|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
UNIVERSITATEA DE VEST "VASILE GOLDIS"
FACULTATEA DE MEDICINA
SPECIALITATEA : ASISTENTA MEDICALA
ANUL 2
REFERAT
BIOLOGIE CELULARA SI MOLECULARA
TEMA:Transportul membranar
Transportul membranar
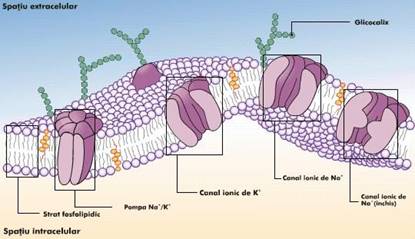
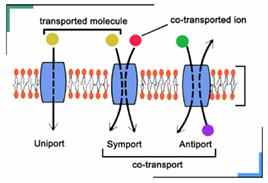
Efectuarea transportului prin membrane sau prin plasmalema este asigurata de proprietatea membranelor sau a plasmalemei de permeabilitate selectiva. Se disting mai multe tipuri de transport, in functie de: consumul energetic: transport pasiv (fara consum de energie) si transport activ (cu consum de energie prin intermediul ATP); marimea moleculei transportate: transport de ioni (molecule mici), transportul macromoleculelor numarul substantelor transportate (sistem uniport, cotransport (simport, antiport)).
TRANSPORTUL PASIV
Difuziunea simpla
poate avea loc prin stratul dublu lipidic sau prin proteine. Ca exemplu pentru
difuziunea simpla prin stratul dublu lipidic putem lua patrunderea substantelor
liposolubile conform coeficientului de partitie intre ulei si apa. Overton a constatat faptul ca patrunderea substantelor
in celule este proportionala cu solubilitatea
substantelor in lipide, masurata prin coeficientul de partitie al substantei
intre ulei si apa. Cu cat acest coeficient este mai
mare, cu atat substanta este mai liposolubila si patrunde mai repede in celule.
Exista insa si exceptii de la regula lui Overton: apa,
ureea si metanolul. Referitor la difuziunea simpla mediata de peptide amintim
transportul ionilor prin intermediul ionoforilor (polipeptide produse de
microorganisme). Exista
2 tipuri de ionofori: transportori mobili (carausi) si de tip canal.
Ionoforii din prima categorie sunt peptide ciclice care au la interior un spatiu polar in care sechestreaza un anumit ion: de
exemplu valinomicina leaga K+. Exteriorul
ionoforului este hidrofob. Ionoforii din aceasta
categorie preiau ionul de pe o suprafata a membranei, difuzeaza prin stratul
dublu lipidic si elibereaza ionul pe fata cealalta, acestia formand
Fiecare proteina transportoare are un loc specific de legare a substratului (substanta transportata). Viteza transportului atinge valoarea maxima (Vmax) caracteristica pentru fiecare transportor, atunci cand acesta este saturat (toate locurile de legare sunt ocupate). Apoi, fiecare transportor are o constanta caracteristica de legare a substantei ce o transporta, numita KM, egala cu concentratia substantei cand viteza de transport atinge ½ din valoarea maxima. Alte caracteristici asemanatoare catalizei enzimatice ar fi: dependenta de pH; inhibitia competitiva prin compusi similari cu substratul; inhibitia necompetitiva prin alte substante (ioni de metale grele) prezente in urme.
Prin difuziune facilitata se realizeaza: transportul anionilor; transportul ureei; transportul glicerolului; transportul unor neelectroliti prin membrana eritrocitului; transportul glucozei; ` transportul aminoacizilor - prin plasmalema celulelor. In cadrul acestui tip de transport, transportatorul este o proteina transmembranara care sufera modificari conformationale reversibile. Intr-o anumita stare conformationala ("pong") locurile de legare pentru substanta ce trebuie transportata sunt expuse la exteriorul bistratului lipidic. In cealalta stare ("ping") aceleasi locuri de legare sunt expuse spre partea opusa a membranei, iar substanta este eliberata. Acesta este mecanismul de "ping-pong". Difuziunea simpla mediata de proteine -canal se deosebeste de difuziunea facilitata prin faptul ca viteza transportului poate fi mai mare decat in cazul difuziunii facilitate si transportul nu se satureaza, adica nu exista Vmax. Unele canale de transport sunt deschise in mod continuu, iar altele numai tranzitoriu. Acestea se numesc "canale de poarta". Unele canale se deschid la legarea unui ligand pe un receptor si le numim "canale cu poarta comandate de ligand". Alte canale dependente de potentialul membranei (voltaj) le numim canale cu poarta comandate de voltaj. Mai exista si canale proteice care se deschid ca raspuns la cresterea concentratiei intracelulare a unor ioni: de exemplu canalele pentru K+ se deschid atunci cand creste concentratia Ca2+ in citosol.
Exemple de canale cu poarta exista in functionarea jonctiunii neuromusculare, in care un impuls nervos produce contractia muschiului. Cel putin 4 canale se deschid in mai putin de o secunda. Descresterea potentialului plasmalemei (depolarizarea) terminatiei neuronului deschide canalul pentru Ca2+. Cresterea concentratiei intracelulare de Ca2+ produce descarcarea veziculelor cu acetilcolina, care se leaga de receptorul acetilcolinei din plasmalema celulei musculare. Acesta este un canal cu poarta comandata de receptor; se produce deschiderea lui, ceea ce permite timp de 1 msec difuziunea Na+ spre interior si a K+ la exteriorul celulei. Cum gradientul de concentratie al Na+ este mult mai mare ca cel al K+, influxul de Na+ depaseste efluxul de K+, deci se produce depolarizarea plasmalemei muschiului. Aceasta depolarizare deschide canalele pentru Na+, dependente de voltaj. Ceea ce produce o unda de depolarizare (potential de actiune) ce se raspandeste pe intreaga membrana musculara. Ca rezultat al potentialului de actiune, se deschid canalele de Ca2+ in citosol. Aceasta crestere brusca a Ca2+ in citosol produce contractia miofibrilelor.
TRANSPORTUL ACTIV

Transportul activ este realizat de proteine transportoare cu o inalta specificitate si are si el caractere comune cu cataliza enzimatica: cinetica de tip Michaelis-Mentel. Ceea ce il deosebeste de difuziunea facilitata este consumul de energie. Transportul ionilor prin plasmalema si alte membrane se realizeaza de proteine care folosesc direct energia din ATP, aceste pompe ionice avand si proprietate ATP-azica. In transportul aminoacizilor si glucidelor prin plasmalema se foloseste energia gradientelor ionice (realizate prin ATP), deoarece transportul acestor substante se face cuplat cu transportul de Na+ (cotransport). In cazul bacteriorodopsinei se foloseste energia luminii pentru pomparea H+. La unele bacterii se foloseste fosfoenolpiruvatul ca sursa de energie pentru mecanismul numit translocare de grup.
Transportul ionilor - pompa de Na+ si K+ din plasmalema si pompa de Ca2+ din plasmalema si din reticulul sarcoplasmic al celulelor musculare.

Plasmalema tuturor celulelor este polarizata, adica prezinta un potential de membrana cu valori cuprinse intre -20mV si -200mV. Fata citoplasmatica a plasmalemei este incarcata negativ, iar cea externa pozitiv. Exista 2 proteine care joaca un rol foarte important in generarea si mentinerea potentialului de membrana: pompa de Na+ si K+, pe de o parte, si canalul de pierdere al K+, pe de alta parte. Concentratia intracelulara de K+, de circa 400mM, este mult mai mare ca cea extracelulara (20mM). Pentru Na+ situatia este inversa, concentratia intracelulara fiind mult mai mica (50mM) fata de cea extracelulara (440mM). Proteina cu canalul de K+ permite difuziunea pasiva a K+ din celula la exterior (in sensul gradientului de concentratie). Dar prin aceasta interiorul celulei devine mai electronegativ si de aceea la o anumita valoare a potentialului de membrana (-75mV) tendinta K+ de a parasi celula datorita gradientului de concentratie este contrabalansata de tendinta K+ de a intra in celula datorita potentialului membranar. Deci gradientul electrochimic al K+ se anuleaza si numarul de ioni de K+ ce ies din celula devine egal cu numarul celor ce intra in celula. Dar canalul de K+ este usor permeabil si pentru Na+. Deci unii ioni de Na+ intra in celula conform gradientului electrochimic al Na+. Prin transport de sarcini pozitive la interiorul celulei scade potentialul membranar, ceea ce permite iesirea altor ioni de K+. prin repetarea proceselor descrise s-ar ajunge la egalarea concentratiilor de Na+ si K+ de o parte si cealalta a membranei si la disparitia potentialului membranar, daca nu ar exista pompa de Na+ si K+ care pompeaza Na+ in afara celulei si K+ in citoplasma (sistem antiport)
. 
Pompa de Na+ si K+ se afla in plasmalema tuturor celulelor animale si este responsabila de mentinerea potentialului membranar, dar si de controlul volumului si de "intretinerea" transportului activ al aminoacizilor si glucidelor. De aceea, peste 1/3 din necesarul de energie al celulei este consumat de aceasta pompa, iar in celulele nervoase se ajunge pana la 70% din consumul energetic sa revina pompei.
S-a descoperit ca pompa de Na+ si K+ este aceeasi proteina cu Na+-K+-ATP-aza, o enzima care scindeaza ATP-ul in ADP si fosfat anorganic (Pi) si care necesita Na+ si K+ pentru activitatea optima, fiind inhibata specific de glicozidele cardiotonice (digitala si oubaina)
Pompele de Ca2+ sunt importante in mentinerea concentratiei scazute de Ca2+ in citosol(10-7 M) fata de o concentratie mult mai mare a Ca2+ extracelular (10-3M). Exista in plasmalema o pompa de Ca2+ numita si Ca2+-ATP-aza care transporta activ Ca2+ la exterior.
Gradientul mare de concentratie al Ca2+ asigura conditii pentru transmiterea de semnale de la exterior la interiorul celulei prin plasmalema. Reglarea concentratiei de Ca2+ in citosol este importanta si pentru procesele de secretie celulara si motilitate.
De asemenea functioneaza o Ca2+-ATP-aza in membrana reticulului sarcoplasmic din muschi, care pompeaza Ca2+ din citosol. Aceasta enzima este destul de bine caracterizata: consta dintr-un singur polipeptid format din circa 100 resturi de aminoacizi, pompeaza 2Ca2+ pentru fiecare molecula de ATP hidrolizata si poate hidroliza pana la 10 molecule ATP/secunda. Cand impulsul nervos depolarizeaza plasmalema celulei musculare, Ca2+ este eliberat din reticulul sarcoplasmic in citosol, stimuland contractia muschiului.
Bacteria Halobacterium halobium are intr-o zona specializata a membranei sale, numita membrana purpurie, proteina bacteriorodopsina ce functioneaza la lumina ca o pompa de protoni, expulzand protonii la exterior, creand astfel potentialul membranar.O caracteristica generala a pompelor ionice este reversibilitatea lor.

Transportul activ cuplat cu gradiente ionice este reprezentat de transportul glucozei si al Transportul glucozei prin plasmalema celulelor din mucoasa intestinala si a celulelor din tubii renali prezinta importanta foarte mare. Astfel, desi filtratul glomerular al rinichiului contine glucoza in concentratie aproape egala cu cea din plasma, in urina nu apare in mod normal glucoza, fiindca ea este reabsorbita in tubii renali printr-un proces de transport activ. Absorbtia intestinala de glucoza ar fi foarte ineficienta daca s-ar face prin transport pasiv, fiindca ar insemna ca in intestin concentratia glucozei sa fie mai mare ca cea din plasma. In realitate, in ambele cazuri, glucoza este transportata impotriva gradientului de concentratie de catre un caraus al glucozei de care se leaga si Na+. Este un sistem simport: Na+ tinde sa intre in celula conform gradientului sau electrochimic si intr-un sens antreneaza glucoza. Cu cat gradientul de Na+ este mai mare si viteza transportului este mai mare; daca se reduce mult gradientul de Na+ se opreste transportul glucozei. Na+ care intra cu glucoza este pompat in afara de Na+-K+-ATP-aza ce mentine gradientul Na+.
Transportul aminoacizilor se face tot prin sisteme simport cu Na+, existand cel putin 5 proteine diferite in plasmalema celulelor animale
Translocarea de grup se intalneste la unele bacterii si consta in fosforilarea unor glucide in cursul trecerii lor prin plasmalema. Sistemul de fosforilare (fosfoenolpiruvat transferaza) este complex, implicand 4 proteine si fsfoenolpiruvatul ca sursa de energie.
Tot la bacterii se mai intalnesc sisteme pentru transportul activ al glucidelor si aminoacizilor ce implica proteine hidrosolubile localizate in spatiul dintre plasmalema si peretele celular.
TRANSPORTUL DIRECT AL UNOR MACROMOLECULE PRIN MEMBRANE
Exista cazuri speciale cand macromoleculele (ADN sau proteine) pot strabate direct membranele celulare fara a se forma vezicule. Asemenea situatii se intalnesc la bacterii in cursul procesului de transformare genetica, in care molecule de ADN trec atat prin peretele celular cat si prin plasmalema. In celulele eucariote proteinele de secretie trec direct prin membrana reticulului endoplasmic rugos pe masura ce sunt sintetizate. Unele toxine bacteriene trec prin plasmalema celulelor animale si isi exercita efectele in citosol. In cele mai multe cazuri macromoleculele sunt trecute prin membrane prin intermediul veziculelor.

TRANSPORTUL PRIN VEZICULE: ENDOCITOZA, EXOCITOZA
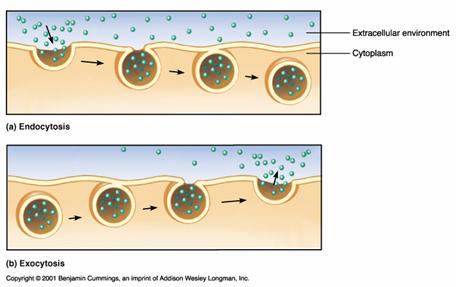
Un mod special de transport prin membranele biologice il reprezinta transportul rpin vezicule. Se disting 3 procese: endocitoza, exocitoza si transcitoza, dupa cum transportul are loc spre interiorul celulei, din citoplasma la exterior sau, respectiv, prin vezicule ce strabat complet celula dintr-o parte in cealalta.
Exocitoza - se produce prin fuziunea unor vezicule din citoplasma cu plasmalema si astfel materialele din vezicule sunt varsate in afara celulei. Asemenea procese apar la nivelul sinapselor, cand veziculele continand mediatori chimici - acetilcolina, noradrenalina - fuzioneaza cu membrana plasmatica a terminatiei neuronului. Mediatorii chimici varsati in spatiul sinaptic se leaga de receptorii specifici din membrana postsinaptica producand depolarizarea ei. In acest fel se asigura transmiterea influxului nervos la nivelul sinapselor.
Endocitoza este de 2 feluri dupa natura substantelor ce patrund in celule: fagocitoza si pinocitoza. Prin fagocitoza patrund substante solide, pe cand prin pinocitoza patrund macromolecule in solutie. In ambele cazuri de endocitoza materialele patrund in celula inglobate in vezicule ce se deprind din plasmalema si care ajunse in citoplasma se numesc endozomi. Acestia se unesc apoi cu un lizozom primar, se formeaza un lizozom secundar in care materialele incorporate sunt digerate, produsii de digestie trecand in citoplasma.
Recunoasterea si atasarea fagocitelor de particule se face prin receptori din plasmalema fagocitului ce recunosc liganzi de pe suprafata particulei.
Inglobarea - se face prin emiterea de catre fagocit a unor prelungiri numite pseudopode, ce inconjoara particula si o inchid apoi complet intr-o vezicula internalizata in citoplasma numita fagozom.
Pinocitoza este de 2 feluri: independenta de receptori si dependenta de receptori. Pinocitoza independenta de receptori se mai numeste endocitoza in faza fluida si se intalneste la multe celule. Pinocitoza dependenta de receptori sau endocitoza mediata de receptori se realizeaza cu ajutorul receptorilor din plasmalema ce recunosc macromoleculele specifice din lichidul extracelular.
BIBLIOGRAFIE
Biologie celulara si moleculara, _ AUREL ARDELEAN Editura VASILE GOLDIS - 2006
Biologia celulara _ MIHAI CRUCE, AUREL ARDELEAN Ed.AIUS CRAIOVA 2002
Lucrari practice de biologie celulara si moleculara _ ARDELEAN PRIBAC _ DAMIAN- Ed. VASILE GOLDIS 2007.
Copyright © 2025 - Toate drepturile rezervate