|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Raport de
Grant: Evaluarea nivelului de eutrofizare a apelor costiere de la litoralul romAnesc al Marii Negre pe baza studiului comunitatilor de polichete
Ca si in cazul apelor dulci, imbogatirea apelor costiere cu substante nutritive duce la aparitia fenomenului de eutrofizare. Daca in apele continentale acest proces are indeosebi cauze naturale, in mediul marin eutrofizarea este provocata de activitatea antropica (Zaitsev, 1991).
Principalele surse ale eutrofizarii sunt: (1) agricultura, care foloseste excesiv ingrasamintele minerale, ceea ce duce la levigarea acestora de pe terenurile agricole si care ajung in cele din urma in mare; (2) industria chimica si arderea combustibililor fosili, datorita emisiilor in atmosfera a oxizilor de azot care apoi ajung in oceane sub forma de nitrati cu apa de precipitatii si care impreuna cu compusii sulfului contribuie la formarea ploilor acide; (3) industria detergentilor, deoarece multi detergentii contin pana la 40% de saruri de fosfor; (4) marile orase situate de-a lungul coastei, care deverseaza apele uzate in mare; (5) deseurile animale provenite de la abatoare si din industria alimentara, care in urma descompunerii elibereaza azotul si fosforul in straturile superficiale ale apei marine.
Grupul de Experti privind Aspectele Stiintifice ale Poluarii Marine (GESAMP) arata ca procesul de eutrofizare antropica a apelor costiere reprezinta una dintre cele mai daunatoare influente umane asupra marilor si oceanelor lumii (GESAMP, 1990, 2001). Acest proces induce in apele naturale o serie de fenomene inter-conexe, reunite sub numele generic de sindrom de eutrofizare (Gomoiu, 1985).
In primul rand, eutrofizarea se manifesta prin dezvoltarea exagerata a algelor planctonice, fenomen cunoscut sub numele de inflorire a apelor sau maree rosie (Zaitsev, 1991). Consecintele acestei dezvoltari exponentiale a microfitelor pelagice constau in modificarea proprietatilor organoleptice ale apei, aceasta capatand o culoare particulara, dependenta de felul algei, un miros specific si o consistenta gelatinoasa, facand-o astfel improprie pentru scaldat. Particulele de aerosoli din zonele cu infloriri pot sa contina substante toxice sau microorganisme. Reducerea transparentei apei si a penetrabilitatii radiatiilor solare, ca urmare a inmultirii fitoplanctonului, duce la reducerea suprafetelor ocupate de macrofite si la disparitia speciilor de alge cu suprafata relativa mica.
Pentru animalele bentonice efectul cel mai nociv al infloririi apelor il reprezinta scaderea concentratiei de oxigen dizolvat in apa din paturile de apa de contact ca urmare a proceselor de descompunere de catre microorganisme a algelor planctonice moarte, care cad pe fund sub forma asa-numitei ploi de cadavre (Gray et al., 2002). Instalarea hipoxiei sau chiar a anoxiei pe zone vaste de fund marin determina mortalitatea in masa a formelor bentonice, inclusiv pestii (Zaitsev & Mamaev, 1997; Surugiu, 2002). Degradarea substantelor organice provenite de la organismele moarte in conditii anaerobe duce la formarea de hidrogen sulfurat toxic.
Eutrofizarea apei duce in timp la declinul biodiversitatii si la schimbarea raporturilor cantitative dintre specii. Astfel, in biocenozele afectate de eutrofizare predomina speciile oportuniste (sensu Grassle & Grassle, 1974), adica acele specii care se dezvolta in masa in conditiile simplificarii structurii cenotice. Printre aceste specii oportuniste, adaptate sa vietuiasca in conditii de mediu improprii, inclusiv la concentratii reduse ale oxigenului dizolvat in apa, se numara si multe specii de polichete (Jakubova & Malm, 1930; Reish, 1970; Losovskaya, 1978). Avcest fapt permite aprecierea gradului de eutrofizare a zonelor costiere prin analiza structurii asociatiilor de polichete, care pot servi in acest fel drept indicatori biologici ai calitatii apelor (Cognetti & Talierico, 1969; Bellan, 1980; Losovskaya, 1983; Pocklington & Wells, 1992).
Alegerea polichetelor ca obiect de studiu s-ar datora si ponderii deosebite pe care acestea il au in desfasurarea proceselor vitale care au loc la nivelul bentosului marin, atat sub aspectul diversitatii specifice, al abundentei numerice cat si al biomasei realizate (Bacescu et al., 1971; Surugiu, 2002). Unele specii filtreaza cantitati apreciabile de apa, eliberand-o de particulele in suspensie, altele reduc cantitatea de substanta organica acumulata in sediment si care este o sursa secundara de poluare.
Toate cele prezentate mai sus constituie argumente puternice pentru necesitatea efectuarii unor studii de impact antropic la coastele romanesti.
Pentru studiul influentei eutrofizarii si poluarii cu materii organice asupra populatiilor de polichete au fost prospectate apele costiere cu diferite grade ale impactului antropic. In functie de aceasta s-au fixat 8 profile perpendiculare fata de linia de tarm, de-a lungul carora s-au prelevat probe de la diferite adancimi (in general, de la 1 m, 10 m si 20 m) (Fig. 1). In fiecare statie s-a luat cate o singura proba. In cazul profilului Agigea, atat datorita localizarii in apropierea Statiunii Biologice Marine Prof. dr. Ioan Borcea de la Agigea cat si a influentei Canalului Dunarea Marea Neagra si a portului Constanta Sud, numarul statiilor si frecventa de prelevare a fost mai mare. Astfel, aici au fost investigate polichetele de la adancimea de 0 m, 2 m, 4 m, 6 m, 8 m, 10 m, 12 m, 14 m si 16 m de cate doua ori pe an. In felul acesta, numarul total al probelor analizate se ridica la 66.
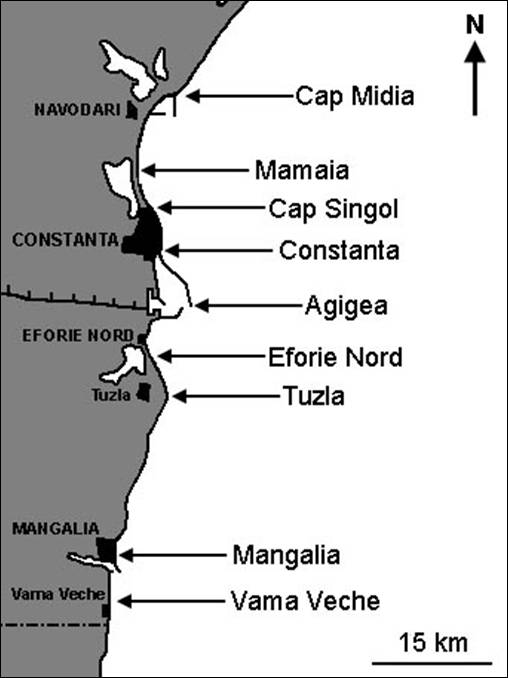
Figura 1. Localizarea punctelor de prelevare a probelor.
In fiecare statie s-a notat data colectarii, pozitia geografica, adancimea si natura substratului. Sporadic au fost masurati si anumiti parametri fizico-chimici precum transparenta, salinitatea, temperatura, concentratia oxigenului dizolvat in apa si cantitatea de materii organice dizolvate (MOD). Coordonatele exacte ale statiilor au fost stabilite cu ajutorul Sistemului Global de Pozitionare (GPS 45). Determinarea adancimilor din fiecare statie s-a facut cu ajutorul profundimetrului de mana. Transparenta apei s-a masurat cu ajutorul discului Secchi. Masurarea temperaturii s-a facut cu ajutorul unui termometru obisnuit de laborator. Salinitatea s-a determinat prin dozarea clorurilor in mediu neutru cu azotat de argint in prezenta cromatului de potasiu ca indicator (metoda Mohr) si transformarea clorinitatii in salinitate dupa formula lui Almazov. Dozarea oxigenului dizolvat in apa s-a facut dupa metoda clasica Winkler. Substantele organice din apa au fost determinate prin oxidarea cu permanganat de potasiu in mediu bazic.
In zona de spargere a valurilor polichetele au fost colectate prin rasturnarea pietrelor sau prin rascolirea sedimentelor moi cu ajutorul unei lopeti. Colectarea de la adancimi mai mari s-a facut din barca sau prin scufundari in apnee sau cu scafandrul autonom. Pentru prelevarea cantitativa de pe substrat nisipos sau malos s-a utilizat o draga de tip van Veen cu deschiderea de 0,1 m2 si o carotiera manuala cu deschiderea de 0,0154 m2, iar in cazul probelor de pe substrat dur s-a recurs la razuirea stancilor cu ajutorul unui cutit de pe o suprafata de 400 cm2, delimitarea facandu-se cu o rama metalica patrata cu latura de 20 cm. Probele astfel prelevate au fost fixate in situ cu formol concentrat, introduse in pungi de polietilena, etichetate si transportate in laborator:
In laborator probele au fost spalate printr-un set de site cu ochiurile de 1 mm, 0,5 mm si 0,125 mm. Materialul retinut pe site a fost transferat in tavite albe din plastic in vederea trierii polichetelor. Sortarea si identificarea polichetelor s-a facut sub lupa binoculara, in unele cazuri fiind nevoie de examinarea chetilor sau a altor structuri la microscop.
Pentru determinarile preliminare s-au folosit cheile prezentate in lucrarile lui Vinogradov si Losovskaya (1968) si Marinov (1977). Precizarea pozitiei taxonomice a unor specii s-a facut apeland, pe cat posibil, la revizuirile cele mai recente.
Dupa ce polichetele au fost identificate si numarate pe specii s-a determinat greutatea umeda a acestora prin cantarire la balanta analitica. Pentru a avea valori comparabile, toate datele au fost apoi extrapolate la metru patrat de suprafata bentala.
In vederea exprimarii unor raporturi cantitative am folosit o varietate de indici care permit caracterizarea structurii si rolului diferitelor specii in comunitatile de polichete:
1) Abundenta (A) desemneaza numarul absolut de indivizi apartinand unei specii prezenti pe o unitate de suprafata (ex./m2).
2) Biomasa (B) reprezinta suma greutatilor umede ale polichetelor prezente pe o unitate de suprafata bentala (g/m2).
![]()
3) Frecventa (F) exprima raportul dintre numarul de statii in care
este prezenta o anumita specie si numarul total de
statii executate intr-o zona oarecare (Bellan, 1964b). Se
calculeaza dupa relatia:
in care: F - frecventa;
p - numarul statiilor in care este prezenta specia data;
P - numarul total al statiilor cercetate;
100 - factorul pentru raportarea procentuala.
In functie de valorile frecventei s-au stabilit 3 categorii de specii:
F < 25% - specii accidentale;
F = 25-50% - specii accesorii;
F > 50% - specii constante.
4) Bogatia specifica (S) reflecta numarul de specii prezente intr-o anumita statie sau proba.
![]()
5) Indicele de diversitate (H) exprima gradul de structurare a
biocenozei si se calculeaza folosind ecuatia lui Shannon-Wiener:
unde: S - numarul de specii;
pi - ponderea speciei i la reprezentarea valorii totale (abundenta relativa).
6) Echitabilitatea (E) este un parametru biocenotic care caracterizeaza gradul de uniformitate a distributiei indivizilor pe diferitele specii. S-a calculat conform indicelui de echitabilitate al lui Pielou:
![]()
unde: Hmax - valoarea maxima posibila a diversitatii, realizata in cazul in care toate speciile ar fi reprezentate prin acelasi numar de indivizi ( = log2 S);
H - valoarea calculata a diversitatii lui Shannon.
Litoralul romanesc (Fig. 2), cu o lungime totala de 228 km, se intinde intre bratul Musura (45°12' lat. N si 29°40' long. E), care face granita comuna cu Ucraina, si frontiera cu Bulgaria ce trece la sud de Vama Veche (43°43' lat. N si 28°35' long. E). Linia de coasta, avand o orientare generala de la nord la sud (cu exceptia sectorului central Ciotic-Midia), este foarte putin crestata, fiind practic lipsita de golfuri sau promontorii. Totusi, de-a lungul litoralului romanesc putem distinge cateva portiuni mai mult sau mai putin intrate in mare, cum ar fi Capul Midia, Capul Tuzla si Capul Aurora. De asemenea, trebuie precizat ca in nordul litoralului, datorita aluviunilor aduse de Dunare, are loc o permanenta modelare a liniei de tarm. Astfel, la sud de gura Sf. Gheorghe s-a format insula Sacalin, iar golful Musura evolueaza spre inchidere si transformarea lui intr-un liman (Bacescu & Dumitrescu, 1958).
Morfologic si genetic tarmul romanesc poate fi impartit in doua sectoare:
1. Sectorul nordic (143 km lungime sau 63%), situat intre meleaua Musura si Capul Midia, este un tarm acumulativ, jos, cu plaje nisipoase intinse, de origine fluvio-marina. Este format din cordoane litorale cu nisipuri fine si scoici, ce separa marea de salba de lacuri litorale.
2. Sectorul sudic (85 km lungime sau 37%), situat intre Capul Midia si Vama Veche, este un tarm inalt (2-40 m), de abraziune, alcatuit din faleze formate din calcare sarmatiene, acoperite cu o patura groasa de depozite loëssoide cu intercalatii de argila rosie. Linia inalta si abrupta a falezelor din sud, supusa pe alocuri actiunii directe a valurilor, este intrerupta in dreptul vechilor vai si lagune marine (portiunea Navodari-Mamaia, intre cele doua Eforii, Costinesti, Mangalia, Vama Veche) de cordoane nisipoase de 1-5 m inaltime care se prelungesc si in mare, formand insule printre fundurile de piatra. In unele locuri, calcarele sarmatiene se pot continua in mare cu platforme calcaroase (ca de exemplu la Agigea, Costinesti si Vama Veche) pana la adancimi de 11-16 m.
Platforma continentala din dreptul tarmului romanesc, delimitata la est de izobata de 200 m, se ingusteaza treptat de la nord la sud, latimea acesteia variind intre 100 si 200 km in nord si pana la 50 km in dreptul Mangaliei. Respectiv, panta medie a platoului continental este foarte mica in nord (0,0014) si se accentueaza spre sud (0,0022). Suprafata totala a platformei continentale romanesti este de aproximativ 23.000 km2 (Bacescu et al., 1971).
Din punct de vedere al reliefului fundului, platforma continentala romaneasca se prezinta ca o campie submarina uniforma, plana, usor inclinata de la vest-nord-vest catre est-sud-est, intersectata de un sistem de vai submarine - adevarate canioane submarine - pe fundul carora se gasesc moluste subfosile intr-o perfecta stare de conservare (Dreissena, Theodoxus, Micromelania etc.), ca si cantitati impresionante de schelete de Coscinodiscus (Bacescu, 1961).
In tot lungul litoralului romanesc exista o serie de lacuri litorale, vechi limanuri si lagune marine. Multe dintre acestea sunt dulci, cu salinitati mai mici de 0,5 (Razim, Golovita, Zmeica, partea nordica a lagunei Sinoie, Gargalac, Tasaul, Siutghiol, Tabacariei, Neptun, Jupiter, Tismana, Venus, Limanu, Hagieni), altele sunt salmastre oligohaline, cu salinitati cuprinse intre 0,5 si 5 (Sinoie langa cetatea Histria, Istria, Nuntasi, Tuzla, Agigea, Belona, Costinesti, Tatlageac, mlastina Hergheliei), sau mezohaline, ca de exemplu lacul Mangalia a carui salinitate este de 11,9-15,2, in timp ce apele Techirghiolului sunt hiperhaline, prezentand o salinitate de 54,5-61,4.
Este bine cunoscut faptul ca natura fundului reprezinta factorul principal care determina distributia vietuitoarelor bentale. Desi varietatea sedimentelor intalnite pe platforma continentala romaneasca a Marii Negre este destul de mare, Bacescu et al. (1971) deosebesc in principal 8 tipuri de baza.
3.2.1. Sedimentele nisipoase sunt prezente de-a lungul intregului litoral romanesc si ocupa o suprafata aproape continua de aproximativ 700 km2 (Petranu, 1997). Latimea benzii acoperite de nisipuri variaza foarte mult. Astfel, in vecinatatea gurilor Dunarii banda nisipoasa are o latime variind intre 1320 si 2520 m si coboara la adancimi de 6-10 m, in zona Portita Mamaia pana la 8800 m si coboara la 22 m, iar in sud plajele submerse coboara intre 12 si 22 m si au o latime intre 1750 si 5550 m (Gomoiu, 1969).
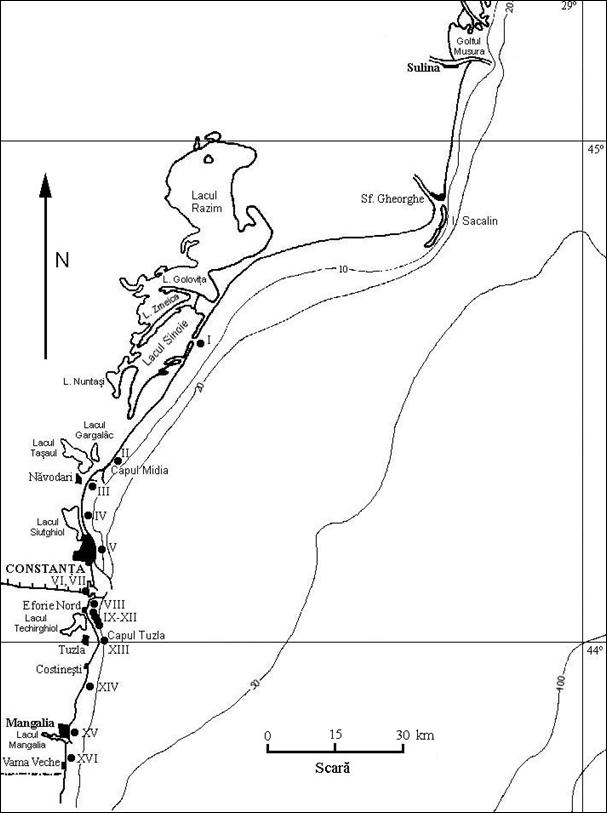
Figura 2. Litoralul romanesc al Marii Negre.
In zona situata la nord de Constanta nisipurile sunt fine (cu diametrul mediu al granulelor de 132-350 μm), cuartoase, de origine fluviala, care formeaza plaje de cateva sute de metri latime intr-o banda aproape continua, de la Sulina la Mamaia. Caracterul fin al sedimentelor se mentine, in linii generale, atat la nisipurile ce formeaza dunele si plajele intinse cat si la cele submerse. Pe masura ce adancimea creste sedimentele devin din ce in ce mai fine, pana ce acestea sunt inlocuite de maluri. Mai trebuie adaugat ca in alcatuirea cordoanelor litorale din nordul litoralului romanesc printre sedimentele fine pot sa apara lentile de sedimente nisipoase cu o structura granulometrica mai grosiera, rezultate in urma amestecului cu scoici maruntite (Bacescu et al., 1971).
In zona situata la sud de Constanta nisipurile formeaza plaje inguste la baza falezelor, intrerupte din loc in loc de stanci calcaroase. Nisipurile de aici prezinta variatii granulometrice ale sedimentelor foarte mari. Nisipurile supra- si pseudolitorale sunt in general medii si grosiere (in care predomina fractiunile granulometrice de 759-1001 μm), calcaroase, de origine biogena (sfaramaturi de moluste, midii in special). Odata cu cresterea adancimii acestea sunt inlocuite de nisipurile cu granulatie medie (cu diametrul mediu al granulelor cuprins intre 203 si 433 μm), tot cochilifere, iar acestea, la randul lor, sunt inlocuite de nisipurile fine, minerale (Gomoiu, 1963, 1969).
3.2.2. Nisipurile maloase formeaza un brau ingust care delimiteaza fundurile acoperite de sedimente nisipoase de cele maloase. Inlocuirea nisipurilor cu nisipuri maloase si maluri nisipoase se face in mod foarte variat, atat in functie de apropierea sau departarea de gurile Dunarii sau a altor fluvii ce aduc aluviuni cat si de o serie de factori hidrologici. Astfel, in fata gurilor Dunarii procesul sedimentarii fiind intens, nisipul malos apare intre 5 si 8 m adancime. Paralel cu aceasta, datorita dinamicii intense a curentilor locali, are loc si un transport permanent si intens de sedimente tarate, determinand o variatie continua a calitatii fundului pana la 16-18 m adancime. In zonele de la sud de Portita, intre Chituc si Constanta, datorita unor conditii de substrat mai stabile, substituirea nisipurilor cu nisipuri maloase are loc de la 18-20 m pana la 30-35 m adancime.
3.2.3. Substratul dur este reprezentat in general de calcare sarmatiene, fie sub forma de platforme intinse de piatra fie ca pietre izolate dispuse neregulat. Fundurile stancoase sunt prezente mai ales in sudul litoralului romanesc, intre Capul Midia si Vama Veche si patrund in adancime pana la 7 m la Capul Midia si 23 m la Mangalia, fiind treptat acoperite de sedimente mobile. Latimea zonei pietroase poate varia intre cateva zeci de metri si 4 km. Falezele, alcatuite din calcare oolitice sarmatiene, intalnesc nivelul marii in mod direct numai in puncte izolate (la Agigea, Costinesti si Mangalia).
In principiu, substratul stancos prezinta 3 forme de relief:
a) placa neregulata cu crapaturi si bolovanis de dimensiuni moderate; denivelarile bruste nu sunt mai mari de 1,5 m, suprafetele orizontale sau putin inclinate sunt dominante in comparatie cu cele verticale sau puternic inclinate; este forma de relief care domina zonele putin adanci, intre 0 si 5 m, repetandu-se apoi spre larg in anumite locuri, la sud de Constanta intre 10 si 14 m adancime, formand zona de tranzitie intre salac si platforma regulata de la marginea dinspre larg a substratului pietros;
b) asa-zisul salac, care reprezinta liniile de falie, paralele cu coasta, cu ingramadiri de blocuri de dimensiuni mari, cu aspect morenaic, desprinse din placa calcaroasa si dispuse neregulat pe fundamentul platformei, cu variatii bruste de nivel ce ating amplitudini de 4-6 m pe o distanta de numai 5-10 m, determinand predominarea suprafetelor verticale sau puternic inclinate; acest tip de relief apare pe portiuni intinse, mai ales intre 5 si 12 m adancime, latimea fasiei ocupate nefiind insa mai mare de 40-50 m;
c) portiuni de platforma propriu-zisa, cu suprafata aproape neteda, fara ca schimbarile bruste de nivel sa depaseasca 0,5 m amplitudine pe verticala, si lipsita in mod practic de blocuri de piatra izolate; aceste portiuni sunt caracteristice mai ales intre 4 si 6 m adancime, repetandu-se apoi mai spre larg, in apropierea limitei inferioare a pietrei.
La nord de Constanta substratul pietros este de natura antropica, reprezentat de recife artificiale cu rol de sparge-val (stabilopozi, evidate, bolovani) ca cei din baia Mamaia (Gomoiu, 1997), de constructiile hidrotehnice ale porturilor Midia si Tomis, precum si de digul canalului navigabil Sulina.
O varianta aparte a substratului dur o reprezinta fundurile argilos-marnoase, dispuse sub forma de insule izolate, intercalate atat in cadrul suprafetelor dominate de sedimentele maloase sau nisipoase cat si in cadrul celor pietroase, la adancimi de 3-12 m. Acest tip de substrat a fost localizat in zona gurilor Dunarii (Bacescu et al., 1965b), la Capul Tabacarie (Gomoiu & Müller, 1962) si la Agigea (Surugiu, 2002). Fundurile de marna argiloasa sunt uneori puternic erodate, cum sunt cele de la Agigea, fara ca denivelarile locale ale substratului sa depaseasca amplitudinea de 1 m.
3.2.4. Scradisul recent este compus din ingramadiri de cochilii de moluste marine actuale (Spisula, Mytilus, Chione, Paphia, Abra, Cerastoderma, Hinia, Cyclope etc.). Se gaseste la adancimi variabile, in functie de jocul curentilor. Un astfel de depozit alcatuit din scradis recent a fost gasit in zona Chituc-Vadu, la adancimi cuprinse intre 12 si 14 m. In unele cazuri scoicile goale au un aspect ruginiu, fiind acoperite cu o pelicula fina de oxizi de fier, in altele acestea sunt de consistenta cretoasa, friabile. Datorita formarii in aceasta zona a unor curenti locali, mai mult sau mai putin circulari, suprafetele ocupate de acest scradis sunt extrem de sarace in sedimente fine, proportia acestora crescand insa catre zonele marginale (pana la 20% din volumul sedimentului). Extensiunea maxima a fasiei ocupate de scradis (de 7-8 km) se gaseste la latitudinea Portitei, latimea ei descrescand treptat catre sud.
In fata gurilor Dunarii, datorita sedimentarii celor mai fine fractiuni de suspensii aluvionare fluviale (cu dimensiunile particulelor cuprinse intre 20 si 10 μm), ia nastere un substrat malos pelitic, foarte putin consistent si bogat in detritus vegetal malurile cu Nephtys. Aceste maluri se situeaza la o distanta de 2-8 km de tarm, intre 12-15 si 20-22 m adancime. Sub actiunea curentilor aceste petice de mal, foarte sarace in scradis (sub 3% din volumul sedimentului), isi pot modifica dimensiunile si chiar pozitia.
O varietate aparte de sedimente o formeaza malurile portuare, negre-albastrui, grase, de tip sapropelic, de cele mai multe ori cu miros puternic de hidrogen sulfurat (Tiganus, 1982b).
3.2.5. Malurile cu Mytilus ocupa in general fundurile cuprinse intre 20 si 60 m adancime, formand o banda continua in intreg bazinul Marii Negre. Se caracterizeaza prin predominarea malurilor cenusii, care mai spre larg pot deveni albastrui, aleuritico-argiloase, destul de mobile, unsuroase la pipait. Aceste maluri, dispuse in straturi de 20-40 cm grosime, se afla in amestec cu scoicile diverselor moluste, ingloband astfel cea mai bogata tanatocenoza din Marea Neagra. In unele zone, in special in zona din fata gurilor Dunarii, aceste maluri sunt acoperite de un strat de 1-4 mm de mal galben, cu aspect de gel coloidal, reprezentand sedimentele cele mai recente.
3.2.6. Sedimentele cu Phyllophora reprezinta varietati ale malurilor cu Mytilus sau ale celor cu Modiolus, in care gasim o bogata tanatocenoza incrustata cu algele calcaroase rosii ale genului Lithothamnion (L. crispum, L. cystoseirae si in special L. propontidis). Coloniile lor moarte, crustoase, impreuna cu valvele de midii pe care se dezvolta, pot forma suprafete intinse, imprumutand substratului o consistenta dura, favorabila fixarii tufelor de Phyllophora nervosa, Ph. brodiaei si Ph. membranifolia.
3.2.7. Malurile faseolinifere, calcaroase, albe, inlocuiesc spre larg pe cele cu Mytilus si acopera fundurile incepand cu 70 m adancime pana la limita platformei continentale romanesti. Aceste maluri de adanc se gasesc in straturi mai subtiri decat malurile precedente (5-20 cm grosime). Ele inglobeaza de asemenea diverse scoici moarte, de Modiolus phaseolinus in special, tanatocenoza fiind ceva mai saraca din punct de vedere calitativ.
Intre malurile faseolinifere de la 80-120 m, pe scradis subfosil de Modiolus, se formeaza o centura de concretiuni fero-manganoase. Acestea se prezinta fie sub forma unei pelicule de oxizi ce acopera valvele de Modiolus, fie sub forma de noduli in care suportul reprezinta mai putin de 20% din volum. Nodulii fero-manganosi, mai mult sau mai putin sferici, de culoare alba-cenusie, pot avea dimensiuni de pana la 2 cm diametru.
3.2.8. Paleoscradisul de tip caspic este dezvoltat mai ales pe fundul vailor submarine, si la adancimi mai mari de 120 m, indicand o abundenta masiva a scoicilor de Dreissena caspia, D. polymorpha, D. rostriformis, D. distincta, Adacna, Monodacna, Micromelania spica, Theodoxus etc. De fapt, aici deosebim doua tipuri de scradis: un orizont superior sau faciesul conchiolinifer (situat intre 90 si 160 m adancime), cu tanatocenoza faseolina mai mult sau mai putin subfosila, cu rare cochilii de tip ponto-caspic si un orizont inferior sau faciesul paleodreissenifer, care se intinde pana la marginea platformei continentale si cuprinde paturile sedimentare de mal alb, acoperite de o foarte bogata si pura tanatocenoza fosila, dominata de Dreissena, in care scradisul reprezenta pana la 90% din volumul sedimentului. In acest mal abunda vertebrele si placile dermale ale lui Syngnathus schmidti, alaturi de resturile diatomeelor planctonice ale genurilor Hyalodiscus si Coscinodiscus.
Intre 200 si 1500 m adancime se intinde domeniul malurilor negre, iar mai jos de 1500 m se intalnesc malurile calcaroase cenusiu-deschise, bogate in carbonat de calciu.
Desfasurarea vietii bentonice, in afara de natura substratului, depinde in mare masura si de conditiile fizico-chimice oferite de patura de apa de contact. Parametrii fizico-chimici ai apei variaza in functie de o multitudine de factori, principalii factori fiind insa adancimea absoluta si departarea fata de gurile Dunarii.
3.3.1. Valurile si vanturile. Viata bentonica este influentata in mod direct de catre valuri doar in etajele bentice superioare (supralitoral si pseudolitoral). Deoarece actiunea miscarilor ondulatorii ale suprafetei marii se resimte si in adancime (uneori chiar si la 20 m), valurile joaca un rol important si in determinarea structurii bionomice a etajelor bentice mai profunde. Actiunea valurilor se manifesta in primul rand prin transportul sedimentelor de pe fundurile moi (maluri, nisipuri). In urma acestui transport are loc si o depunere fractionata a sedimentelor in functie de granulometria lor, realizandu-se astfel o diferentiere corespunzatoare a biotopului care se reflecta si in compozitia calitativa si cantitativa a bentosului.
Actiunea mecanica a valurilor in zona litorala depinde de mai multi factori, cum ar fi adancimea, configuratia tarmului, relieful fundului si prezenta sloiurilor de gheata, cand acestea se formeaza.
Directia de propagare a valurilor este determinata de directia predominanta a vanturilor. In partea nord-vestica a Marii Negre directia predominanta a vanturilor este din sectorul nordic, prin urmare si directia de propagare a valurilor va fi mai ales dinspre nord si nord-est. Deoarece sectorul nord-vestic al Marii Negre prezinta cele mai frecvente perturbatii atmosferice, agitatia marii aici este aproape continua, mai ales in sezonul rece.
Cele mai inalte valuri sunt produse de vanturile care sufla din directia nord-est, care, mai ales pe timp de iarna, pot depasi 3,5 m inaltime la o viteza a vantului de 30-40 m/s. Valurile provocate de vanturile din est si din sud sunt mai mici, de 3 si respectiv 1 m inaltime. Viteza medie anuala a valurilor este de 7,1 m/s la Sulina, 4,3 m/s la Constanta si 3,4 m/s la Mangalia (Bacescu et al., 1971).
Actiunea distructiva a valurilor asupra hidrobiontilor este amplificata in timpul iernilor aspre, cand la tarm se formeaza banchize de gheata, la care se pot adauga sloiurile aduse de Dunare in mare. Bucatile de gheata, puse in miscare de catre valuri si vant, pot distruge partial sau total organismele bentonice din apele putin adanci (Borcea, 1931; Bacescu, 1954).
3.3.2. Curentii. Curentii marini de suprafata, care iau nastere sub actiunea miscarilor maselor de aer, influenteaza viata bentonica prin aducerea unor ape cu salinitati scazute in timpul viiturilor Dunarii. Tinand cont de predominarea vanturilor ce bat dinspre nord, orientarea curentilor marini de suprafata in dreptul litoralului romanesc este de la nord spre sud. Viteza acestui curent este de 0,20-0,56 m/s. In perioadele de vara si cand vanturile cu caracter constant lipsesc apar o serie de curenti marini de deriva cu o dinamica neregulata. Directia curentului principal cu orientarea nord-sud mai este perturbata si din cauza configuratiei tarmului, reliefului fundului si a gurilor de varsare ale Dunarii. Configuratia coastei determina aparitia unor curenti turbionari eliptici anticiclonali (care se rotesc in sensul acelor de ceasornic) in meleaua Musura si in baia Portita. Curenti turbionari locali asemanatori s-au remarcat si la sud de digul canalului Sulina, de portul Midia si de noul port Constanta Sud - Agigea, unde curentul circular principal a fost deviat spre larg.
Vanturile pot pune in miscare masele de apa de pana la 20 m adancime in zona litorala din partea nordica a coastelor romanesti si de pana la 10 m adancime in partea sudica. Prin urmare, orientarea curentilor de fund, pana la 20-25 m adancime, este de la nord spre sud.
In timpul vanturilor puternice de vest, cu un caracter mai constant, care imping masele de apa superficiala din apropierea coastei spre larg, ia nastere un curent compensatoriu de fund cu sens opus, care urca panta fundului.
Curentului de suprafata cu directia nord-sud ii corespunde un contracurent de profunzime, in sens invers, de la sud-est la nord-vest, situat la 50-100 m adancime, care aduce ape sarate dinspre Bosfor.
3.3.3. Temperatura. In ceea ce priveste variatia temperaturilor maselor de apa de contact in zona platformei continentale, putem distinge doua zone:
- portiunea de fund cuprinsa intre 0 si 30 m adancime, ce prezinta o amplitudine larga a variatiilor de temperatura in functie de sezon;
- zona adancimilor mai mari de 30 m, in care variatiile temperaturii sunt mici.
In zona de mica adancime temperatura apei prezinta variatii mari, fiind supusa in mare masura, diminuat ca valoare si decalat in timp, evolutiei temperaturii aerului. Local, evolutia temperaturii apelor mai este dictata de masele de apa fluviale si de curentii ascendenti de adancime.
Pe parcursul cercetarilor noastre apele de mica adancime (0-10 m) au prezentat o amplitudine a variatiilor de temperatura cuprinsa intre 26°C (august) si temperatura de inghet a apei marine (februarie).
In timpul lunilor de iarna (decembrie-februarie) temperatura apei creste odata cu adancimea. Temperaturile de iarna prezinta valori cuprinse intre 0,5 si 2,0°C la suprafata si intre 5,2 si 7,7°C la 25 m adancime. In dreptul gurilor de varsare ale Dunarii in sezonul rece, datorita faptului ca temperatura apelor dunarene este mai scazuta decat cea a apelor marine (uneori chiar cu 5,8°C), in limitele adancimilor de 0-10 m, se pot inregistra frecvent temperaturi negative (-1,3°C). Din acest motiv, in iernile mai severe (cum ar fi cazul iernilor 1953/1954, 1962, 1972/1972 si 1995/1996), marea poate ingheta pe o distanta de cateva zeci sau chiar sute de metri spre larg.
La sfarsitul lunii martie si inceputul lunii aprilie se produce homotermia de primavara, cand intre 0-40 m adancime temperatura apei variaza intre 5 si 6°C. Din acest moment are loc o incalzire progresiva a straturilor de apa superficiale.
In perioada calda a anului (mai-septembrie) temperatura apei din apropierea fundului scade cu cresterea adancimii. Temperatura paturii de apa de contact poate creste pana la 24°C la 15 m adancime si peste 10°C la 25 m. In zona de amestec a apelor de la gurile Dunarii se constata usoare ridicari ale temperaturii apelor costiere datorita aportului apelor fluviale care sunt cu 0,5-3°C mai ridicate decat temperatura apelor marine. Incalzirea puternica din lunile iulie-septembrie determina aparitia termoclinei, care in general se situeaza intre 15-30 m adancime, fiind limitata in partea superioara de izoterma de 21°C, iar in partea inferioara de izoterma de 10°C. In luna octombrie termoclina coboara sub 30 m adancime. Pozitia termoclinei este puternic influentata de curenti si de vant. Astfel, in urma vanturilor persistente din vest si nord-vest, datorita impingerii apelor costiere bine incalzite spre larg, are loc o racire brusca a apelor pana la temperaturi de 9-11°C, fenomen cunoscut sub numele de holodnic.
In luna octombrie se petrece o omogenizare termica a apelor, ducand la uniformizarea temperaturilor de pana la 25-30 m adancime. Aceasta omogenizare are loc in majoritatea cazurilor la o temperatura de 17°C. In luna noiembrie se produce homotermia de toamna, cand toata patura de apa pana la 35 m adancime prezinta temperaturi de 10-12°C. Din acest moment racirea apelor de mica adancime devine evidenta fata de masele de apa de fund aflate la adancimi mai mari de 10 m, ceea ce marcheaza inceputul unei raciri progresive in timpul iernii.
Zona adancimilor mai mari de 30 m prezinta conditii termice mult mai constante. Aici amplitudinea oscilatiilor de temperatura scade odata cu cresterea adancimilor. Astfel, la adancimi cuprinse intre 30 si 50 m temperatura apelor de contact oscileaza intre 4,3 si 9,5°C, intre 50 si 100 m adancime extremele se situeaza intre 6 si 9°C, iar la adancimi care depasesc 100 m temperaturile variaza doar intre 6 si 8°C.
3.3.4. Salinitatea este direct influentata de curenti si de aportul de apa dulce, astfel incat variaza mult in raport cu departarea fata de gurile Dunarii si de adancime. Analizand caracteristicile salinitatilor la fund pe platforma continentala romaneasca s-a observat in fata gurilor Dunarii, ca o consecinta a existentei mecanismelor de upwelling compensatoriu, ca salinitatea apelor de profunzime este neobisnuit de ridicata. Patrunderea lentilelor de apa sarata dinspre est sau sud-est nu are loc in mod permanent, ci numai atunci cand masele de apa ale curentului principal nord-sud nu patrund in adancime pana la fund, adica in cazul unor viituri moderate sau in lipsa unor viituri statornice din sectorul nordic. In atare conditii, salinitati de 18 sau peste aceasta valoare se inregistreaza frecvent chiar si la 10 m adancime, in special la nord de paralela de 45°. Sporadic in fata Deltei Dunarii se pot inregistra lentile de apa de fund cu o salinitate de 19 sau chiar de 20. Difuziunea lor in masele de apa costiera sau in amestecul de ape intermediare contribuie si ea la mentinerea unor salinitati ridicate intre izobatele de 10 si 30 m, salinitatea medie a apelor de fund dintre aceste izobate fiind cea mai ridicata in aceasta regiune comparativ cu tot restul platformei continentale romanesti 18,31. Luand in considerare intreaga platforma din fata coastelor romanesti, salinitati ce trec de 18,8 s-au inregistrat la sud de paralela 44°40' numai la adancimi ce depasesc 60 m.
Salinitatea apelor de contact la fundurile de mica adancime, intre 0 si 30 m, sufera variatii foarte mari, in special in cadrul zonelor de amestec permanent din fata gurilor Dunarii, cat si in baia de la extremitatea sudica a insulei Sacalin si Portita. In aceste zone oscilatiile extreme constituie un fenomen obisnuit si permanent; doar mai la sud de paralela 44°40' avem de-a face cu oscilatii mari, cu caracter sezonier.
Pentru adancimile cuprinse intre 0 si 30 m zona cea mai instabila este aceea din largul Portitei. Aici amplitudinea variatiei salinitatii la fund prezinta valori extreme: intre 12,5 si 18,1 la 0-10 m, intre 14,4 si 18,1 la 10-20 m si intre 16,4 si 18,2 pentru izobatele de 20 si 30 m. Intre aceleasi izobate la extremitatea sudica a litoralului romanesc (la est de Mangalia) valorile extreme oscileaza intre limite mult mai reduse: 15,9-18,3, 16,2-18,2 si, respectiv, 17,4-18,6.
La adancimile ce depasesc 30 m se poate vorbi de existenta unor concentratii saline stabile de cel putin 18. Facand media tuturor datelor de salinitate la fund, media generala pentru spatiul cuprins intre izobatele de 30 si 50 m este de 18,29, desi extremele de variatie oscileaza intre 16,3 si 19,7 la nord de paralela 44°40' si intre 16,8 si 19,2 la sud de aceasta paralela. Media salinitatilor determinate la fund intre izobatele de 50 si 100 m este de 18,37, extremele corespunzatoare fiind cuprinse intre 17,7 si 20,3. Suprafata de fund dintre aceste doua izobate, in fata coastelor romanesti, reprezinta aproximativ jumatate din suprafata intregii platforme. Astfel, este firesc sa se inregistreze trepte de salinitate care duc spre o crestere a mediei odata cu cresterea adancimii. Media salinitatii de fund din dreptul izobatei de 50 m este de 18,21, intre izobatele de 70 si 80 m de 18,43, iar in dreptul izobatei de 100 m media salinitatilor masurate atinge 19,03, desi izohalina constanta de 19 se afla in preajma izobatei de 150 m.
In privinta salinitatii de la adancimi ce depasesc 120 m, media tuturor determinarilor este egala cu 20,46, valorile extreme fiind situate intre 19,04 si 21,40 (Bacescu, 1963).
3.3.5. Concentratia oxigenului. In apele din imediata vecinatate a coastei concentratia oxigenului dizolvat se mentine intre 7,15-11,44 mg/l in sezonul cald si 8,58-15,73 mg/l in sezonul rece.
Atat in aceasta zona cat si in stratul mai adanc, pana la 50 m, variatiile concentratiei oxigenului, precum si saturabilitatea, urmeaza variatiile de temperatura si pe cele ale intensitatii fotosintezei. Astfel, variatiile concentratiei oxigenului la 50 m adancime sunt cuprinse la sfarsitul iernii intre 8,58 si 10,73 mg/l, putand insa ajunge la sfarsitul verii valori minime de ordinul a 5,72-7,15 mg/l. Prin urmare, apele de contact pana la 50 m adancime sunt in mod constant suficient de bine oxigenate, prezentand o concentratie medie anuala a oxigenului in jurul valorii de 10 mg/l. Ocazional, dupa infloririle algale din timpul verii, pot avea loc scaderi drastice ale concentratiei de oxigen de la valori obisnuite la 1,43-2,86 mg/l sau chiar zero.
Sub 50 m adancime concentratia oxigenului prezinta in mod regulat scaderi puternice, oxigenarea relativ slaba fiind una dintre caracteristicile apelor de profunzime. La 100 m adancime oxigenul dizolvat prezinta variatii intre 2,86 si 10 mg/l. Scaderea cea mai brusca se inregistreaza intre 125 si 200 m, de la o amplitudine de variatii intre 2,57 si 10 mg/l pana la disparitia totala a oxigenului la 200 m adancime.
Studiile efectuate recent arata cu sub stratul de salt al concentratiei de oxigen dizolvat in apa (oxiclina) se afla un strat de apa subanaerob, de numai 30-50 m grosime, care desparte apele aerobe de cele anaerobe cu hidrogen sulfurat. In acest strat concentratia oxigenului este de 0,29-0,43 mg/l (Sergeeva si Zaika, 2000).
Intr-o alta lucrare Sergeeva (2000) arata ca si in zona cu hidrogen sulfurat, de la de aproximativ 2000 m adancime, in patura de apa de contact de 1-10 m grosime ar exista mici cantitati de oxigen (intre 0,43 si 1 mg/l), concentratii care permit existenta unor organisme meiobentice.
3.3.6. Hidrogenul sulfurat. In limitele platformei continentale romanesti hidrogenul sulfurat se face simtit incepand de la 150 m adancime. In preajma izobatei de 200 m concentratia H2S poate depasi 1 mg/l (Bacescu, 1961). Cantitatea de H2S din apele de contact corespunzatoare taluzului continental inregistreaza valori cuprinse intre 6,8 mg/l la 645 m adancime si de 10,2 mg/l la 1100 m (Sergeeva, 2000). Mai jos de 1200 m adancime cantitatea de H2S din apa poate ajunge pana la 12 mg/l (Müller, 1995). Trebuie precizat ca exista fluctuatii sezoniere si anuale ale nivelului interfetei oxic-anoxic. De regula, granita hidrogenului sulfurat urca in timpul verii si coboara primavara (Zaitsev & Mamaev, 1997).
In ultima vreme se constata o contaminare cu hidrogen sulfurat si a apelor de mica adancime datorita cresterii gradului de eutrofizare si a instalarii fenomenelor de hipoxie si anoxie, cand H2S se produce in urma proceselor de degradare anaeroba a proteinelor. Prezenta H2S in apele de mica adancime este, insa, tranzitorie, acest gaz acumulandu-se numai in perioada de vara si toamna, cand exista o stratificare termo-salina, iar apele sunt calme si disparand iarna si primavara, cand are loc o omogenizare a coloanei de apa. Intr-o masura mica hidrogenul sulfurat poate proveni in apele de mica adancime si din sursele geotermale submarine (cum ar fi cele de la Mangalia).
3.3.7. Elementele biogene. In ultimii 20-30 de ani s-a constatat o crestere generala a concentratiei apei marine in substante nutritive, atat de natura minerala cat si de natura organica, in toata partea de nord-vest a Marii Negre, inclusiv la litoralul romanesc. Acest fapt este rezultatul cresterii aportului substantelor biogene din fluviile tributare din aceasta parte a marii si in primul rand din Dunare, ca urmare a dezvoltarii rapide a agriculturii, industriei si altor activitati umane in bazinul versant al Marii Negre (Zaitsev, 1991). In cele ce urmeaza este prezentata pe scurt evolutia principalelor elemente biogene de la litoralul romanesc.
In mod obisnuit cantitatea de fosfati din apa creste odata cu adancimea, fiind maxima toamna si minima vara, ceea ce este in stricta concordanta cu dezvoltarea fitoplanctonului care ii consuma (Bacescu, 1961). Valorile medii ale cantitatii de fosfati din apele marine de mica adancime au crescut considerabil de la 10,5 µg/l in anii 1960-1970 pana la 262,0 µg/l in 1986-1988 (Petranu, 1997).
Azotatii se gasesc in cantitati mai mari in apa marii in apropierea coastei si mai ales in zona gurilor de varsare ale Dunarii si in apropierea debuseurilor de ape menajere si aproape ca lipsesc complet la adancimi mai mari. Astfel, valorile medii ale concentratiei azotatilor inregistrate in apropierea Constantei au aratat o crestere spectaculoasa, fiind in prezent de 112,2 µg/l, comparativ cu 22,5 µg/l la nivelul anilor 1960-1970 (Petranu, 1997).
In schimb, concentratia silicatilor in apa marii a scazut continuu incepand cu anul 1970, astfel incat in 1994 valoarea acestora a fost de 266 µg/l, fata de 1029 µg/l in anii 1960-1970. Aceasta descrestere a cantitatii de silicati poate fi explicata prin inmagazinarea acestora in exoscheletul diatomeelor (Petranu, 1997).
In ceea ce priveste substanta organica dizolvata in apa, masuratorile noastre efectuate in zona Agigea-Tuzla au indicat o valoare medie de 3,47 mg O2/l, fiind de cca. 1,8 ori mai ridicata decat cea din anii 1960-1970.
Aceasta crestere a cantitatii de nutrienti s-a tradus prin cresterea troficitatii apelor marine de mica adancime. Astfel daca in anii 1959-1969 apele costiere romanesti prezentau un regim oligo-mezotrofic, acestea au devenit gradual eutrofe in 1969-1970, iar in zilele noastre politrofe (Petranu, 1997). In prezent putem vorbi de aparitia asa-numitului sindrom de eutrofizare (Gomoiu, 1985), care se traduce prin cronicizarea fenomenelor de inflorire a apei, urmate de instalarea hipoxiei sau chiar anoxiei pe mari suprafete ale fundului marin si soldate in final cu mortalitatea in masa a organismelor bentice. Deoarece descompunerea cadavrelor acestora se face cu consum de oxigen, deficitul de oxigen creste si mai mult. In timp eutrofizarea duce la diminuarea biodiversitatii, abundentei si biomasei organismelor bentice, substituirea unor specii prin altele mai rezistente, reducerea transparentei, degradarea macrofitocenozelor etc. (Losovskaya, 1977a, 1978; Zaitsev, 1991). In perioada 1994-1999 aproape in fiecare an la litoralul romanedsc se puteau urmari consecintele unor astfel de infloriri algale, cand la mal se gaseau cantitati impresionante de guvizi, crabi, creveti, midii si multe alte animale moarte (Surugiu, 2002). In pe timpul cercetarilor noastre (vara anilor 2003 si 2004) nu s-au constatat infloriri ale algelor, ceea ce indica o diminuare a eutrofizarii.
Caracterizarea principalelor biocenoze bentale din dreptul litoralului romanesc al Marii Negre s-a facut pe baza observatiilor personale pentru adancimi de pana in 20 m cat si pe baza datelor din literatura pentru adancimile mai mari de 20 m (Fig. 3). Ca sursa de referinta ne-a servit monografia de ecologie bentala a lui Bacescu, Müller si Gomoiu (1971), care sintetizeaza situatia biocenozelor bentale dinainte de anul 1970. Tinand cont insa de schimbarile majore care au avut loc in ultimii 30 de ani, ca urmare a intensificarii poluarii si eutrofizarii, in prezentarea bionomiei bentale a platformei continentale romanesti am folosit lucrarile cele mai recente.
Biocenoza pseudolitoralului nisipos. Elementul caracteristic al biocenozei nisipurilor fine, cuartoase, din zona de spargere a valurilor din sectorul dintre Sulina si Constanta este amfipodul Pontogammarus maeoticus, ale carui populatii prezinta o abundenta medie de 1.150 ex./m2 (Petranu, 1997). Local specia conducatoare poate fi insotita de polichetul Scolelepis cirratulus (syn. Nerine cirratulus). Dintre formele meiobentale ce se dezvolta in numar mare in aceasta biocenoza pot fi mentionate ciliatele microporale ale genurilor Tracheloraphis, Trachelocerca, Trachelonema, Remanella, Geleia si Condylostoma, turbelariatele din genul Otoplana, copepodul Ectinosoma melaniceps, cumaceele, misidele, nematodele etc. (Bacescu et al., 1971).
3.4.2. Subcenoza Mytilaster - Mytilus Balanus. Pseudolitoralul stancos este prezent doar in cateva puncte izolate situate la sud de Constanta, acolo unde faleza calcaroasa sau bolovanii si pietrele desprinse din aceasta sunt supuse actiunii directe si continue a valurilor (Agigea, Costinesti si Mangalia). Extinderea pseudolitoralului stancos este variabila, functie de starea vremii. Aceasta zona este populata de putine organisme capabile sa reziste actiunii mecanice a valurilor pe de o parte si exondarii temporare pe de alta. In functie de configuratia substratului stancos aici putem deosebi suprafete expuse direct actiunii valurilor si suprafete mai adapostite.
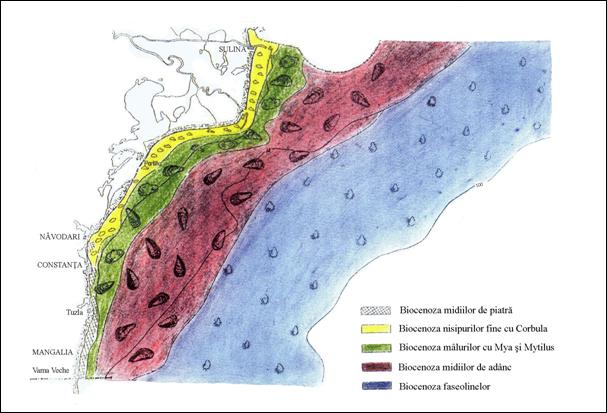
Figura 3. Principalele biocenoze bentale din dreptul litoralului romanesc al Marii Negre (dupa Siganus & Dumitrache, 1992 din Petranu, 1997).
Specia cea mai caracteristica pentru suprafetele stancoase expuse direct actiunii valurilor este Mytilaster lineatus. Deoarece acestei specii conducatoare i se asociaza adeseori Mytilus galloprovincialis si ciripedul Balanus improvisus, aceste trei specii determina o asociatie caracteristica - subcenoza Mytilaster Mytilus Balanus (Bacescu et al., 1971).
Reprezentanti obisnuiti ai acestei subcenoze sunt algele verzi ale genului Enteromorpha, care acopera adesea in intregime stancile litorale si poliplacoforele Middendorfia caprearum si Lepidochitona cinerea. In spatiile interstitiale ale coloniilor celor trei specii sesile care desemneaza subcenoza se pot instala unele elemente ale faunei vagile provenite din etajele inferioare (de ex. polichetele Spio decoratus, Grubeosyllis clavata, harpacticoidul Cumella perplexa etc.).
Pe suprafetele mai putin expuse actiunii valurilor, cum ar fi, de exemplu, crapaturile mai adanci din roca sau spatiile dintre blocurile de stanca ce formeaza digurile porturilor si plajelor, apar frecvent nemertieni, polichetul Platynereis dumerilii, crabul Pachygrapsus marmoratus si izopodele Jaera nordmanni, Idotea baltica basteri, Sphaeroma serratum si S. pulchellum.
Un loc intermediar intre pseudolitoralul stancos si cel nisipos il ocupa blocurile de piatra izolate din zona de spargere a valurilor, dispuse direct pe substratul nisipos. Partea lor inferioara poate concentra anumite elemente caracteristice cum ar fi polichetul Namanereis littoralis, melcul pulmonat Ovatella myosotis, amfipodele Hyale pontica si Marinogammarus olivii, larve de chironomide, diferite turbelariate mici, nemertieni, nematode, ostracode, harpacticide, halacaride etc.
Biocenoza nisipurilor fine cu Corbula. Aceasta biocenoza este dezvoltata mai ales in partea de nord a litoralului nostru. In lungul litoralului romanesc suprafata ocupata de nisipurile cu Corbula depaseste cu putin 600 km2, avand extensiunea maxima intre Mamaia si Portita. Substratul este un nisip fin, cuartos-micaceu, amestecat in unele cazuri cu sfaramaturi de cochilii.
Studiile speciale efectuate asupra nisipurilor sublitorale, care pot ajunge in fata litoralului romanesc pana la 22 m adancime, au completat considerabil reprezentarile noastre in ceea ce priveste compozitia calitativa si cantitativa a vietii de aici (Bacescu et al., 1957, 1965a, 1965b, 1967; Tiganus, 1982a, 1990).
Elementul cel mai caracteristic si cel mai reprezentativ al biocenozei este mica bivalva Corbula mediterranea, care prezinta un coeficient de constanta de 100%, o densitate medie de 3700 ex./m2 si o biomasa de aproape 30 g/m2. Specii insotitoare permanente ale acestei biocenoze sunt polichetul Spio decoratus (cu 800 ex./m2), scoica Cerastoderma edule (300 ex./m2) si amfipodul Ampelisca diadema (10.000 ex./m2). In ultima vreme aceasta biocenoza a fost invadata de bivalvele Mya arenaria (240 ex./m2) si Cunearca cornea (cunoscuta sub numele de Scapharca inaequivalvis), de polichetele Neanthes succinea (264 ex./m2) si Polydora cornuta si de crabul-albastru Callinectes sapidus, fapt care a dus la o crestere considerabila a biomaselor realizate de aceasta biocenoza. Populatii numeroase prezinta si creveta-de-nisip (Crangon crangon). Astfel, dupa o puternica inflorire din august 1999 am gasit sute de mii de exemplare din aceasta specie aruncate pe plaja de la Mamaia. Ciripedul Balanus improvisus, desi este o forma caracteristica fundurilor tari, prezinta densitati ridicate (in medie de 400 ex./m2), fixandu-si casutele sale calcaroase pe cochiliile molustelor. Dintre alte specii, mai rare, dar caracteristice nisipurilor sublitorale fine, pot fi citate nemertienii Amphiporus hastatus, Arenonemertes microps, Prostomatella arenicola si Pontolineus arenarius, polichetele Eteone picta, Hediste diversicolor, Capitella capitata si C. minima, molustele Cyclope neritea, Hinia reticulata, Chione gallina, cumaceele Iphinoe maeotica si Pseudocuma longicornis pontica, misidul Gastrosaccus sanctus, bigecul Diogenes pugilator si gebia Upogebia pusilla. Dintre formele meiobentale tipic psamobionte fac parte nematodele Enoplus littoralis, Epsilonema sp., Sabatieria sp., ostracodele Pontocythere bacescui si Cyprideis littoralis, harpacticoidele Ectinosoma intermedia, E. elongatum, Canuella perplexa, C. furcigera, Harpacticus flexus si Microarthridion litorale.
3.4.4. Biocenoza midiilor de piatra. Fundurile stancoase sunt raspandite in zona situata la sud de Constanta, ocupand in fata coastelor romanesti o suprafata de aproximativ 70 km2. Acestea cuprind portiunea de fund marin care este tot timpul imersa si ajunge pana la adancimi de 11-17 m. In ciuda suprafetei relativ restranse a substratului dur, bogatia in substante nutritive cat si epibiozele bogate date de algele macrofite (Enteromorpha intestinalis, E. linza, Cladophora sericea, Cl. laetevirens, Bryopsis plumosa, Ulva rigida, Ceramium elegans, C. arborescens, C. diaphanum, C. rubrum, Porphyra leucosticta etc.) si de unele macrobentonte sesile (Mytilus galloprovincialis, Mytilaster lineatus, Balanus improvisus etc.) determina o vasta diferentiere de nise ecologice si microbiotopuri, oferind conditii favorabile pentru dezvoltarea unei vieti foarte bogate si diversificate (33 specii macrobentice in prezent).
Specia cea mai caracteristica a sublitoralului stancos este Mytilus galloprovincialis, desemnand biocenoza midiilor de piatra (Bacescu et al., 1963; Tiganus, 1978, 1981). Speciile insotitoare cele mai caracteristice sunt Actinia equina, Balanus improvisus si speciile genului Corophium.
In cadrul biocenozei midiilor de piatra, datorita existentei variatiilor largi in ceea ce priveste aspectul si intinderea substratului, putem distinge doua subcenoze bine delimitate: subcenoza Mytilus Actinia equina si subcenoza tipica a midiilor de piatra.
a) Subcenoza Mytilus galloprovincialis Actinia equina. Suprafata ocupata de aceasta subcenoza corespunde in spatiu formei de relief de platforma neregulata cu crapaturi, presarata cu bolovanis cu aspect morenaic, de pana in 5 m adancime. Din cauza adancimilor relativ mici actiunea mecanica a valurilor, fenomenele de resedimentare a particulelor si actiunea inghetului se resimt puternic in aceasta subcenoza, determinand un aspect eterogen al populatiilor de Mytilus galloprovincialis de aici, care sunt compuse din indivizi de marimi foarte diferite. Cealalta specie lider, Actinia equina, domina de regula suprafetele laterale si inferioare ale blocurilor de piatra, nepopulate de midii. Al treilea element cel mai comun al acestei subcenoze este ciripedul Balanus improvisus. Din cauza unui hidrodinamism puternic in aceasta subcenoza exista numeroase specii constructoare de tuburi, cum ar fi amfipodele Corophium bonelli, C. crassicorne, Erichthonius difformis, E. brasiliensis, Amphithoe vaillanti si Jassa ocia, tanaidul tubicol Tanais cavolinii si polichetul perforant Polydora websteri, care sfredeleste in pietrele calcaroase galerii in forma literei U. Un alt polichet tubicol, Fabricia stellaris stellaris, populeaza suprafetele stancilor colmatate cu nisip sau detritus. Polichetul calcros Janua pagenstecheri se intalneste in numar mare pe suprafata algelor macrofite si cea a cochiliilor de midii.
Dintre speciile vagile identificate in aceasta subcenoza mentionam turbelariatul Stylochus tauricus, nemertienii Zygonemertes maslowskyi, Tetrastemma glandulidorsum, Cephalothrix rufifrons si Emplectonema gracile, polichetele Grubeosyllis clavata, Nereis zonata, Platynereis dumerilii, Syllis gracilis, molustele Middendorfia caprearum, Hydrobia pontieuxini, Doridella obscura, amfipodele netubicole Stenothoe monoculoides, Microdeutopus gryllotalpa si Melita palmata, izopodele Jaera nordmanni, Synisoma capito, Idotea baltica si Sphaeroma pulchellum. Creveta-de-piatra (Palaemon elegans) formeaza populatii numeroase pe partile laterale ale bolovanilor si stancilor. Crabii Xantho poressa, Rhithropanopeus harrisi tridentata, Eriphia verrucosa si Pilumnus hirtellus se adapostesc sub pietre si in crapaturile acestora. Meiobentosul este bogat reprezentat prin hidracarieni, oligochete, ostracode, nematode, cumacee, tanaide etc.
b) Subcenoza tipica a midiilor de piatra caracterizeaza fasia sublitoralului pietros, mai jos de adancimea de 5-6 m. In partea sa superioara este limitata de pragul stancos al faliei paralele cu coasta, iar in partea dinspre larg de marginea substratului pietros. Datorita caracterului mai ferm al substratului si conditiilor hidrologice mai putin dinamice, subcenoza se diferentiaza net prin aspectul omogen al populatiilor de midii. Si in aceasta subcenoza specia insotitoare caracteristica este Balanus improvisus. O specie strict localizata in limitele subcenozei tipice a midiilor de piatra este melcul rapitor Rapana thomasiana, care produce ravagii in bancurile de midii. Acest melc, ce a patruns in apele romanesti in 1964, atinge in prezent densitati de pana la 10-15 ex./m2. Tot in aceasta subcenoza sunt intalnite frecvent coloniile albastrui ale spongierului Dysidea fragilis, amfipodele Microdeutopus gryllotalpa, Melita palmata, Jassa ocia, Stenothoe monoculoides, Corophium bonelli, Amphithoe vaillanti si izopodele Jaera nordmanni si Naessa bidentata. Intre filamentele de bisus ale coloniilor de midii se adaposteste o fauna vagila specifica: polichetele Harmothoe impar, Nereiphylla rubiginosa, Neanthes succinea, turbelariatul Stylochus tauricus, gasteropode Setia valvatoides, Hydrobia pontieuxini. In ultima vreme in aceasta subcenoza apare destul de frecvent si scoica Mya arenaria.
Dintre speciile fitofile, caracteristice desisurilor de macrofite, pot fi amintite polichetul Platynereis dumerilii si amfipodul Caprella acanthifera ferox.
Biocenoza malurilor cu Mya si Mytilus. Aceasta biocenoza de tranzitie, care a substituit-o pe cea a lui Spisula subtruncata, se intinde sub forma unei centuri continui intre gurile Dunarii si Capul Caliacra, intre 12 m si 30-40 m adancime, marcand in spatiu limita dintre biocenoza nisipurilor cu Corbula si cea a malurilor cu Mytilus. Sedimentul este reprezentat de maluri, amestecate in partea de nord a litoralului nostru cu nisip fin.
Din cele 15 specii macrobentale identificate in aceasta cenoza, marea majoritate este data de polichete. Dintre acestea, abundentele cele mai ridicate le prezenta speciile oportuniste de polichete Neanthes succinea si Polydora cornuta. Pe langa polichete aici au mai fost semnalate 5 specii de moluste (dintre care dominante sunt Mytilus galloprovincialis si Mya arenaria) si 2 de crustacee (Petranu, 1997).
3.4.6. Biocenoza midiilor de adanc. Este una dintre cele mai caracteristice si bine delimitate biocenoze, atat pentru litoralul romanesc cat si pentru intreg bazinul Marii Negre (Marinov, 1977). In dreptul coastelor romanesti aceasta biocenoza ocupa o zona vasta cuprinsa intre 25-45 m si 48-70 m adancime, dar se prezinta in forma sa tipica mai ales intre 50 si 65 m adancime. Ea ocupa totdeauna un spatiu cu o diferenta de nivel pe verticala de cel putin 20 m, latimea fasiei de fund ocupata variind intre minimum 8 km (la sud-est de Sf. Gheorghe) si maximum 100 km (la est de Sulina). Suprafata totala ocupata, in fata coastelor romanesti, cuprinde o suprafata de circa 7000 km2 (Bacescu et al., 1971).
Substratul este alcatuit din maluri cenusii cu o proportie variabila de scradis. O alta caracteristica a acestei biocenoze o reprezinta stabilitatea mai mare a factorilor de mediu (salinitate, temperatura, curenti).
Din cauza malului aleuritic foarte putin consistent populatiile formei conducatoare - Mytilus galloprovincialis - au o distributie neuniforma, formand mici aglomerari, asa-zisele cuiburi de midii. Dintre formele sesile, care se fixeaza pe suprafata midiilor, caracteristice acestei biocenoze sunt antozoarul Actinothoe clavata, polichetul tubicol Pomatoceros triqueter, urocordatele Ascidiella aspersa si Ciona intestinalis. In mal traiesc numeroase polichete ca Melinna palmata, Nephtys hombergii, Terebellides stroemii, Pectinaria koreni, molustele Mya arenaria, Abra alba, Spisula subtruncata, Polititapes aurea, Acanthocardia paucicostata, Hinia reticulata, amfipodul Ampelisca diadema si castravetele-de-mare Leptosynapta inhaerens. In cuiburile de midii reprezentative sunt nemertianul Cerebratulus ventrosulcatus, polichetele Phyllodoce maculata, Nereiphylla rubiginosa si Harmothoe impar, amfipodele Microdeutopus damnoniensis, M. anomalus si Coremapus versiculatus, cumaceele Cumella pygmaea euxinica si Eudorella truncatula, misidul Pseudoparamysis pontica si crabul Macropipus arcuatus. In timp ce in alcatuirea biomasei totale ponderea cea mai mare o au molustele, in ceea ce priveste densitatea primul loc il detin polichetele. Biomasa medie generla a biocenozei midiilor de adanc este de 416,1 g/m2, din care peste 75% revin midiei (Bacescu et al., 1971).
Datorita variatiei calitative a substratului in cadrul acestei biocenoze exista deosebiri locale insemnate in ceea ce priveste compozitia calitativa si cantitativa a speciilor ce se instaleaza, fapt care ne permite sa distingem la litoralul nostru entitati bionomice cu rang inferior, printre care cele mai importante sunt:
a) Subcenoza Mytilus Lithothamnion Phyllophora. In fata coastelor romanesti aceasta subcenoza ocupa spatiul aflat la est de meridianul de 30s si la nord de paralela de 45s, patrunzand in adancime pana la 45-48 m. Substratul este caracterizat prin dezvoltarea masiva a algelor calcaroase rosii din genul Lithothamnion, determinand o natura mai dura a substratului. Rarele taluri ale algei rosii Phyllophora se pot fixa pe un astfel de substrat, iar specia dominanta a bentosului, Mytilus galloprovincialis, prezinta populatii mai uniform distribuite. Biomasa medie generala a bentosului se ridica la aproape 1.695,2 g/m2, din care 89% este data de midii. Biodiversitatea acestei subcenoze este mai mare decat in cea tipica. Pe langa speciile caracteristice subcenozei tipice, aici mai pot fi intalnite melcul Calyptraea chinensis, crustaceele Phtisica marina si Caprella acanthifera, spongierii din genul Haliclona.
b) Subcenoza Mytilus Modiolus phaseolinus face tranzitia de la malurile cenusii cu Mytilus la cele albastrui cu Modiolus.
In afara celor doua populatii de moluste conducatoare, Mytilus galloprovincialis si Modiolus phaseolinus, in aceasta subcenoza se intalnesc nemertianul Micrura fasciolata, polichetele Nephtys hombergii, Prionospio cirrifera, Sphaerosyllis bulbosa si Protodrilus flavocapitatus si melcul Trophonopsis breviata.
c) malurile cu Melinna palmata. Acumularea detritusului in sedimente din ultima vreme a permis dezvoltarea masiva a populatiilor polichetului Melinna palmata (Losovskaya, 1977a; Gomoiu, 1982). Astfel, se poate vorbi despre aparitia la litoralul nostru a unei asociatii noi, dezvoltate in cadrul suprafetelor ocupate de subcenoza tipica a lui Mytilus, in care specia dominanta este acest polichet iliofil. Aceasta asociatie, cunoscuta sub numele de mal cu Melinna, a fost descrisa din sectoarele invecinate platformei continentale romanesti (golful Odessa si golful Varna), avand statut de biocenoza (Marinov, 1977).
In dreptul coastelor romanesti malurile cu Melinna au maximul de dezvoltare in zona Chituc - Capul Midia, intre 20 si 40 m adancime, unde ocupa o suprafata de peste 1.100 km2 (Gomoiu, 1982; Tiganus, 1982a). Substratul este alcatuit din maluri aluvionare fine, sarace in scradis.
Populatiile de Melinna palmata de aici au o densitate medie de 2.331 ex./m2 si o biomasa de 73,2 g/m2, insa valorile maxime pot depasi 17.700 ex./m2 si 570 g/m2 (Gomoiu, 1982). Desi Melinna palmata reprezinta in mod constant peste 90% din abundenta totala a macrobentosului, biomasele sunt dominate cu peste 50% de Mya arenaria si Mytilus galloprovincialis.
Dintre celelalte grupuri de organisme macrobentale o densitate mai mare o au unele bivalvele Cerastoderma edule, Polititapes aurea, Spisula subtruncata, Abra alba, Chione gallina, Hinia reticulata, polichetele Nephtys hombergii, Pectinaria koreni, Glycera alba, Aricidea claudiae, Capitella capitata si Heteromastus filiformis, crustaceele Ampelisca diadema si Upogebia pusilla si antozoarul Actinothoe clavata.
Biocenoza faseolinelor. Vasta biocenoza a lui Modiolus phaseolinus este o alta biocenoza foarte bine individualizata si specifica Marii Negre, care ocupa numai in fata coastelor romanesti o intindere de aproximativ 10.000 km2, ceea ce ar reprezenta 40% din suprafata totala a platformei continentale (Bacescu et al., 1971). Biocenoza faseolinelor este raspandita intre 50-70 m si 120-130 m, aspectul tipic al biocenozei prezentandu-se intre 58-90 m adancime. Substratul este alcatuit spre tarm din maluri aleuritico-argiloase albastrui, iar spre larg din maluri calcaroase, albe.
Specia cea mai caracteristica a biocenozei este mica scoicuta paroasa cat un bob de fasole Modiolus phaseolinus, cu o densitate medie de 226 ex./m2 si o biomasa de 10 g/m2. Cercetarile efectuate in cadrul acestei biocenoze indica o abundenta medie totala de 1957 ex./m2 si o biomasa generala de 45 g/m2. In aceasta biocenoza au fost identificate 33 de specii de organisme macrozoobentice, dintre care 10 de polichete, 4 de moluste si 10 de crustacee (Dumitrache, 1996-1997).
Printre speciile insotitoare cele mai comune se numara polichetul Terebellides stroemii si anizopodul Apseudopsis ostroumovi.
Alte vietuitoare mai des intalnite in malul faseolinoid sunt: echinodermele Amphiura stepanovi si Leptosynapta inhaerens, foronidul Phoronis euxinicola, celenteratul Pachicerianthus solitarius, tunicierii Ascidiella aspersa si Ciona intestinalis, amfipodele Ampelisca diadema, Microdeutopus damnoniensis, Phtisica marina, Corophium runcicorne si Coremapus versiculatus, molustele Abra alba si Trophonopsis breviata, spongierul calcaros Sycon ciliatum, nemertienii Micrura fasciolata si Tetrastemma coronatum, polichetele Phyllodoce maculata, Exogone naidina, Nephtys hombergii, Polydora cornuta, Notomastus profundus si Protodrillus flavocapitatus, cumaceul Eudorella truncatula si pantopodul Callipallene phantoma (Gomoiu, 1976a; Gomoiu si Tiganis, 1977).
Dintre formele meiobentale care prezinta in aceasta biocenoza abundente ridicate se numara nematodele, copepodele, ostracodele, kinorinchii, halacaridele si turbelariatele.
In braul ce margineste platforma continentala, situat la adancimi cuprinse intre 100-200 m, denumit de Bacescu (1963) etaj periazoic, iar de Sergeeva si Zaika (2000) strat limitrof al bentalului, au fost identificati 92 de taxoni, dintre care 50 apartin meiobentosului si 40 macrobentosului.
Aici substratul este alcatuit din maluri calcaroase compacte, cenusii, uneori cu pete negre datorate unor sedimente mai reduse. Aceste maluri sunt amestecate cu scradis subfosil marunt de cochilii de Modiolus phaseolinus si de moluste ale complexului pontocaspic, Dreissena in special. La suprafata aceste maluri compacte prezinta o patura maronie laxa, de 1-2 cm grosime, rezultata din depunerea ploii de cadavre.
Adancimile cuprinse intre 90 si 160 m se caracterizeaza prin prezenta unei asociatii atipice cu Modiolus phaseolinus in care apar putine exemplare si numai juvenile, alaturi de care apar Terebellides stroemii, Apseudopsis ostroumovi, Cerianthus vestitus, Callipallene phantoma, Halacarus basteri, Peloscolex svirenkoi, Rhizodrilus ponticus, Syllides sp. si Sphaerosyllis bulbosa. Polichetele Vigtorniella zaikai si Protodrilus sp. I au fost gasite deocamdata numai la aceste adancimi (Sergeeva si Zaika, 2000).
Intre 150 si 200 m adancime bentosul este reprezentat aproape exclusiv de forme meiobentale ca nematodele (Enoplus euxinus, Sabatiera clavicauda, Tricoma sp., Desmoscolex sp., Oxystomina sp.) si foraminiferele (Lagena laevis, Ammonia lepida, Nonion depressulum). Singurii reprezentanti ai macrobentosului sunt hidrozoarul Bougainvillia cf. ramosa, bivalva Abra alba si tunicatul Ctenicella amesophleba. Deoarece Bougainvillia are aici o abundenta maxima, Bacescu et al. (1971) propune pentru aceasta centura inferioara, ce in fata coastelor romanesti ocupa o suprafata de aproximativ 700 km2, termenul de biocenoza a malurilor albe cu Bougainvillia.
Conform ultimelor date (Sergeeva, 2000) bentalul Marii Negre este populat si la adancimi mai mari de 200 m. Fauna bentala de aici este reprezentata fie de specii ajunse aici de pe platforma continentala fie de specii necunoscute pana acum pentru Marea Neagra sau chiar noi pentru stiinta. Astfel, intre 470-2250 m adancime au fost identificati aproximativ 60 de taxoni, apartenenta taxonomica a multora dintre acestia fiind inca necunoscuta. In cea mai mare parte aceste organisme apartin meiobentosului (foraminifere, nematode, kinorinchi, harpacticoide, anizopode, ostracode si halacaride), macrobentosul fiind reprezentat de unele amfipode (Gammaridea g. sp. si Caprellidea g. sp.). Un fapt curios este prezenta, chiar si la 2250 m adancime, a cianobacteriilor si algelor brune (Ectocarpus sp.). Densitatea totala a bentosului profund oscileaza intre 1-2 pana la cateva zeci de mii de indivizi pe metru patrat (!).
4. REZULTATE SI DISCUTII
4.1. Conspectul sistematic al speciilor de polichete identificate in cercetarile efectuate
In urma cercetarilor intreprinse au fost analizate 4805 exemplare de polichete apartinand la 23 de specii (Tab. 1), ceea ce ar reprezenta aproximativ 30% din totalul speciilor de polichete semnalate de-a lungul timpului de catre diversi autori in dreptul litoralului romanesc al Marii Negre. Acest numar mic al speciilor de polichete se datoreaza in principal faptului ca probele noastre au fost prelevate de la adancimi mai mici de 20 m, rezumandu-se in esenta la speciile de polichete ce populeaza fundurile pietroase si cele nisipoase. Peste aceasta adancime se intalnesc speciile complexului iliofil. Pe de alta parte, o serie de specii intalnite in trecut la litoralul romanesc al Marii Negre au devenit extrem de rare sau au disparut complet ca urmare a modificarilor induse de om (vezi paragraful 4.6).
Tabelul 1. Compozitia specifica a faunei de polichete din statiile cercetate (MD = Cap Midia; MM = Mamaia; SG = Cap Singol; CT = Constanta; AG = Agigea; EF = Eforie Nord; TZ = Tuzla; MG = Mangalia; VV = Vama Veche).
|
Specia Profilul |
MD |
MM |
SG |
CT |
AG |
EF |
TZ |
MG |
VV |
|
Harmothoe imbricata (Linné, 1767) | |||||||||
|
Harmothoe impar (Johnston, 1839) | |||||||||
|
Nereiphylla rubiginosa (Saint-Joseph, 1888) | |||||||||
|
Eteone picta Quatrefages, 1866 | |||||||||
|
Syllis gracilis Grube, 1850 | |||||||||
|
Typosyllis hyalina (Grube, 1863) | |||||||||
|
Grubeosyllis clavata (Claparède, 1863) |
| ||||||||
|
Nereis zonata Malmgren, 1867 | |||||||||
|
Neanthes succinea (Frey & Leuckart, 1847) | |||||||||
|
Hediste diversicolor (O.F. Müller, 1776) | |||||||||
|
Perinereis cultrifera (Grube, 1840) | |||||||||
|
Platynereis dumerilii (Aud. & M.-Edwards, 1833) | |||||||||
|
Nephtys hombergii Savigny, 1818 | |||||||||
|
Micronephthys stammeri (Ehlers, 1887) | |||||||||
|
Polydora cornuta Bosc, 1802 | |||||||||
|
Spio decoratus Bobretzky, 1871 | |||||||||
|
Prionospio cirrifera Wirén, 1883 | |||||||||
|
Capitella 'capitata' (Fabricius, 1780) | |||||||||
|
Capitella minima Langerhans, 1880 | |||||||||
|
Heteromastus filiformis (Claparède, 1864) | |||||||||
|
Pectinaria koreni (Malmgren, 1866) | |||||||||
|
Melinna palmata Grube, 1870 | |||||||||
|
Fabricia stellaris (Müller, 1774) |
4.2. Structura si distributia faunei de polichete din zonele studiate
Pe substratul nisipos fin, bine tasat, cu mult detritus de macrofite (in special de Enteromorpha) din dreptul Capului Midia dominanta este specia tipic psamobionta Spio decoratus cu o abundenta numerica medie de 1480 ex./m2, insotita de speciile ce prefera nisipul-malos ca Heteromastus filiformis, Neanthes succinea, Polydora cornuta si Capitella minima (Tab. 2) La adancimi mai mari, unde proportia de mal este si mai insemnata, apar speciile Harmothoe imbricata si Prionospio cirrifera. De asemenea, se poate observa ca abundenta numerica a speciilor Neanthes succinea si Heteromastus filiformis creste cu adancimea.
La Mamaia conditiile de mediu sunt oarecum asemanatoare cu cele de la Capul Midia, cu exceptia faptului ca nisipul fin contine o proportie mai ridicata de mal. Acest fapt se datoreaza impiedicarii circulatiei apei din cauza structurilor cu rol de sparge-val din larg, ceea ce permite sedimentarea particulelor fine mai aproape de coasta. Asa se explica aici prezenta speciei iliofile Nephtys hombergii chiar la o adancime de 10 m (Tab. 3). Dintre speciile caracteristice substratului nisipos aici au fost prezente Spio decoratus si Hediste diversicolor, iar dintre cele care se intalnesc preferential pe substrat malos Neanthes succinea, Prionospio cirrifera, Capitella capitata si Heteromastus filiformis.
Tabelul 2. Abundenta numerica a polichetelor (ex./m2) din dreptul Capului Midia.
|
Specia Adancimea |
5 m |
10 m |
20 m |
|
Harmothoe imbricata | |||
|
Nereiphylla rubiginosa | |||
|
Neanthes succinea | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Prionospio cirrifera | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Total |
Tabelul 3. Abundenta numerica a polichetelor (ex./m2) de la Mamaia.
|
Specia Adancimea |
5 m |
10 m |
20 m |
|
Neanthes succinea | |||
|
Hediste diversicolor | |||
|
Nephtys hombergii | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Prionospio cirrifera | |||
|
Capitella capitata | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Total |
Tabelul 4. Abundenta numerica a polichetelor (ex./m2) din dreptul Capului Singol.
|
Specia Adancimea |
10 m |
20 m |
|
Harmothoe impar | ||
|
Grubeosyllis clavata | ||
|
Neanthes succinea | ||
|
Platynereis dumerilii | ||
|
Polydora cornuta | ||
|
Spio decoratus | ||
|
Capitella minima | ||
|
Heteromastus filiformis | ||
|
Total |
In dreptul Capului Singol (Constanta I.N.C.D.M.) substratul este pietros cu nisip fin si mal acumulat in spatiile dintre coloniile de midii (la 10 m adancime) si nisipos fin (la 20 m adancime). Aici au fost gasite doar 8 specii de polichete. Si aici specia dominanta este Spio decoratus, insotita de Polydora cornuta, Neanthes succinea si Heteromastus filiformis (Tab. 4). In zonele cu substrat pietros apar Platynereis dumerilii, Grubeosyllis clavata si Hatrmothoe impar.
La Constanta, in dreptul plajei Modern, in nisipul fin cu mal abunda Spio decoratus (cu pana la 1960 ex./m2) si Heteromastus filiformis (pana la 280 ex./m2). In probele de pe substrat pietros cu nisip fin si mal depus intre interstitiile coloniilor de midii (5 m adancime) apare Grubeosyllis clavata care este o specie caracteristica substratului pietros. La 20 m adancime, unde proportia de mal din sediment devine importanta, apar si speciile iliofile ca Micronephthys stammeri si Melinna palmata (Tab. 5).
Tabelul 5. Abundenta numerica a polichetelor (ex./m2) din dreptul Constantei.
|
Specia Adancimea |
5 m |
10 m |
20 m |
|
Grubeosyllis clavata | |||
|
Neanthes succinea | |||
|
Hediste diversicolor | |||
|
Platynereis dumerilii | |||
|
Micronephthys stammeri | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Prionospio cirrifera | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Melinna palmata | |||
|
Total |
In zona maritima a canalului Dunare Marea Neagra si in acvatoriul portului Constanta Sud (Agigea) are loc ingemanarea apelor dulci ale canalului cu cele marine, fapt pentru care salinitatea de aici sufera variatii largi si bruste. Aportul mare de ape bogate in substante biogene determina aici fenomene de inflorire a apei frecvente, ceea ce atrage dupa sine instalarea hipoxiei si chiar anoxiei in patura de apa de contact, cauzand mortalitatea in masa a organismelor bentonice. In urma acestor procese, peste substratul artificial alcatuit din blocuri de piatra si din beton, s-a acumulat in timp un strat de mal detritic negru de 20-40 cm grosime in amestec cu cochilii goale de midii si cu un miros puternic de hidrogen sulfurat. Singurele polichete prezente aici sunt Neanthes succinea (A = 235 ex./m2 si B = 2,346 g ex./m2) si Polydora cornuta (A = 489 ex./m2 si B = 0,5 g/m2), specii cu pronuntate valente oportuniste, ce se dezvolta in numar mare in conditiile simplificarii structurii cenotice a comunitatilor (Surugiu, 2000; Surugiu et al., 2004). Acest fapt ne permite sa le incadram in categoria speciilor indicatoare de poluare cu materii organice si de eutrofizare.
In afara radei portului, ceva mai la sud de punctul pescaresc Agigea, substratul este pietros, sub forma unei placi calcaroase cu panta lina, care pe la 4-5 m adancime prezinta o denivelare brusca (asa-zisul salac), dupa care coboara din nou lin. Intre 1 si 5 m adancime fundurile pietrose sunt intercalate de petice de sedimente nisipoase cat si de sedimente marnos-argiloase. Odata cu cresterea adancimii proportia de mal creste, astfel incat pe la 15-18 m adancime substratul devine malos. Fauna de polichete de pe fund pietros este dominata numeric de formele mici precum Polydora cornuta (densitatea medie 2986 ex./m2) si Grubeosyllis clavata (abundenta medie 1303 ex./m2). In schimb, la realizarea biomasei totale contributia cea mai mare o au nereididele de talie mare Platynereis dumerilii (abundenta medie de 922 ex./m2 si biomasa medie de 20,416 g/m2) si Neanthes succinea (abundenta medie de 547 ex./m2 si biomasa medie de 18,904 g/m2). Acestora li se mai adauga Nereis zonata, Nereiphylla rubiginosa, Syllis gracilis si Typosyllis hyalina (Tab. 6). Pe fundurile acoperite de marne argiloase pot fi gasite speciile Polydora cornuta (unde realizeaza densitati de pana la 110.000 ex./m2) si Neanthes succinea. Pe substrat malos, cenusiu-negricios, cu miros slab de hidrogen sulfurat si cu scradis de Mytilus galloprovincialis de la adancimi mai mari de 15 m adancime sunt prezente speciile Nephtys hombergii (48 ex./m2), Melinna palmata (5 ex./m2), Pectinaria koreni, Prionospio cirrifera si Harmothoe impar.
Din tabelul 6 se poate observa ca, in linii generale, densitatea numerica a speciei Harmothoe impar creste odata cu cresterea adancimii, indicand preferinta acesteia pentru substratul pietros puternic colmatat cu mal. In schimb efectivele speciei Grubeosyllis clavata descresc odata cu cresterea adancimii, ceea ce arata afinitatea acesteia pentru substratul pietros necolmatat. Polichetele Polydora cornuta si Neanthes succinea par a fi indiferente fata de natura substratului, iar pentru specia Platynereis dumerilii se poate observa o descrestere a abundentei numerice cu adancimea, prezentand un maxim la adancimea de 4 m. Aceste date concorda bine cu biologia speciei, care este o specie asociata algelor macrofite. Abundenta numerica maxima a polichetelor se inregistreaza la 4 m adancime,fiind de 12.350 ex./m2. In schimb biomasa maxima totala a polichetelor (26,085 g/m2) a fost gasita la 14 m adancime.
Tabelul 6. Abundenta numerica a polichetelor (ex./m2) de la Agigea in functie de adancime.
|
Proba |
AG 9 |
AG 8 |
AG 7 |
AG 6 |
AG 5 |
AG 4 |
AG 3 |
AG 2 |
AG 1 |
|
Specia Adanc. |
0 m |
2 m |
4 m |
6 m |
8 m |
10 m |
12 m |
14 m |
16 m |
|
Harmothoe cf. impar | |||||||||
|
Harmothoe imbricata | |||||||||
|
Nereiphylla rubiginosa | |||||||||
|
Syllis gracilis | |||||||||
|
Typosyllis hyalina | |||||||||
|
Grubeosyllis clavata | |||||||||
|
Nereis zonata | |||||||||
|
Neanthes succinea | |||||||||
|
Platynereis dumerilii | |||||||||
|
Polydora cornuta | |||||||||
|
Prionospio cirrifera | |||||||||
|
Total |
Tabelul 7. Biomasele realizate de catre polichete (g/m2) la profilul Agigea in functie de adancime.
|
Proba |
AG 9 |
AG 8 |
AG 7 |
AG 6 |
AG 5 |
AG 4 |
AG 3 |
AG 2 |
AG 1 |
|
Specia Adanc. |
0 m |
2 m |
4 m |
6 m |
8 m |
10 m |
12 m |
14 m |
16 m |
|
Harmothoe cf. impar | |||||||||
|
Harmothoe imbricata | |||||||||
|
Nereiphylla rubiginosa | |||||||||
|
Syllis gracilis | |||||||||
|
Typosyllis hyalina | |||||||||
|
Grubeosyllis clavata | |||||||||
|
Nereis zonata | |||||||||
|
Neanthes succinea | |||||||||
|
Platynereis dumerilii | |||||||||
|
Polydora cornuta | |||||||||
|
Prionospio cirrifera | |||||||||
|
Total |
Comparand probele prelevate la Agigea de la 17 m adancime de pe substrat dur natural, alcatuit din placi calcaroase puternic colmatate cu mal, si de pe substrat artificial, format din ingramadiri de stabilopozi ce intra in alcatuirea Digului de Sud se pot constata unele diferente (Tab. 8). In primul rand, neregularitatile dintre stabilopozi ofera mai multe microhabitate fata de placile calcaroase netede si mai mult sau mai putin orizontale. In plus, prezenta suprafetelor verticale in cazul substratului artificial impiedica acumularea malului. Asa se explica prezenta pe substrat natural a speciilor iliobionte (Nephtys hombergii, Melinna palmata si Pectinaria koreni) si iliofile (Harmothoe imbricata, Heteromastus filiformis si Prionospio cirrifera). Desi substratul natural de aici are o aparenta mai degraba de substrat malos, curios este faptul ca aici se intalnesc si specii tipic petricole ca Nereiphylla rubiginosa, Syllis gracilis si Grubeosyllis clavata. De asemenea, se poate observa din tabelele 8 si 9 ca abundentele numerice si biomasele sunt mai ridicate in cazul substratului pietros natural comparativ cu cel artificial.
Tabelul 8. Abundenta numerica a polichetelor (ex./m2) de la Agigea de la 17 m adancime de pe substrat artificial (P01* si P02*) si natural (P03** si P04**).
|
Specia Proba |
P01* |
P02* |
P03** |
P04** |
|
Harmothoe imbricata | ||||
|
Harmothoe impar | ||||
|
Nereiphylla rubiginosa | ||||
|
Syllis gracilis | ||||
|
Grubeosyllis clavata | ||||
|
Syllidae indet. | ||||
|
Neanthes succinea | ||||
|
Polydora cornuta | ||||
|
Prionospio cirrifera | ||||
|
Capitella minima | ||||
|
Heteromastus filiformis | ||||
|
Pectinaria koreni | ||||
|
Melinna palmata | ||||
|
Polychaeta varia | ||||
|
Total |
Tabelul 9. Biomasa polichetelor (g/m2) de la Agigea (17 m adancime) de pe substrat artificial (P01 si P02) si natural (P03 si P04).
|
Specia Proba |
P01* |
P02* |
P03** |
P04** |
|
Harmothoe imbricata | ||||
|
Harmothoe impar | ||||
|
Nereiphylla rubiginosa | ||||
|
Syllis gracilis | ||||
|
Grubeosyllis clavata | ||||
|
Syllidae indet. | ||||
|
Neanthes succinea | ||||
|
Polydora cornuta | ||||
|
Prionospio cirrifera | ||||
|
Capitella minima | ||||
|
Heteromastus filiformis | ||||
|
Pectinaria koreni | ||||
|
Melinna palmata | ||||
|
Polychaeta varia | ||||
|
Total |
La Eforie Nord substratul este alcatuit din nisip mediu si fin. Aici au fost identificate 9 specii de polichete (Tab. 10). Populatiile de polichete de aici prezinta o structura asemanatoare cu cea a nisipurilor de la nord de Constanta. Astfel, specia lider de la Eforie Nord este Spio decoratus cu o abubdenta numerica de pana la 3248 ex./m2, urmata de Polydora cornuta cu o densitate pana la 160 ex./m2. In plus fata de polichetele ce populeaza substratul nisipos din partea nordica a litoralului romanesc, in zonele cu nisip malos apare specia Eteone picta. La adancimi mai mari, unde nisipul malos este inlocuit de mal domina polichetele iliofile ca Melinna palmata, Nephtys hombergii si Micronephthys stammeri. Abundenta numerica totala a polichetelor de la Eforie Nord descreste pe masura cresterii adancimii, din cauza diminuarii efectivelor speciei Spio decoratus.
Tabelul 10. Abundenta numerica a polichetelor (ex./m2) de la Eforie Nord.
|
Specia Adancimea |
5 m |
10 m |
20 m |
|
Eteone picta | |||
|
Neanthes succinea | |||
|
Nephtys hombergii | |||
|
Micronephthys stammeri | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Melinna palmata | |||
|
Total |
La Capul Tuzla pe fundul nisipos in amestec cu scradis au fost identificate 14 specii de polichete (Tab. 11). Dintre speciile dominante pot fi citate Spio decoratus (cu pana la 1140 ex./m2), Prionospio cirrifera (pana la 500 ex./m2) si Capitella minima (380 ex./m2). Pe substrat pietros la 5 m adancime au fost gasite speciile Neanthes succinea, Platynereis dumerilii, Nereis zonata, Nereiphylla rubiginosa si Harmothoe impar. La adancimi mai mari incep sa apara formele iubitoare de mal ca Nephtys hombergii si Melinna palmata. Diversitatea specifica mare si prezenta unor specii rare ca Nereiphylla rubiginoss si Nereis zonata indica ape cu o incarcatura slaba a substantelor biogene.
Tabelul 11. Abundenta numerica a polichetelor (ex./m2) de la Capul Tuzla.
|
Specia Adancimea |
5 m |
10 m |
20 m |
|
Harmothoe impar | |||
|
Nereihpylla rubiginosa | |||
|
Eteone picta | |||
|
Nereis zonata | |||
|
Neanthes succinea | |||
|
Platynereis dumerilii | |||
|
Nephtys hombergii | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Prionospio cirrifera | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Melinna palmata | |||
|
Total |
La Mangalia substratul se prezinta sub forma unui platou calcaros aproape orizontal, presarat cu bolovani mari din loc in loc. Acestia din urma sunt acoperiti aproape in intregime de spongieri. Aici specia cea mai comuna este Fabricia sabella (3480 ex./m2), urmata de Grubeosyllis clavata (1160 ex./m2) si Neanthes succinea (160 ex./m2). Mai putin abundente sunt Capitella minima, Polydora cornuta si Hediste diversicolor.
La Vama Veche, in perimetrul rezervatiei submarine, speciile dominante numeric sunt viermii Grubeosyllis clavata (8500 ex./m2), Polydora cornuta (1231 ex./m2) si Neanthes succinea (1077 ex./m2). Dezvoltarea masiva a populatiilor unor specii sensibile precum Syllis gracilis (cu densitati de pana la 620 ex./m2) si Perinereis cultrifera indica ape curate, nesupuse poluarii organice. Acest lucru este confirmat si de prezenta unor specii rare ca Harmothoe imbricata si Nereiphylla rubiginosa.
Tabelul 12. Abundenta numerica a polichetelor (ex./m2) de la Mangalia si Vama Veche.
|
Specia Adancimea |
Mangalia 0,5 m |
Vama Veche 3 m |
Vama Veche 5 m |
|
Harmothoe imbricata | |||
|
Harmothoe impar | |||
|
Nereihpylla rubiginosa | |||
|
Syllis gracilis | |||
|
Grubeosyllis clavata | |||
|
Neanthes succinea | |||
|
Hediste diversicolor | |||
|
Perinereis cultrifera | |||
|
Platynereis dumerilii | |||
|
Polydora cornuta | |||
|
Spio decoratus | |||
|
Capitella minima | |||
|
Heteromastus filiformis | |||
|
Fabricia stellaris | |||
|
Total |
Tabelul 13. Indicatorii ecologici analitici si sintetici din profilele studiate.
|
Profilul Indicele |
Numarul total de specii (S) |
Abundenta numerica totala (A) |
Indicele de diversitate specifica Shannon-Wiener (H') |
Indicele de echitabilitate Pielou (E) |
|
Cap Midia | ||||
|
Mamaia | ||||
|
Cap Singol |
| |||
|
Constanta | ||||
|
Agigea | ||||
|
Eforie Nord | ||||
|
Tuzla | ||||
|
Mangalia | ||||
|
Vama Veche |
Urmarind diversitatea specifica pe profile se poate observa ca numarul de specii a variat intre 8 la Capul Midia si 16 la Agigea (Tab. 13). In general, in zonele cu substrat predominant nisipos (Cap Midia, Mamaia, Cap Singol, Constanta si Eforie Nord) diversitatea specifica este mai mica decat in cele cu substrat predominant pietros (Agigea, Cap Tuzla, Mangalia si Vama Veche). De asemenea, zonele mai putin influentate de eutrofizare din sudul litoralului prezinta un numar mai mare de specii (Fig. 4). Diversitatea specifica ridicata de la Agigea se explica prin numarul mult mai mare de probe prelevate aici, dar si din cauza curentului ciclonal care aduce ape mai putin poluate din sud (vezi si paragraful 3.3.2.).
Abundenta numerica totala a polichetelor a variat intre 453 ex./m2 la Mamaia si 7050 ex./m2 la Vama Veche (Tab. 13). In linii mari, abundenta numerica este mai ridicata in zonele cu substrat pietros datorita prezentei unui numar mai mare de microhabitate (Fig. 5). Totusi urmarind variatia abundentei numerice in diferite profile cu acelasi tip de substrat se poate observa ca zonele mai putin afectate de poluare prezinta densitati mai mari ale polichetelor. Deoarece directia generala a curentilor este de la nord la sud, punctele situate mai spre sud sunt mai putin influentate de apele Dunarii incarcate cu substante biogene, si deci cu ape mai putin poluate. Aceasta explica cresterea abundentei numerice de la nord la sud. In dreptul profilelor unde exista deversari de ape poluate (combinatul petrochimic de la Navodari si statiile de epurare a apei de la Eforie Sud si Mangalia) se poate observa o descrestere semnificativa a densitatii polichetelor.
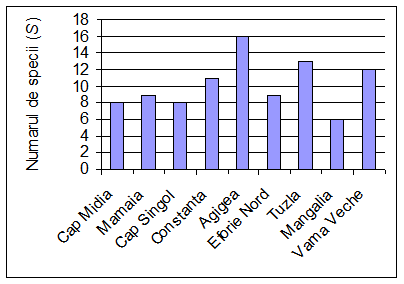
Figura 4. Variatia numarului total de specii in profilele analizate.
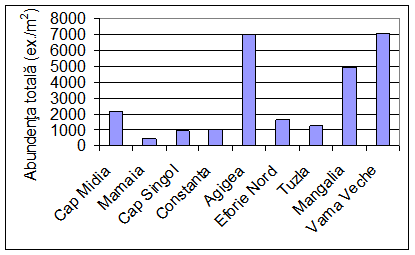
Figura 5. Variatia abundentei totale a polichetelor in profilele analizate.
Indicele de diversitate Shannon-Wiener (H) a variat intre 1,211 la Mamaia si 2,192 la Capul Tuzla, iar echitabilitatea (E) a variat intre 0,470 la Mangalia si 0,760 la Tuzla (Tab. 13). Echitsabilitatea mica de la Mangalia se explica prin dominanta puternica a speciilor Fabricia sabella si Grubeosyllis clavata. Atat indicele de diversitate Shannon cat si echitabilitatea prezinta valori mai mici in zonele supuse eutrofizarii (Mamaia, Eforie Nord si Mangalia) si mai mari in zonele cu ape relativ curate din sudul litoralului (Agigea, Tuzla si Vama Veche) (Fig. 6 si 7). Comparand datele noastre cu cele din perioada 1998-1999 (Surugiu, 2002) se poate constata o crestere a valorilor indicelui de diversitate si a echitabilitatii, ceea ce denota o scadere a nivelului de eutrofizare si de poluare organica (Tab. 14).
Astfel, la Constanta, la Tuzla si la Vama Veche s-au inregistrat valori mai mari atat pentru indicele lui Shannon cat si pentru echitabilitate. La Agigea si la Eforie Nord, desi valoarea indicelui Shannon s-a diminuat, echitabilitatea a crescut, ceea ce indica diminuarea efectivelor speciilor oportuniste, tolerante la poluare. Doar la Mamaia si la Mangalia s-au inregistrat valori mai mici decat in anii 1998-1999.
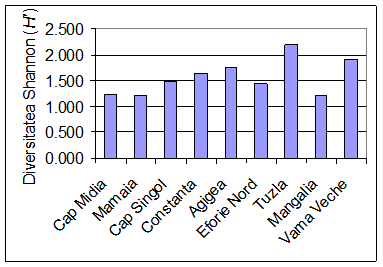
Figura 6. Variatia indicelui de diversitate Shannon in profilele analizate.
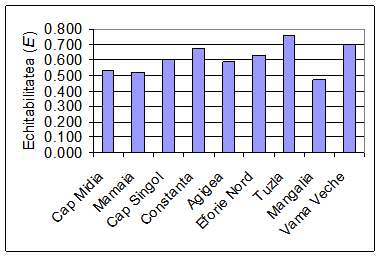
Figura 7. Variatia echitabilitatii in profilele analizate.
Tabelul 14. Compatie intre indicii de diversitate.
|
Profilul | ||||
|
H |
E |
H |
E |
|
|
Mamaia | ||||
|
Constanta | ||||
|
Agigea | ||||
|
Eforie Nord | ||||
|
Tuzla | ||||
|
Mangalia | ||||
|
Vama Veche | ||||
4.3. Caracteristicile ecologice ale unor specii de polichete
In cele ce urmeaza sunt prezentate caracteristicile ecologice ale speciilor care au preferinte bine definite fata de conditiile de mediu si care, in acest fel, pot fi utilizate ca bioindicatori ai eutrofizarii sau poluarii cu substante organice. Semnificatia lor ecologica rezida din faptul ca prezenta sau abundenta lor poate indica un anumit tip de substrat, de biocenoza sau existenta unor perturbari ale conditiilor de mediu.
Harmothoe imbricata (Linné, 1767), specie rara in trecut in partea de nord-vest a Marii Negre, in prezent, odata cu cresterea nivelului de eutrofizare, a devenit o specie comuna. In probele noastre H. imbricata a avut o frecventa de 15,2% si a aparut in cazul substratului pietros cu o abundenta maxima de 178 ex./m2 si o biomasa maxima de 1,920 g/m2. Jakubova si Malm (1930) arata ca H. imbricata poate rezista in conditii de anoxie timp de 7 zile si la prezenta hidrogenului sulfurat timp de 3-6 zile. Raspunde in mod pozitiv la cresterea nivelului de eutrofizare.
Harmothoe impar (Johnston, 1839) este o specie mai frecvent intalnita (42,4% in probele noastre), dar mult mai sensibila la prezenta hidrogenului sulfurat decat H. reticulata. Noi am gasit-o in detritusul si malul acumulat in interstitiile coloniilor de midii. Densitatea si biomasa maxima, de 1200 ex./m2 si de respectiv 7,244 g/m2, a fost inregistrata la 17 m adancime la Agigea.
Nereiphylla rubiginosa (Saint-Joseph, 1888) raspunde in mod negativ la cresterea nivelului de eutrofizare si poluare organica, devenind in prezent o specie rara in epibiozele litorale de la 5-10 m adancime, cu o frecventa de 21,2%. Specie sensibila la diminuarea concentratiei de oxigen din apa. A fost gasita la Capul Midia, Agigea, Tuzla si Vama Veche.
Eteone picta Quatrefages, 1866 este o specie indicatoare a perturbarilor naturale sau de origine antropica, deoarece este o forma rezistenta la conditii de anaerobioza si la prezenta hidrogenului sulfurat (Jacubova & Malm, 1930). In probele noastre a aparut cu o frecventa de numai 6%.
Syllis gracilis Grube, 1840 (Fig. 8) si Typosyllis hyalina (Grube, 1863) sunt specii sensibile, prezente numai in apele relativ curate din partea sudica a litoralului romanesc al Marii Negre (la Agigea si in rezervatia marina Vama Veche 2 Mai). Asa se explica raritatea lor in probele noastre, find de 27,3% si respectiv 3%.

Figura 8. Syllis gracilis - specie indicatoare pentru ape curate.
Grubeosyllis clavata (Claparède, 1863) este singura specie dintre silide care pare sa prospere in conditii de eutrofizare si incarcare cu materii organice a apei si sedimentelor marine (frecventa de 48,5%). A fost identificata chiar si in apele politrofe ale golfului Mangalia.
Neanthes succinea (Frey & Leuckart, 1847) (Fig. 9) este o specie euritopa, capabila sa se raspandeasca intens datorita stadiului de dezvoltare pelagic, umpland repede nisele eliberate ca rezultat al mortalitatii in masa a bentosului. Forma rara in trecut, prezenta mai ales in limanuri (Markovski, 1954), in momentul de fata, odata cu amplificarea eutrofizarii, a devenit specia cea mai comuna (cu o frecventa in probe de 75,8%), nu numai la litoralul romanesc ci si in intreg bazinul Marii Negre (Murina si Mikhailova, 1994). Losovskaya (1988) vorbeste chiar de existenta unei biocenoze cu Neanthes succinea. Acest polichet populeaza aproape pe toate tipurile de substrat, dar prefera mai ales sedimentele detritice moi (mal, nisip malos, uneori pe scradis) in care se poate ingropa pana la 10 cm adancime. Populeaza si fouling-ul de pe coca navelor (Dumitrescu, 1962; Surugiu, date nepubl.). In prezent aceasta specie reprezinta 23,3% din abundenta totala si, respectiv, 81,8% din biomasa totala a polichetelor de la litoralul romanesc al Marii Negre (Surugiu, 2004).

Figura 9. Neanthes succinea indicator al eutrofizarii si poluarii cu materii organic.
In apele politrofe sau in cele poluate cu produse petroliere, ca de exemplu in portul Constata Sud Agigea sau in golful Mangalia, acest polichet formeaza populatii bogate si caracteristice impreuna cu Polydora cornuta. In asemenea conditii indivizii de Neanthes succinea ating dimensiuni foarte mari, de pana la 100 mm lungime. Aceasta specie este indicator al perturbarii naturale sau antropice numai in cazul in care atinge abundente, dar mai ales biomase ridicate. De exemplu, daca in conditii de mare deschisa acest polichet prezinta o abundenta medie de 4300 ex./m2 si o biomasa medie de 16,4 g/m2, in apele eutrofizate la o abundenta medie de 2800 ex./m2 biomasele date de acesta se ridica la 378 g/m2 (Surugiu, 2002).
Nereis zonata Malmgren, 1867 poate fi considerata drept o specie sensibila, indicatoare de ape slab eutrofizate. Daca in trecut aceasta specie prezenta in biocenoza midiilor de piatra efective mari, de pana la 2910 ex./m2 (Bacescu et al., 1963), in conditiile actuale de eutrofizare si poluare organica nu prezinta densitati mai mari de 100 ex./m2. Locul acestei specii in biocenoza a fost preluat de catre populatiile de Neanthes succinea, care este o specie mai toleranta la perturbarile conditiilor de mediu. De asemenea, N. zonata este foarte sensibila la conditiile anoxice sau la prezenta hidrogenului sulfurat (Jakubova si Malm, 1930; Losovskaya, 1962). Frecventa acesteia in probe a fost de 12,1%.
Hediste diversicolor (O.F. Müller, 1776) este o specie extrem de euribionta, indicatoare de instabilitate a conditiilor de mediu. Desi H. diversicolor prezinta aceleasi cerinte fata de conditiile de viata ca si N. succinea, unii autori (Rasmussen, 1973; Kisseleva, 1981) atrag atentia asupra faptului ca nu au fost observate populatii mixte ale celor doua specii. In marea majoritate a probelor noastre insa am putut gasi indivizi apartinand ambelor specii, cei de H. diversicolor fiind insa mult mai putin numerosi. Capacitatea de dispersie redusa a acesteia din urma fata de prima a dus la inlocuirea aproape totala a populatiilor de Hediste diversicolor cu cele de Neanthes succinea (Manoleli, 1980). Aceasta specie a devenit rara si in apele deschise (prezenta numai in 9% din probe).
Specia este rezistenta la reducerea cantitatii de oxigen dizolvat si la prezenta H2S in apa si in sediment. Poate supravietui in conditii anaerobe timp de 7 zile si rezista 3 zile la 7,5 cm3 H2S/l apa (Jakubova si Malm, 1930).
Perinereis cultrifera (Grube, 1840) poate fi considerata drept specie indicatoare de ape relativ curate, deoarece a fost gasita de noi doar la Vama Veche in epibiozele pietrelor si stancilor din zona litorala, ceea ce explica faptul ca frecventa acestei specii in probe a fost de numai 6% (Fig. 10).

Figura 10. Perinereis cultrifera, specie specie sensibila, indicatoare de ape nepoluate.
Platynereis dumerilii (Audouin & Milne-Edwards, 1833) este o specie de masa, indicatoare de poluare. Specie fitofila prin excelenta, in trecut era una dintre formele conducatoare in desisurile de Cystoseira barbata, iar in prezent s-a adaptat noilor conditii de viata, fiind foarte abundenta in microbiotopul algelor macrofite de la Agigea unde prezenta densitati de pana la 2925 ex./m2. Specie foarte sensibila la anoxie si la prezenta hidrogenului sulfurat (Losovskaya, 1962). Colonist secundar. Cresterea densitatilor acestei specii comparativ cu cele ale lui Neanthes succinea din ultimii 3 ani denota o ameliorare a calitatii mediului marin, frecventa acestei specii in probe crescand la 39,4%.
Nephtys hombergii Savigny, 1818 populeaza sedimentele maloase si malos-nisipoase cu o frecventa de 15,2%. Este o specie toleranta la poluare organica si la diminuarea concentratiei oxigenului dizolvat in apa.

Figura 11. Polydora cornuta, specie oportunista, indicatoare de eutrofizare.
Spio decoratus Bobretzky, 1871 este o forma tipic psamobionta, caracteristica biocenozei nisipurilor fine cu Corbula mediterranea. Specie toleranta la poluarea cu substante organice. In conditii de sedimentare puternica trece de la modul de hranire suspensivor la cel pelofag, ceea ce explica succesul competitiv al acestei specii in conditiile eutrofizarii si poluarii cu materii organice. In nisipurile fine de la Eforie Nord atinge densitati de 3240 ex./m2. Frecventa acestei specii este de 48,5%.
Polydora cornuta Bosc, 1802 (Fig. 11) prefera apele eutrofe, bogate in detritus si elemente biogene ale estuarelor, fiind toleranta la salinitati reduse (Blake, 1996). In partea nord-vestica a Marii Negre acest polichet a fost gasit la salinitati ce au oscilat intre 6,4 si 17,9 (Losovskaya, 1977b; Surugiu, 2002). In unele cazuri acest polichet a fost gasit in portiunile de fund puternic poluate cu produse petroliere (Losovskaya, 1976).
In dreptul litorlului romanesc Poydora cornuta a suferit o dezvoltare de tip exploziv, astfel ca abundente de 6000-22.000 ex./m2 au devenit obisnuite (Fig. 9). Dezvoltarea in masa a acestei specii, mai ales in zonele supuse influentei apelor dulci, probeaza un proces de eutrofizare a apelor. In prezent este specia cel mai frecvent intalnita, fiind prezenta in 87,9% din probe.
Prionospio cirrifera Wirén, 1883 este o specie rezistenta la poluarea cu substante organice sau cu produse petroliere, fiind mai abundenta in zonele cu poluare moderata. Cresterea efectivelor acestei specii sunt de asemenea indicatoare pentru intensificarea eutrofizarii.
Capitella capitata (Fabricius, 1780) este o specie eurioica, cu largi valente oportuniste. Este caracteristica malurilor negre lichide, cu miros de hidrogen sulfurat, prezentand o toleranta crescuta fata de diminuarea cantitatii de oxigen dizolvat. Potrivit cercetarilor lui Jakubova si Malm (1930) capitelidele din Marea Neagra pot supravietui un timp indelungat in ape cu o concentratie de pana la 20,4 ml H2S /l. Este considerata a fi un indicator al perturbarilor naturale sau antropice, toleranta la poluare organica si industriala (Reish, 1970). Inca Zernov (1913) a atras atentia asupra faptului ca aceasta specie este caracteristica zonelor celor mai impurificate. Potereaev (1936, citat de Milovidova, 1975) o incadreaza pe Capitella capitata in categoria organismelor α-mezosaprobe. Losovskaya (1977b) gaseste efective numeroase ale acestui polichet (pana la 1360 ex./m2) in zona celor mai impurificate plaje ale Odessei. In conditiile eutrofizarii abundenta numerica a acestei specii atinge valori de 4859 ex./m2. In probele noastre speciile genului Capitella au aratat o puternica corelatie foarte puternica intre abundenta numerica si cantitatea de carbon organic din sediment, r2 = 0,94 (Fig. 3). De asemenea, aceasta corelatie este semnificativa din punct de vedere statistic (t = 5,84, 3 g.l., P < 0,01).
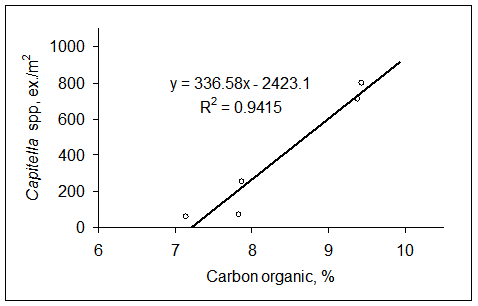
Figura 10. Variatia abundentei numerice a speciilor genului Capitella in functie de continutul de carbon organic din sediment.
Heteromastus filiformis (Claparède, 1864) si Capitella minima Langerhans, 1880 sunt specii euritope, indicatoare pentru perturbari naturale sau induse de om.
Pectinaria koreni (Malmgren, 1866) este caracteristica sedimetelor nisipoase, dar poate tolera mici cantitati de mal. Este o specie care indica nivele intermediare de poluare. Populatiile sale au prezentat cresteri ca raspuns la eutrofizare.
Melinna palmata Grube, 1870 caracterizeaza in mod pozitiv gradul de eutrofizare si poluare a apei si sedimentelor cu substante organice. Specie rezistenta la conditii de anoxie.
Fabricia stellaris stellaris (Müller, 1774) este o specie indicatoare de poluare sau de instabilitate a conditiilor de mediu.

Figura 10. Ficopomatus enigmaticus forma larg dezvoltata in apele poluate ale porturilor.
Ficopomatus enigmaticus (Fauvel, 1923) este o forma toleranta la poluare, care colonizeaza suprafetele submerse ale navelor sau ale constructiilor hidrotehnice. In conditii prielnice este capabila sa formeze chiar pseudorecifi. Pentru a se putea dezvolta necesita ape linistite (in porturi, golfuri, canale, estuare etc.). Abundenta acestui polichet in portul Constanta Sud se cifreaza la 120.000 ex./m 2, iar greutatea tuburilor a putut fi estimata la 6,9 kg/m2 (Surugiu, 2002).
4.4. Eutrofizarea si poluarea cu substante organice
Eutrofizarea apelor costiere a marilor si oceanelor a evoluat rapid in ultimii 30-40 de ani ca rezultat al unor activitati agricole si industriale intensive. Totusi, gradul de eutrofizare pentru fiecare bazin acvatic in parte depinde de anumite particularitati geografice, fizice, chimice si biologice.
Marea Neagra, ca obiect al eutrofizarii antropice, a atras atentia specialistilor inca de la sfarsitul anilor 60. Potrivit lui Odum (1971, citat de Zaitsev, 1991) sursa principala a eutrofizarii este aportul continental de ape sau outwelling-ul. Deoarece in partea de nord-vest a Marii Negre (din care face parte si litoralul romanesc) sunt localizate gurile de varsare ale celor mai mari fluvii din acest bazin Dunarea (cu un debut anual de 208 km3), Nistrul (10,2 km3/an) si Niprul (51,2 km3/an), aici s-a observat cresterea cea mai insemnata a concentratiei substantelor biogene insotita de o dezvoltare extraordinara a abundentei si biomasei planctonice. In prezent concentratia de nitrati din apele costiere romanesti este de cinci ori mai mare fata de cea din anii 60-70, cea a fosfatilor de 25 ori, iar cea a substantei organice de 1,2 ori (Tab. 15).
In acest fel putem aprecia ca in momentul de fata apele de suprafata ale Marii Negre fac parte din categoria apelor mezotrofe si eutrofe, in timp ce in zona de influenta a gurilor de varsare ale Dunarii sau in acvatoriile semiinchise cum ar fi cele ale porturilor Midia, Constanta sau Mangalia apele sunt hipertrofe.
Mareele rosii, care in trecut erau un fenomen foarte rar, in zilele noastre se petrec cu regularitate in fiecare an, uneori chiar de 2-3 ori pe an. In perioada 1983-1988 in apele romanesti au fost inregistrate 24 de infloriri algale monospecifice, determinate de 8 taxoni. Cele mai ridicate densitati numerice de la litoralul romanesc le-au avut algele planctonice Prorocentrum cordatum (pana la 87.600.000 celule/l), Sceletonema costatum (141.400.000 celule/l) si Coccolithus huxley (291.200.000 celule/l). Dintre organismele zooplanctonice ctenoforul Noctiluca scintillans poate atinge o abundenta de 6.800.000.000 ex./m3 si o biomasa de 500 kg/m3, iar ciliatul Mezodinium rubrum o abundenta de 4.600.000 celule/l si o biomasa de 280 g/m3. Cel mai mare numar al ctenoforului Mnemiopsis leidyi, nou-patruns in Marea Neagra din Oceanul Atlantic, se inregistreaza de asemenea in partea de nord-vest a Marii Negre. Abundenta acestei specii in apele costiere depaseste 500 ex./m3.
Tabelul 15. Variatiile multianuale ale concentratiei fosfatilor, azotatilor, silicatilor si substantei organice dizolvate (MOD) in apele costiere din dreptul Constantei.
|
Perioada |
P-PO4 (μg/l) |
N-NO3 (μg/l) |
Si-SiO4 (μg/l) |
Substanta organica (mgO2/l) |
In ultimii ani in mari adesea se dezvolta in numar mare algele peridinee (de ex. Gonioaulax), care pot elimina substante toxice si care apoi nimeresc in lanturile trofice. Aceste toxine ajung prin intermediul nevertebratelor bentonice filtratoare in corpul pestilor si in cele din urma se pot dovedi fatale pentru om.
Incepand cu anul 1973 pe arii largi ale platformei continentale din nord-vestul Marii Negre au inceput sa fie semnalate mortalitati in masa ale organismelor de fund (Zaitsev & Mamaev, 1997; Surugiu, 2002). S-a stabilit ca din cauza hipoxiei intre 100 si 200 t de organisme marine, inclusiv pesti, mor pe fiecare kilometru patrat de fund marin. Intre 1973 si 1990 pierderile de organisme bentonice numarau 60 milioane tone, dintre care 5000 t reveneau pestilor (Zaitsev, 1992). Mortalitatile in masa dispar odata cu instalarea homotermiei de toamna, cand apele din profunzime sunt oxigenate datorita amestecului de ape produs de vant si de modificarea densitatii apei. Aceste zone depopulate incep sa fie recolonizate de catre larvele pelagice aduse de catre curenti din zonele limitrofe sau neafectate de hipoxie. Populatiile bentonice sunt reinnoite si acest proces continua pana la instalarea urmatoarei perioade cu deficit de oxigen si ciclul se repeta.
4.5. Polichetele indicatori ai calitatii mediului
Componenta macrozoobentica este cea mai des utilizata in aprecierea starii de sanatate a mediului marin deoarece organismele bentice:
- sunt forme mai mult sau mai putin sedentare, neputand evita deteriorarea calitatii apei sau sedimentelor;
- prezinta o longevitate relativ mare, indicand o integrare la conditiile de calitate a apei si sedimentului;
- apartin la diferite categorii sistematice, adica prezinta grade de toleranta la stres diferite si, prin urmare, pot fi clasificate in grupe functionale;
- prezinta importanta economica sau sunt o resursa de hrana importanta pentru specii cu valoare economica mare;
- joaca un rol important in transferul materiei organice si altor substante intre sediment si coloana de apa.
Pearson & Rosenberg (1978) descriu un model calitativ general al efectelor eutrofizarii si poluarii organice asupra organismelor bentice. Ei arata ca prima modificare detectabila consta in cresterea numarului de specii, urmata de cresterea biomasei si apoi de cea a abundentei numerice. Abundenta creste drastic la nivele ridicate ale poluarii organice si se datoreaza speciilor oportuniste. Pe masura cresterii incarcarii organice biomasa, numarul de specii si indivizi descreste rapid odata cu instalarea anaerobiozei.
Pe baza acestui model au fost propuse numeroase metode pentru descrierea si cuantificarea efectelor poluarii cu substante organice (Lambshead et al., 1983; Warwick, 1986, 1988, 1993; Heip et al., 1988; Gray et al., 1990, 2002; Clarke, 1993 etc.).
Pearson & Rosenberg (1978) propun 4 nivele de raspuns al bentosului la poluarea cu materii organice, care pot fi folosite in aprecierea gradului de eutrofizare a bentosului:
1) nivelul normal se caracterizeaza prin prezenta speciilor de talie mare, precum decapodele, si a speciilor care se pot ingropa adanc in sediment;
2) nivelul intermediar, caracterizat prin prezenta organismelor de talie mai mica, ce se hranesc la suprafata sedimentelor;
3) nivelul poluat, puternic dominat de polichete tubicole de talie mica si care indica o eutrofizare puternica;
4) nivelul puternic poluat, lipsit total de macrofauna din cauza prezentei la suprafata sedimentelor a unor portiuni contaminate cu hidrogen sulfurat.
Autorii realizeaza o ordonare a prezentei speciilor macrobentice de-a lungul unui gradient descrescator al poluarii organice. Astfel, din cele 99 de specii citate de ei primele 58 sunt polichete!
Prin urmare, predominarea anumitor specii de polichete constituie un indiciu bun al starii de sanatate a domeniului bental. La ora actuala exista numeroase studii in care stabilirea nivelului de eutrofizare si poluare organica s-a facut pe baza analizei comunitatilor de polichete (Reish, 1973; Grassle & Grassle, 1974; Losovskaya, 1978, 1983; Bellan, 1964a, 1980; Cognetti, 1969, 1972, 1973; Peres & Bellan, 1972).
Dupa Pocklington & Wells (1993) avantajele utilizarii polichetelor in biomonitoringul marin constau in:
- sunt abundente si relativ simplu de prelevat;
- apartin diferitor nivele trofice si prezinta o mare diversitate a strategiilor de reproducere, ceea ce le avantajeaza in popularea diferitor medii;
- raspund la diferite perturbari induse de diferite tipuri de poluare, prezentand modificari cuantificabile ale distributiei cantitative.
Multe specii de polichetele pot sa supravietuiasca un timp indelungat in conditiile diminuarii cantitatii de oxigen dizolvat in apa. Studiul realizat de Jakubova & Malm (1930) arata ca speciile Hediste diversicolor, Harmothoe imbricata si Eteone picta, care traiesc in malurile negre, pot supravietui timp de 7 zile in conditii de anaerobioza si timp de 3-6 zile in prezenta unei concentratii de hidrogen sulfurat de pana la 8,2 cm3/l. Dintre speciile ce populeaza biocenoza midiilor de piatra foarte sensibile la lipsa oxigenului s-au dovedit a fi Nereis zonata, Harmothoe impar, Platynereis dumerilii, Nereiphylla rubiginosa etc., in timp ce specia Lysidice ninetta a supravietuit 5 zile in lipsa oxigenului si 8 zile in prezenta unei concentratii de 12,6 cm3 H2S/l. Reprezentantii familiei Capitellidae supravietuiesc timp indelungat la concentratii de H2S de 20,4 cm3/l. Losovskaya (1962) constata in conditii de lipsa totala de oxigen din apa o mortalitate de 100% dupa 8 ore la Platynereis dumerilii si dupa 48 ore la Nereis zonata, in timp ce la Melinna palmata, in aceleasi conditii, dupa 48 ore mortalitatea era de doar 25%.
Un numar mare de polichete contribuie in mod activ la procesul de autoepurare naturala a apei marine. Multe specii de polichete sedentare din familiile Sabellidae, Serpulidae si Spirorbidae filtreaza mari cantitati de apa, eliberand-o de particulele in suspensie. Speciile de polichete detritivore pelofage (familiile Spionidae, Ampharetidae, Terebellidae si unele Nereididae) reduc cantitatea de substanta organica acumulata in sediment, care este o sursa secundara de poluare, detritusul organic fiind puternic transformat si incorporat ca biomasa proprie. In acest fel polichetele grabesc reintroducerea substantelor organice din apa si sediment in circuitul trofic, fara a mai fi trecute prin faza minerala. Polichetele sedimentivore (familiile Pectinariidae, Arenicolidae si Capitellidae) inghit cantitati mari de mal sau de nisip pe care il trec prin tubul lor digestiv, jucand un rol asemanator cu cel al ramelor in afanarea sedimentelor (Tenore, 1975, 1977). Acest lucru face posibila activitatea bacteriilor aerobe si grabirea mineralizarii substantelor organice din sediment.
Manoleli (1975) a estimat ca polichetul relict Hypania invalida filtreaza in 24 ore o cantitate de sediment ce depaseste de 9-10 ori volumul animalului insusi. Gomoiu (1982) a dedus ca populatiile de Melinna palmata din limitele platformei continentale romanesti pot prelucra pana la 4,8-9,6 kg mal/m2/zi!
Cel mai cunoscut indicator biologic este considerata a fi specia Capitella capitata, care caracterizeaza pozitiv gradul de poluare a apei si sedimentelor cu substante organice (Reish, 1957, 1970; Milovidova, 1975; Eagle & Rees, 1973; Grassle & Grassle, 1974). Dupa Losovskaya (1977, 1978) eutrofizarea si poluarea organica a acvatoriului Marii Negre a favorizat si dezvoltarea masiva a speciilor detritivore ca Polydora cornuta, Heteromastus filiformis, Pectinaria koreni si Melinna palmata.
O alta categorie de indicatori biologici o reprezinta speciile sensibile sau intolerante, care raspund in mod negativ la cresterea gradului de poluare. Dintre acestea Bellan (1980) mentioneaza pe reprezentantii genului Syllis sensu lato. Rygg (1985) face o clasificare a speciilor in conformitate cu distributia lor de-a lungul gradientului de diversitate. Din aproximativ 45 de specii absente din zonele cu diversitate mica, 25 sunt polichete!
Deoarece pentru o apreciere cat mai corecta a calitatii mediului acvatic este nevoie sa se tina cont de starea intregii biocenoze, in ultima vreme accentul se pune nu pe speciile indicatoare ci pe conceptul de comunitati indicatoare (Losovskaya, 1983).
Losovskaya (1977b, 1978) arata ca un indicator al degradarii biocenozei malurilor cu Mytilus galloprovincialis in partea de nord-vest a Marii Negre, ca urmare a instalarii in ultimul timp a mortalitatii in masa a organismelor bentonice cauzata de fenomenele de hipoxie si anoxie, poate servi si saracirea puternica a compozitiei specifice a polichetelor, caracterizata prin absenta cvasi-totala a speciilor epibentice (Nereiphylla rubiginosa, Harmothoe impar, Pomatoceros triqueter). Un indiciu privind refacerea acestei biocenoze il constituie cresterea proportiei speciilor detritofage mici (Spio decoratus, Polydora cornuta, Prionospio cirrifera, Capitella capitata si Capitella minima). Pe masura imbunatatirii starii biocenozei midiei creste numarul relativ al speciilor epibentice, in detrimentul polichetelor detritivore mici care nu intra in categoria speciilor caracteristice pentru biocenoza data.
Mai tarziu, Losovskaya (1983) propune pentru stabilirea calitatii mediului marin utilizarea raportului dintre numarul total al reprezentantilor familiei Capitellidae si cel al Spionidelor, ultimele fiind amensalii primilor. Predominarea numerica a Capitellidelor asupra Spionidelor da posibilitatea aprecierii indirecte a inrautatirii situatiei ecologice pentru sectorul marin dat (acumularea de substanta organica, inrautatirea regimului gazos cu toate consecintele sale etc.).
Bellan (1980) propune pentru exprimarea calitatii mediului marin un indice anelidian de poluare care reprezinta raportul dintre suma dominantelor speciilor santinele de apa poluata (de ex. Platynereis dumerilii) si a speciilor santinele de apa curata (de ex. speciile genului Syllis). Acest indice prezinta valori supraunitare in cazul mediilor poluate si subunitare in cazul mediilor nepoluate sau slab poluate.
Presiunea crescanda a factorului antropic asupra faunei bentonice din ultimii ani (regularizarea debitelor fluviale, urbanizarea si industrializarea litoralului, lucrarile de amenajare costiera, extinderea retelei de porturi, intensificarea traficului maritim etc.), precum si fenomenul global de eutrofizare si poluare, au determinat schimbari profunde in ceea ce priveste compozitia calitativa si distributia cantitativa a populatiilor de polichete de la litoralul romanesc al Marii Negre.
Cele mai afectate au fost populatiile de polichete din imediata apropiere a coastei, acestea fiind supuse in cea mai mare masura efectelor eutrofizarii antropice si poluarii organice. Efectele poluarii mediilor marine se traduc printr-o diminuare a numarului de specii, prin inlocuirea unor ansambluri de specii cu altele si prin modificarea raporturilor cantitative dintre specii.
Ca regula generala, in mediul marin se observa o oarecare reducere a numarului de specii odata cu cresterea severitatii degradarii conditiilor de mediu. Astfel, daca in trecut in nidsipurile fine de la nord de Constanta au fost identificate 27 de specii de polichete (Bacescu et al., 1967), in cercetarile noastre am putut identifica doar 16 specii. In mod asemanator, daca acum 30-40 de ani pentru substrat pietros au fost citate 35 de specii (Dumitrescu, 1973), noi nu am gasit decat 21 de specii. Totusi, comparand rezultatele cercetarilor noastre cu cele intreprinse in anii 1998-1999 (Surugiu, 2002, 2003), cand pe substrat nisipos au fost gasite 7 specii, iar pe cel pietros 20, se poate urmari o oarecare crestere a numarului de specii. In general, putem constata ca un numar de 25 de specii de polichete care traiau altadata la adancimi mai mici de 20 m nu au fost gasite in cercetarile noastre. Unele dintre acestea se pare ca au disparut definitiv, nu numai de la litoralul nostru ci si din intreg bazinul Marii Negre.
O consecinta a cresterii gradului de poluare este cresterea proportiei speciilor euriece in detrimentul celor stenece. Astfel, in conditiile cresterii cantitatii de suspensii aduse de fluviile tributare in partea de nord-vest a Marii Negre, in dreptul coastelor romanesti are loc o dezvoltare masiva a formelor de polichete detritofage endobentice, in special Spionidae si Capitellidae si o diminuare a efectivelor speciilor erante epibentice (Phyllodocidae, Syllidae). Dezvoltarea populatiilor acestor polichete mici duce la diminuarea biomaselor realizate de polichete si, prin urmare, la saracirea bazei trofice a pestilor bentofagi.
Eutrofizarea si poluarea organica a biocenozei nisipurilor fine cu Corbula mediterranea favorizeaza dezvoltarea masiva a speciilor detritivore ca Eteone picta, Spio decoratus, Polydora cornuta, Capitella capitata, Heteromastus filiformis, Pectinaria koreni si Melinna palmata. In schimb, specii precum Nephtys cirrosa, Micronephthys stammeri, Glycera alba, G. convoluta, Magelona rosea, M. papilicornis, Leiochone clypeata, Scolelepis cirratulus sau Pygospio elegans au devenit extrem de rare sau chiar au disparut.
5. CONCLUZII
1. Analiza structurii calitative si a distributiei cantitative a polichetelor din zonele luate in studiu a aratat ca, in linii mari, bogatia specifica si abundenta numerica este mai mica in punctele situate la nord de Constanta comparativ cu cele din sud. Indicele de diversitate specifica si echitabilitatea prezinta valori mici pentru Capul Midia, Mamaia si Mangalia, adica tocmai in punctele in care exista activitati portuare sau deversari de ape reziduale.
2. Gradul de eutrofizare si de poluare cu materii organice joaca, alaturi de tipul de sediment si de adancime, un rol foarte important in distributia populatiilor de polichete in zonele cercetate. In zonele afectate de poluare organica predomina speciile rezistente la hipoxie si chiar la anoxie temporara. Aceste specii, avand la dispozitie o resursa trofica abundenta sub forma materiei organice particulate (MOP) si in lipsa concurentei din partea altor specii, prezinta se dezvolta in masa, atingand biomase foarte ridicate.
3. Comparand datele existente in literatura cu cele ale cercetarilor noastre, am stabilit ca in decursul ultimilor 30 de ani, sub influenta intensificarii impactului antropic, fauna de polichete de la litoralul romanesc al Marii Negre a suferit schimbari calitative majore in sensul reducerii biodiversitatii. Astfel, am constatat ca un numar de 25 de specii de polichete care puteau fi intalnite altadata la adancimi mai mici de 20 m nu au mai fost semnalate.
4. In urma cercetarilor intreprinse in zona Agigei am reusit sa surprindem o serie de modificari ale structurii calitative si distributiei cantitative a faunei de polichete ca urmare a construirii Canalului Dunare - Marea Neagra si a extinderii portului Constanta. In primul rand, aceasta interventie majora s-a tradus prin disparitia completa a unor asociatii intregi impreuna cu speciile de polichete caracteristice lor. Chiar si in biocenozele neatinse de constructiile hidrotehnice am constatat o saracire faunistica substantiala, ceea ce indica si aici o stare generala de degradare a mediului marin.
5. Cercetarile noastre au probat ca la Agigea populatiile polichetului Nereis zonata, mai putin adaptat stresului poluarii si eutrofizarii, au fost inlocuite treptat de cele ale polichetului oportunist Neanthes succinea. In ultima vreme se constata o dezvoltare a populatiilor polichetului fitofil Platynereis dumerilii in detrimentul celor ale speciei Neanthes succinea, ceea ce ar indica o ameliorare a conditiilor de mediu de la litoralul romanesc.
6. In urma cercetarilor efectuate am constatat ca in categoria speciilor-indicatoare ale calitatii mediului marin trebuie sa fie incluse atat specii sensibile, stenobionte, precum Nereiphylla rubiginosa, Syllis gracilis, Typosyllis hyalina, Nereis zonata si Perinereis cultrifera, care raspund in mod negativ la degradarea conditiilor de mediu, cat si specii tolerante, euribionte, cu valente oportuniste, ca Neanthes succinea, Polydora cornuta, Capitella capitata, Prionospio cirrifera si Harmothoe imbricata, care raspund pozitiv la cresterea gradului de poluare si eutrofizare.
7. Pentru o apreciere cat mai corecta a calitatii mediului marin este nevoie, ca pe langa prezenta sau absenta speciilor indicatoare, sa se tina cont de structura cantitativa a comunitatilor de polichete, de abundentele si mai ales de biomasele realizate de speciile care intra in compozitia biocenozelor.
8. Studiul comunitatilor de polichete de la litoralul romanesc al Marii Negre ne permite sa apreciem ca in prezent apele costiere prezinta un regim mezotrof, in timp ce in zona de influenta a apelor Dunarene si in acvatoriile semiinchise ale porturilor Midia, Constanta Sud si Mangalia apele sunt eutrofe sau chiar hipertrofe.
bibliografie
Bacescu, M., 1954. Influenta iernii grele 1954 asupra vietii si pescuitului din Marea Neagra. Bul. Inst. Cerc. Pisc., 13(4): 5-12.
Bacescu, M., 1961. Cercetari fizico-chimice si biologice rominesti la Marea Neagra, efectuate in perioada 1954-1959. Hidrobiologia, 3: 17-46.
Bacescu, M., 1963. Contribution à la biocoenologie de la Mer Noire. L'étage périazoïque et la faciès paleo-dreissenifère et leurs caractéristiques. Rapp. et Pr. Verb. d. Réun. C.I.E.S.M.M., 17(2): 107-122.
Bacescu, M., 1965. Un aspect al influentei lucrarilor hidrotehnice asupra vietii marine litorale. Studii de hidraulica, 9: 137-149.
Bacescu, M., Dumitrescu, H., 1958. Les lagunes en formation aux embouchures du Danube et leur importance pour les poissons migrateurs. Verh. Internat. Ver. Limnologie, Stuttgart, 13: 699-709.
Bacescu, M., Dumitrescu, E., Manea, V., Por, F., Mayer, R., 1957. Les sables à Corbulomya (Aloidis) maeotica Mil., base trophique de premier ordre pour les poissons de la Mer Noire. Trav. Mus. Hist. Nat. Gr. Antipa, 1: 305-374.
Bacescu, M., Dumitrescu, E., Marcus, A., Palladian, G., Mayer, R., 1963. Données quantitatives sur la faune pétricole de la Mer Noire à Agigea (secteur roumain) dans la conditions spéciales de l'année 1961. Trav. Mus. Hist. nat. Gr. Antipa, 4: 131-155.
Bacescu, M., Gomoiu, M.-T., Bodeanu, N., Petran, A., 1965a. Studii asupra variatiei vietii marine in zona litorala nisipoasa de la Constanta. Ecologie marina, 1: 7-138.
Bacescu, M., Müller, G., Skolka, H., Petran, A., Elian, V., Gomoiu, M.-T., Bodeanu, N., Stanescu, S., 1965b. Cercetari de ecologie marina in sectorul predeltaic in conditiile anilor 1960-1961. Ecologie marina, 1: 185-344.
Bacescu, M., Gomoiu, M.-T., Bodeanu, N., Petran, A., Müller, G.I., Chirila, V., 1967. Dinamica populatiilor animale si vegetale din zona nisipurilor fine de la nord de Constanta in conditiile anilor 1962-1965. Ecologie marina, 2: 7-167.
Bacescu, M., Müller, G., Gomoiu, M.-T., 1971. Cercetari de ecologie bentala in Marea Neagra. Analiza cantitativa, calitativa si comparata a faunei bentale pontice. Ecologie marina, 4, 1-357.
Bellan, G., 1964a. Influence de la pollution sur la faune annélidienne des substrats meubles. Comm. int. Explor. sci. Mer Médit., Symp. Pollut. mar. Par Microorgan. Prod. pétrol., Monaco, 123-126.
Bellan, G., 1964b. Contributions à létude systématique, bionomique et écologique des Annélides Polychètes de la Méditerranée. Rec. Trav. Stat. Mar.Endoume, 49(33): 1-371.
Bellan, G., 1980. Annélides Polychètes des substrats solides de trois milieux polluées sur les côtes de Provence (France): Cortiou, Golfe de Fos, Vieux Port de Marseille. Tethys, 9(3): 267-278.
Blake, J.A., 1996. Family Spionidae Grube, 1850. Including a review of the genera and species from California and a revision of the genus Polydora Bosc, 1802. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel, Blake JA., B. Hilbig & P.H. Scott (eds.), Santa Barbara Museum of Natural History, Vol. 6: 81-223.
Borcea, I., 1931b. Action du froid et du gel sur la faune littorale de la Mer Noire. Ann. Sci. Univ. Jassy, 16: 751-759.
Clarke, K.R. & Warwick, R.M., 1994. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth Marine Laboratory, Natural Environment Research Council, 144 pp.
Cognetti, G., 1972. Distribution of polychaeta in polluted waters. Rev. Intern. Océanogr. Méd., 25: 23-34.
Cognetti, G., 1973. Sur la répartition de quelques polychètes en eau marine polluée. Archivio Oceanogr. Limnol., 18 (suppl.): 125-126.
Cognetti, G., Talierico, P., 1969. Policheti indicatori dellinquinamento delle acque. Pubbl. Staz. Zool. Napoli, 37(2 suppl.): 149-154.
Dumitrache Camelia, 1996-1997. Present state of the zoobenthos from the Romanian Black sea continental shelf. Cercetari marine, 29-30: 141-150.
Dumitrescu Elena, 1957. Contributii la studiul polichetelor din Marea Neagra, litoralul romanesc. Bul. stiint. Sect. biol. Seria zool., 9(2): 119-130.
Dumitrescu Elena, 1962. Nouvelle contribution à létude des Plychètes de la Mer Noire. Trav. Mus. Hist. Nat. Gr. Antipa, 3: 61-68.
Dumitrescu Elena, 1963. Polychètes marins de la zone littorale roumaine (1 à 20 m de profondeur). Trav. Mus. Hist. Nat. Gr. Antipa, 4: 181-192.
Dumitrescu Elena, 1973. Nouvelles données écologiques et quantitatives sur les Polychètes pétricoles de la Mer Noire (littoral roumain). Trav. Mus. Hist. Nat. Gr. Antipa, 13: 39-46.
Fauchald, K., 1977. The polychaete worms. Definitions and Keys to the Orders, Families and Genera. Natural Histiry Museum of Los Angeles County, Science Series, 28, 188 pp.
GESAMP, 1990. The State of the Marine Environment. United Nations Environment Programme. UNEP Regional Seas Rep. Stud., 115: 1-111.
GESAMP, 2001. A sea of troubles. GESAMP Rep. Stud., 70: 1-35.
Gomoiu, M.-T., 1963. LAnalyse granulométrique des sables de quelques plages de la Mer Noire (Côte Roumaine). Rapp. et Pr. Verb. d. Réun. C.I.E.S.M.M., 17(2): 123-131.
Gomoiu, M.-T., 1969. Studiul sedimentelor nisipoase de la litoralul romanesc al Marii Negre. Ecologie marina, 3: 227-325.
Gomoiu, M.-T., 1976a. Changes in the structure of benthic biocenoses from the romanian littoral of the Black Sea. Cercetari marine, 9 suppl.: 119-142.
Gomoiu, M.-T., 1982. On the populations of Melinna palmata Grube at the romanian littoral of the Black Sea. Cercetari marine, 15: 115-131.
Gomoiu, M.-T., 1985. Problemes concernant leutrophisation marine. Cercetari marine, I.R.C.M. Constanta, 18. 59-95.
Gomoiu, M.-T., 1997. Recifi artificiali la litoralul romanesc. An. Univ. Ovidius Constanta, Seria Biologie-Ecologie, 1(1): 159-174.
Gomoiu, M.-T., Müller, G.J., 1962. Studies concerning the benthic association dominated by Barnea candida in the Black Sea. Rev. Biol., 7(2): 255-271.
Gomoiu, M.-T., Tiganus, V., 1977. Données quantitatives concernant la structure des populations apartenant à la biocénose à Modiolus phaseolinus de la Mer Noire. Rapp. Comm. int. Mer Médit., 24(4): 123-124.
Grall, J., Chauvaud, L., 2002. Marine eutrophication and benthos: the need for new approaches and concepts. Global Change Biology, 8: 813-830.
Grassle, J.F., Grassle, J.P., 1974. Opportunistic life histories and genetic systems in marine benthic polychaetes. J. Mar. Res., 32(2): 253-284.
Grassle, J.P., Grassle, J.F., 1976. Sibling species in the marine pollution indicator Capitella (Polychaeta). Science, 192: 567-569.
Gray, J.S., Clarke, K.R., Warwick, R.M., Hobbs, G., 1990. Detection of initial effects of pollution on marine benthos: an example from the Ekofisk oilfields, North Sea. Mar. Ecol. Prog. Ser., 66: 285-299.
Gray, J.S., Shiu-sun Wu, R., Ying Or., Y., 2002. Effects of hypoxia and organic enrichment on the coastal marine environment. Mar. Ecol.Prog. Ser., 238: 249-279.
Jakubova, L.I., Malm, E.N., 1930. Iavlenie vremennogo anaerobioza u nekotorah predstavitelei bentosa Cernogo moria. Doklada Akad. Nauk SSSR, 14: 363-366.
Kiseleva, M.I., 1981. Bentos rahlah gruntov Cernogo moria. Izd. Naukova dumka, Kiev, 168 pp.
Losovskaya, G.V., 1962. Otnosenie nekotorah cernomorskih polihet k izmeneniu solenosti, gazovogo rejima i grunta. Voprosa ekologhii, 5: 115-117.
Losovskaya, G.V., 1976. O rassirenii areala Polydora limicola Annenkova novogo dlia Cernogo moria vida polihet. Ghidrobiol. Jurn., 12(1): 102-103.
Losovskaya, G.V., 1977a. Nekotorae osobennosti sovremennogo sostoiania zoobentosa severo-zapadnoi ciasti Cernogo moria. Biologhia moria, 43: 25-33.
Losovskaya, G.V., 1977b. The ecology of polychaetes of the Black Sea. Naukova Dumka, Kiev, 92 pp.
Losovskaya, G.V., 1978. Vlianie ghipo- i anoksii na vidovoi sostav i cislennost cernomorskih Polychaeta. Ghidrobiol. Jurn., 14(4): 29-32.
Losovskaya, G.V., 1983. O znacenii polihet kak vozmojnah indikatorov kacestva sreda Cernogo moria. Ekologhia moria, 12: 73-78.
Losovskaya, G.V., 1988. Long-term changes in the composition and distribution of polychaetes of the north-western part of the Black Sea. Gidrobiol. zhurn., 24(4): 21-25.
Losovskaya, G.V., 1991. Melkie detritoiadnae poliheta v donnah soobscestvah severo-zapadnoi ciasti Cernogo moria. Ghidrobiol. Jurn., 27(6): 24-29.
Losovskaya, G.V., Nesterova, D.A.,1964. O massovom razvitii novoi dlia Cernogo moria forma mnogoscetinkovogo kolciatogo cervia Polydora ciliata ssp. limicola Annenkova v Suhom limane (severo-zapadnaia ciast Cernogo moria): Zool. Jurn., 43(10): 1559-1560.
Marinov, T., 1977. Fauna na Balgaria. Mnogocetinesti cervei (Polychaeta). Izd. na Balgarskata Akademia na Naukite, Sofia, 6, 258 pp.
Markovski, N., 1954. Fauna bespozvonocinah nizoviev rek Ukraina, uslovia eio suscestvovania iputi ispolzovania. Dneprovsko-Bugskii liman, vol II, Izd. Akad. Nauk. USSR, Kiev, 196 pp.
Manoleli, D., 1975. On the biology, distribution and origin of Polychaeta from the Danube and Danube Delta. Trav. Mus. Hist. Nat. Gr. Antipa, 16: 27-33.
Manoleli, D., 1979. Des modifications survenues ces 30 dernieres annes dans la composition de la faune benthique du littoral roumain (Mer Noire). Atti Soc. Tosc. Sci. Nat., Mem., ser. B, 86, suppl.: 152-159.
Manoleli, D., 1980. Les populations de Polychètes des eaux saumatres oligohalines de la Roumanie. Quelques observations et problèmes. Trav. Mus. Hist. Nat. Gr. Antipa, 21: 111-129.
Milovidova, N.Yu., 1975. Izmenenie donnah biotenozov Sevastopolskoi buhta za period s 1913 po 1973 gg. Biologhia moria, 35: 117-124.
Müller, G.J., 1995. Marea Neagra, prezentare generala. in Diversitatea lumii vii. Determinatorul ilustrat al florei si faunei romanesti, Vol. I - Mediul marin, Ed. Bucura Mond, Bucuresti: 1-29.
Murina, V.V., Mikhailova, T.V., 1994. Raspredelenie Neanthes succinea (Polychaeta) v bentali I pelaghiali Phyllophornogo polia Cernogo moria. Ghidrobiol. Jurn., 30(1): 19-27.
Nicoara, M., Mustata, Gh., Trandafirescu, I., Surugiu, V., Palici, Cristina, 1995-1997. Factors causing spatial variation in zooplancton and benthic populations of the Black Sea coastal waters in the Portita-Periboina area. Analele Stiintifice ale Universitatii Al. I. Cuza Iasi, s. Biologie animala, Tom 41-43: 17-24.
Peranu A., 1997. Black Sea Biological Diversity. Romania. Black Sea Environmental Series, Vol. 4, UN Publications, 314 pp.
Pearson, T.H., Rosenberg,R., 1978. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev., 16: 229-311.
Pérès, J.-M., Bellan, G., 1972. Aperçu sur lInfluence des Pollutions sur les Peuplements Benthiques. 3-14.
Pocklington, P. & Wells, P.G., 1992. Polychaetes - Key taxa for marine environmental quality monitoring. Mar. Poll. Bull., 24: 593-598.
Rasmussen, E., 1973. Systematics and ecology of the Isefjord marine fauna (Denmark). Ophelia, 11(1-2): 1-507.
Reish, D.J., 1970. The effects of varying concentrations of nutrients, chlorinity, and dissolved oxygen on polychaetous annelids. Water Research, 4: 721-735.
Reish, D.J., 1973. The use of benthic animals in monitoring the marine environment. Journal of Environmental Planning and Pollution Control, 1(3): 32-38.
Reish, D.J., 1973. The use of marine invertebrates as indicators of water quality. Journal of Environmental Planning and Pollution Control, 1(3): 92-103.
Rygg, B., 1985. Distribution of Species along Pollution-induced Diversity Gradients in Benthic Communities in Norwegian Fjords. Marine Pollution Bulletin, 16(12): 469-474.
Sergeeva, N.G., 2000. K voprosu o biologhiceskom raznoobrazii glubokovodnogo bentosa Cernogo moria. Ekologhia moria, 50(7): 57-62.
Sergeeva, N.G., Zaika, V.E., 2000. Ekologhia polihet iz pogranicinah soobscestv pelaghiali i bentali Cernogo moria. Dopovidi Nationalnoi Akad. Nauk Ukraini, 1: 197-201.
Surugiu, V., 2000. Des modifications survenues dans la structure des populations des Annélides Polychètes dAgigea dans les 30 dernières années. Analele Stiintifice ale Universitatii Al. I. Cuza Iasi, s. Biologie animala
Surugiu, 2002. Populatiile de polichete (Annelida) de la litoralul romanesc al Marii Negre si din sistemul lagunar si rolul lor ecologic. Teza de doctorat, Universitatea Alexandru Ioan Cuza din Iasi, 1-277.
Surugiu, V., 2003. Near-shore polychaete assemblages of the southern sector of the Romanian Black Sea littoral. Revue Roumaine de Biologie, Série de Biologie animale
Surugiu, V., 2004. Utilizarea polichetelor ca indicatori ai eutrofizarii apelor costiere. Studiu de caz: litoralul romanesc al Marii Negre. Sesiunea Stiintifica Nationala Biologia la inceputul secolului XXI, Iasi, 13-14 noiembrie, Program si rezumate: 9-10.
Surugiu, V., Palici Cristina, Manoleli, D., 2001. Date preliminare asupra biodiversitatii bentofaunei lacului Mangalia. in 'Diversitatea, valorificarea rationala si protectia lumii animale', CE USM, Chisinau, pp. 236-238.
Surugiu, V., Zamfirescu, St., 2004. Quantitative Analysis of Macrozoobenthic Communities of Mangalia Bay. Studii si Cercetari Stiintifice Seria Biologie, 9: 48-53.
Surugiu, V., Mustata, Gh., Hartascu, M., 2004. Contributions to the qualitative and quantitative study of the macrozoobenthos from the Danube Black Sea Canal. Studii si Cercetari Stiintifice Seria Biologie
Tiganus, V., 1978. Observations sur la structure qualitative et quantitative de la biocénose des moules de rocher du littoral roumain de la Mer Noire. Rapp. Comm. int. Mer Médit., 25: 159-160.
Tiganus, V., 1981. Données quantitatives sur la faune pétricole de petite profondeur du littoral roumain de la Mer Noire. Rapp. Comm. int. Mer Médit., 27(2): 157-158.
Tiganus, V., 1982a. Evolution des principales communautés benthiques du secteur marin situé devant les embouchures du Danube. Cercetari marine, 15: 89-106.
Tiganus, V., 1982b. Données préliminaires sur le zoobenthos du substrat meuble de la zone portuaire Constanta. Cercetari marine, 15: 107-114.
Tiganus V., 1986. Structure des peuplements de Polychètes de substrat sableux sous condition de forte eutrophisation en Mer Noire. Rapp. Comm. int. Mer Médit., 30(2): 20.
Tiganus V., 1990. Evolution des peuplements macrobenthiques du substrat sableux sur le littoral roumain. Rapp. Comm. int. Mer Médit., 32(1): 22.
Tiganus, V., 1992. Evolution des populations de certaines especes de masse de Polychètes de la zone marine roumaine. Rapp. Comm. Int. mer Médit., 33: 54.
Tiganus, V., 1997. Importanta cunoasterii starii comunitatilor bentale in aprecierea gradului de poluare a unei zone marine. An. Univ. Ovidius Constanta, Seria Biologie-Ecologie, 1(1): 99-104.
Vinogradov, K.A., Losovskaya, G.V., 1968. Tip kolciatae cervi - Annelida: Klass mnogoscetinkovaie cervi - Polychaeta, in Opredelitel fauna Cernogo i Azovskogo morei, vol. 1, Izd. Naukova Dumka, Kiev: 251-359.
Warwick, R.M., 1986. A new method for detecting pollution effects on marine macrobenthic communitise. Marine Biology, 92: 557-562.
Warwick, R.M., 1988. The Level of Taxonomic Discrimination required to Detect Pollution Effects on Marine Benthic Communitise. Marine Pollution Bulletin, 19(6): 259-268.
Zaitsev, Zu.P., 1991. Cultural eutrophication of the Black Sea and other South European Seas. Bull. Soc. fr.-jap. Océanogr., 29(1): 1-7.
Zaitsev, Yu., Mamaev, V., 1997. Biological Diversity in the Black Sea. A Study of Change and Decline. Black Sea Environmental Series, Vol. 3, UN Publications, 208 pp.
Zernov, S.A., 1913. K voprosu ob izucenii jizni Cernogo moria. Zap. Imp. Akad. Nauk S.Pb., ser. 8, Fiz.-mat. otd-nie, 32(1): 1-299.
Copyright © 2025 - Toate drepturile rezervate