|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
STRUCTURA ARN
1. Structura primara a ARN
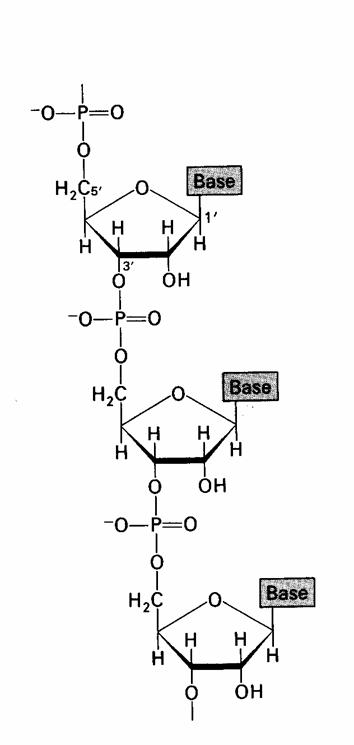
Ca si in cazul ADN, structura primara a ARN este determinata de succesiunea nucleotidelor in catena. Desi apropiata de structura ADN, exista doua deosebiri:
prezenta gruparii hidroxil in pozitia 2' a ribozei;
prezenta, ca baza pirimidinica, a uracilului in locul timinei.
|
|
|
ARN. Structura primara |
Prezenta gruparii hidroxil impiedica - partial - asocierea ARN sub forma bicatenara, marind flexibilitatea catenei, astfel ca, diversitatea conformationala a ARN, ca si reactivitatea acestuia sunt mai accentuate comparativ cu ADN.
Determinarea structurii primare a ARN, realizata pentru prima data prin hidroliza alcalina a condus la un rezultat oarecum neasteptat: au fost identificati atat 3'- cat si 2'-ribonucleozidfosfati.
O structura primara similara cu ADN, in care bazele
se succed in ordinea
Prezenta, intre
produsii de hidroliza neenzimatica, a 2'-monofosfatilor
sugera insa si existenta unor legaturi 2'-
Interesant este faptul ca, din punct de vedere termodinamic,
legaturile 3'-
2. Structura secundara a ARN
Desi prezenta gruparilor hidroxil
Se pare ca aceeasi grupare
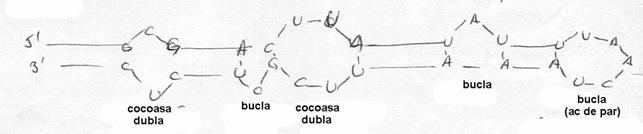
Aceste secvente dublu-catenare contin nu numai perechi normale A:U si G:C, dar si numeroase perechi G:U, mai putin stabile energetic.
Moleculele ARN mai lungi, ca de exemplu fragmentul 16S al ARNr contine probabil mai multe secvente dublu catenare, separate prin bucle (lupinguri) monocatenare.
|
|
|
Structura ARN de tip secundar |
Mentinerea structurii depinde de mai multi factori: concentratia de sare, pH, temperatura, ioni (Mg2+, ioni poliaminici). La temperaturi suficient de ridicate ARN este "total" denaturat, disparand secventele dublu-catenare.
2.1. Secventele de tip "cocoasa", ac de par si buclat
Lungimea mai redusa a ARN a permis realizarea unor studii teoretice privind constantele de asociere a oricaror perechi de baze situate in diverse vecinatati, inclusiv in cazul existentei unor structuri deosebite de tip "cocoasa", "ac de par" sau "buclat". Astfel, formarea unei structuri ac de par ce contine 5-6 baze, nelegate, necesita 25 kJ/mol, o cocoasa aprox. 9 kJ/mol, iar o bucla ~13 kJ/mol. Folosindu-se aceste date, ca si alte determinari, a putut fi determinata structura unor ARN, de ex. a unui fragment de 55 nucleotide din ARN al virusului R17.
|
|
|
Secvente de tip "cocoasa", ac de par si buclat |
2.2. Structura ARN de transfer (transport)
Datorita catenei mai scurte, ARNt reprezinta una dintre cele mai studiate molecule de acid nucleic, atat in stare nativa (a fost determinata secventa primara a cateva sute de ARNt), cat si a unor forme obtinute prin sinteza.
Toate moleculele studiate
contin contin o secventa identica -CCA la capatul
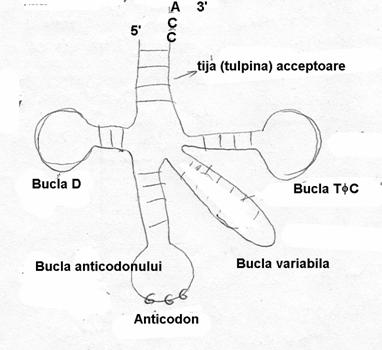
Desi numarul de resturi poate varia intre 74 - 95 nucleotide, totusi ele adopta o conformatie secundara similara in forma de trifoi in care patru sau cinci regiuni dublu catenare (stem= tulpina, tija) sunt separate prin bucle monocatenare.
|
|
|
Structura ARNt |
Toate moleculele ARNt au acelasi numar de baze in tija acceptoare (7pb) si in bucla anticodonului si, respectiv,TΨC ambele au cate 7 baze in bucla si 5 pb in tija.
Lungimea variabila a diferitelor molecule de ARN este determinata de existenta unor regiuni variabile: bucla D si bucla "variabila".
A fost determinata si structura tertiara a unor ARNt; ca urmare a "impachetarii", apar legaturi de hidrogen intre baze situate la distante relativ mari in structura secundara. Rezulta o conformatie in forma de L, fiecare latura fiind reprezentata de secvente dublu catenare; conectarea este realizata prin buclele D si TΨC. Bucla anticodonului se gaseste la capatul bratului lung, iar zona acceptoare la capatul bratului scurt. Prin difractie de raze X s-a constatat ca aceeasi structura L se pastreaza si in cazul asocierii ARNt cu proteine.
Sunt prezente numeroase imperecheri neuzuale (Hoogston si
altele), legaturi 2'-
2.3. ARN antisens
ARN antisens este o molecula scurta, lipsita de capacitatea de codare, dar care se caracterizeaza printr-un grad inalt de complementaritate cu alta molecula de ARN, cu care poate forma hibrizi. ARN antisens poate astfel functiona ca un modulator (represor) al expresiei ARN "tinta". Astfel de molecule au fost obtinute prin sinteza, dar au fost detectate si in celule de procariote, fiind probabil implicate in replicarea ADN plasmidic, in transcrierea ADN bacterian, ca si in translatia ADN mesager din bacterii si bacterofagi.
3. Duplexuri ADN-ARN
In afara de asocierea cu molecule de acelasi tip (duplexuri ADN-ADN si respectiv ARN-ARN) foarte importante sunt si asocierile ADN-ARN. Acestea se pot forma in mai multe cazuri:
transcrierea ADN in ARN mesager complementar;
sinteza unei catena ADN prin retrotranscriptia din ARN viral;
asocierea cu fragmentele ARN "primer" la sinteza fragmentelor Okazaki - (in replicarea ADN),
fiind posibile si alte cazuri.
Hibrizi similari se pot forma si in vitro prin asocierea a doua catene ce comtin baze complementare, de ex. cele formate de catenele de tip ARN poli (rA) poli (dT) si poli (rI)poli(dC). Primul are o conformatie de tip A-ARN (cu 11 pb/spira), iar cel de-al doilea are o conformatie de tip A'-ARN.
Un decamer autocomplementar r(GCG)d(TATACGC) formeaza un duplex de tip Watson - Crick (corespunzator A-ADN sau A-ARN).
Acesti hibrizi sunt mai stabili la procesele de denaturare comparativ cu duplexul ADN-ADN.
Stabilitatea deosebita a duplexului ADN-ARN a condus la ideea construirii unor oligomeri ADN antisens care sa formeze hibrizi si sa inhibe ARNm.
Desi astfel de secvente ADN (ca de altfel si ARN) antisens au fost sintetizate, au fost intampinate greutati deosebite privind traversarea membranelor celulare sau atacul enzimelor de hidroliza. Pentru marirea rezistentei, restul fosforic din legatura fosfodiesterica a fost inlocuit cu resturi fosfortionate, fosforditionate sau metilfosforice, dar heteroduplexurile rezultate au constante de asociere mult mai reduse comparativ cu duplexurile ARN-ADN. Studiile continua, utilizarea de oligonucleotide ARN sau ADN antisens fiind una dintre marile sperante ale terapiei genice.
4. Triplexuri de acizi nucleici
In afara de duplexurile prezentate anterior, sunt posibile si asocieri realizate intre trei catene de acizi nucleici. Primul triplex de acest tip a fost sintetizat in 1957 cand s-a obtinut un complex 2 poli(rU).poli(rA), ulterior fiind obtinute si alte triplexuri; ca de exemplu: 2 poli(rC).poli(rG), una dintre catenele rC fiind protonata.
S-au obtinut si structuri mai complexe, prin asocierea intre carene ribozice si deoxiribozice, ex. cele formate prin asocierea poli(rU-rC) sau poli(dT-dC) cu duplexul poli (dT-dC).poli(dG-dA); ca si in celelalte cazuri, triplexul este stabil la pH acid in prezenta de MgCl2. Duplexul are o structura de tip A', cea de-a treia catena fiind situata in santul major relativ adanc al duplexului. De asemenea, se observa ca cea de-a treia catena se leaga in special prin legaturi de tip Hoogsteen.
Pentru asigurarea stabilitatii triplexului este necesara utilizarea unui mediu acid, care prin protonarea noii catene reduce repulsia dintre catena anionica si cea polipirimidinica. In afara de Mg2 alti stabilizatori sunt Ca(NH3)6 sau spermina.
Se presupune ca triplexuri ADN se formeaza in cazul plasmidelor supraincolacite care contin secvente repetitive tip (GAA)9(CIT)9 si (AG)12(TC)12.
Copyright © 2025 - Toate drepturile rezervate