Aferentele de la hipotalamus, sistemul limbic (durerea, emotiile) influenteaza activitatea centrilor respiratorii
Exista si aferente spre centrii respiratori de la neocortex - respiratia poate fi oprita sau intensificata voluntar. Controlul voluntar intervine intervine in innot, vorbire, cantat.
Controlul automat si cel voluntar sunt separate uneori, cel automat poate fi dereglat fara a fi cel voluntar, deci se respira constient - situatia clinica - in compresii ale bulbului, poliomielita bulbara sau sectiuni ale cailor controlului automat (respiratia devine constienta*blestemul Ondinei*).
Asupra centrilor respiratori actioneaza si aferente de la proprioceptorii din muschi si articulatii.
In efortul fizic, cu tot consumul crescut al O2 (250-4000ml/min) si eliminarea unei cantitati mari de CO2 (200-8000 ml/min), PO2 si PCO2 in sangele arterial raman constante. Ventilatia pulmonara se intensifica de la inceputul efortului fizic, nu prin factori chimici, ci prin factori nervosi. La inceputul efortului, concomitent cu transmitera impulsurilor de la centrii motori corticali spre musculatura activa, sunt transmisi stimuli si spre centri respiratori, intensificand ventilatia (la fel ca actiunea asupra centrului cardio-vasomotor). Deasemenea stimulii de la proprioceptori intensifica ventilatia. Factorii chimici intervin in cazurile in care activarea centrilor respiratorii pe cale nervoasa nu corespunde exact cerintelor metabolice si ajusteaza intensificarea ventilatiei la necesitati. In cazul efortului fizic intervin si:
cresterea debitului (30x);
cresterea capacitatii de difuziune (de 3x);
cresterea coeficientului de utilizarea oxigenului (de 3 ori).
Toate acestea permit cresterea ratei metabolice de 100 de ori asigurand in acest fel intensificarea activitatilor metabolica.
Hipoxia reprezinta scaderea PO2 la nivelul tesuturilor si are multe cauze:
1.Hipoxie hipoxica - consecinta scaderii PO2 alveolar;
2.Hipoxie stagnanta - consecinta scaderii vitezei de circulatie a sangelui intr-o anumita regiune.;
3.Hipoxie anemica - consecinta scaderii Hb;
4.Hipoxie histotoxica - consecinta incapacitatii celulare de a utiliza O2 ( sub actiunea cianurii sau altor toxice).
In cazul hipoxiei stagnante e afectat tesutul slab irigat. In celelalte cazuri, in primul rand este afectat S.N. Cand PO2 alveolar scade sub 20 mm Hg se pierde constienta, se instaleaza coma, iar in 4-5 minute survine moartea. O hipoxie mai putin severa duce la tulburari psihice:
- stare de excitatie;
- dezorientare in timp si spatiu;
- incapacitatea unei activitati de gandire, memorare;
- cefalee,
- tulburari digestive;
- tahicardie;
- hipotensiune;
- scaderea fortei fizice.
In hipoxia severa se poate ajunge la edem cerebral si edem pulmonar.
Hipoxia hipoxica se instaleaza fie in afectiuni pulmonare fie in timpul ascensionarilor cu avionul sau pe jos.In cazul ascensionarilor - deoarece presiunea atmosferica scade si odata cu ea si PO2 din aer, a carei concentratie ramane tot cca 21% astfel:
|
p. barom =760 |
PO2 =159 |
PO2 alv.=104 |
Sat Hb=97% |
|
Efectele hipoxiei la cei neaclimatizati apare la 4.000 m si se traduc prin oboseala fizica si psihica, dezorientare, tulburari digestive, cefalee, uneori euforie. La peste 4.500 m apar convulsii. La peste 6.000 m se instaleaza coma. Daca se respira O2 pur, acesta dizloca N2 (4/5 din aerul alveolar) astfel incat la 10.000 m PO2 alveolar in loc de 18 mm Hg creste la 139 mm Hg. Dupa ce s-a ajuns la altitudine, raul de altitudine (insomnie, cefalee, tulburari digestive) se instaleaza in 8-24 de ore si dureaza 4 - 8 zile, simptome care se amelioreaza prin reducerea alcalozei (hiperventilatie) si evitarea in primele 3 zile a efortului fizic.
Aclimatizarea se face prin:
intensificarea ventilatiei (creste sensibilitatea centrilor respiratori la scaderea PO2 prin scaderea pH din LCR);
cresterea numarului hematiilor (secretia de eritropoietina - efect in 3 zile);
intensificarea consumului de oxigen la nivelul tesuturilor (creste nr.mitocondriilor, numarul enzimelor respiratorii citocromoxidaza)
In Anzi si Himalaya exista locuinte umane permanente la 5.500 m. Nativii din aceste regiuni au policitemie si PO2 alveolar scazut, in rest fiind normali.
Presiunea oxigenului crescuta: Sunt situatii care necesita administrare de O2 (interventii pentru afectiuni congenitale cardiace, intoxicatii cu CO, cianuri), in tratarea gangrenei gazoase. Oxigenul la presiune crescuta este toxic pentru tesuturi deoarece formeaza anioni de superoxid si apa oxigenata, deci formarea de radicali liberi cu efect de oxidare care produc leziuni pulmonare (inhiba activitatea macrofagelor, reduce cantitatea de surfactant) si leziuni tisulare. In caz de administrare se recomanda - sub 5 ore si sub o presiune de 3 atm.
Cresterea PCO2 (hipercapnie) - apare mai ales in caz de hipoventilatie si duce la intensificarea respiratiei, deprimarea SNC, deci a centrilor respiratorii - coma - moarte. Cand CO2 din alveole ajunge la 7% se ajunge la inhibarea SNC (narcoza prin CO2 ).
Hipocapnia (scaderea CO2 ) - apare ca urmare a hiperventilatiei cronice intalnite la nevrotici si duce la scaderea debitului circulator la nivelul creierului cu 30% si are ca urmare: aparitia paresteziilor, alcaloza respiratorie (pH=7,5-7,6), scaderea Ca2+ plasmatic, care duce la tetanie si hiperexcitabilitate.
Efectele cresterii presiunii barometrice (la scafandri): pentru fiecare 10 m, presiunea din apa creste cu 1 atm, de aceea in chesoane (camera inchisa presurizata), presiunea aerului inspirat creste la valoarea presiunii barometrice pentru a egala presiunea ce se exercita asupra cutiei toracice si abdomenului. Se foloseste amestec de O2 (20%) si N2 (80%). La presiunea de 4-5 atm, azotul poate produce euforie (betia adancurilor) asemanatoare celei de dupa consumul de alcool, se reduc functiile intelectuale si precizia miscarilor fine. Se prefera utilizarea unui amestec de O2 si He pentru a evita narcoza cu N2 . Probleme apar la decompresie, deoarece la presiuni crescute N2 (liposolubil) patrunde in cantitate mare in tesuturi in care se dizolva si, in caz de decompresie brusca, se elibereaza sub forma de bule, producand leziuni tisulare si astupari ale vaselor mici (embolii) ce duc la ischemii tisulare. Apar dureri in muschi si articulatii, pot aparea si simptome nervoase (5-10%) - paralizii, colaps cu pierderea constiintei. In 2% apare socul prin obstructia masiva a capilarelor pulmonare, edem pulmonar si moarte. Din aceasta cauza se practica decompresia treptata cu o durata de cca 3 ore ( la cei ce stau sub apa mai mult de o ora).
Copyright © 2025 - Toate drepturile rezervate