|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Holoproteidele sau proteinele, sunt substante complexe, macromoleculare, cu un grad inalt de organizare structurala, alcatuite numai din aminoacizi. Masele moleculare a acestora variaza intre 1.103 si 1.105 .
Molecula proteinelor este formata din resturi de aminoacizi uniti intre ei prin legaturi peptidice, dand nastere unui polimer liniar. In principiu, formula structurala a unui fragment din catena polipeptidica a unei proteine, poate fi reprezentat astfel:
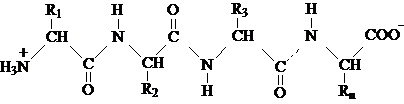
In aceasta catena, radicalii R1, R2 . . Rn caracteristici aminoacizilor constituenti sunt:
resturi hidrocarburice nepolare, hidrofobe ( - CH3 , - CH(CH2)3 provenite de la alanina, valina, etc).
resturi ce contin grupari polare (- OH , - SH provenite de la serina, cisteina).
resturi ionogene, hidrofile ( - NH2 , - COOH provenite de la aminoacizii cu caracter bazic sau acid).
Fiecare proteina specifica se caracterizeaza prin numarul aminoacizilor constituenti, felul acestora, precum si ordinea (secventa) lor in lantul polipeptidic.
Pentru stabilirea secventei aminoacizilor in lantul polipeptidic se utilizeaza hidroliza enzimatica, la care participa urmatoarele enzime: tripsina, chimotripsina, pepsina si papaina. Acestea scindeaza legaturile peptidice ale proteinei, rezultand aminoacizii componenti.
Structura complexa a proteinelor se datoreste faptului ca ele sunt substante macromoleculare si prezinta o configuratie spatiala, tridimensionala.
Configuratia macromoleculelor proteice. Proteinele se caracterizeaza prin organizare spatiala specifica, respectiv prin configuratie. Aceasta se refera la modul in care atomii sau gruparile de atomi, prezente in molecula sunt orientate in spatiu, precum si la legaturile ce se stabilesc intre lanturile polipeptidice.
Factorii amintiti contribuie direct la formarea edificiului spatial al macromoleculei proteice. Aceasta determina proprietatile biologice ale proteinelor.
Structura de ansamblu a unei macromolecule proteice este rezultatul interactiunii a diferite nivele de organizare structurala si anume:
structura primara;
structura secundara;
structura tertiara;
structura cuaternara.
Structura primara se refera la natura aminoacizilor, numarul acestora, precum si la ordinea in care ei se asaza in lantul polipeptidic. Resturile de α-aminoacizi se leaga intre ele prin legaturi peptidice - CO - NH - , care se stabilesc intre gruparile - COOH, respectiv - NH2 ale diferitilor aminoacizi. Catena polipeptidica proiectata in plan are forma unui zig-zag, iar radiacalii R ale aminoacizilor constituenti sunt dispusi alternativ deasupra si dedesubtul planurilor formate de aceasta:
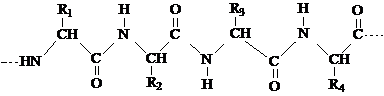
Pentru stabilirea structurii primare a proteinelor se utilizeaza:
metode fizico-chimice;
metode chimice;
metode biochimice. Intre metodele utilizate cu mult succes in stabilirea
structurii primare a proteinelor s-au remarcat: cromatografia, electroforeza, hidroliza enzimatica, hidroliza chimica, etc.).
Metodele analitice utilizate pentru elucidarea structurii primare sunt deosebit de complexe si laborioase. Din acest motiv, abia in 1955 Sanger reuseste sa stabileasca structura insulinei (hormon polipeptidic format din 51 aminoacizi si M = 6000), iar cativa ani mai tarziu Moore si Stein stabilesc structura primara a ribonucleazei (enzima formata din 124 aminoacizi si M = 13000).
Studiul cu radiatii X a proteinelor cristalizate a aratat ca macromoleculele proteice au conformatii tridimensionale. Lanturile polipeptidice ale proteinelor sunt legate intre ele prin diferite tipuri de legaturi, care pot fi mai puternice sau mai slabe. Acestea sunt:
legaturi de hidrogen;
legaturi covalente;
legaturi electrovalente;
forte van der Waals.
Structura secundara se prezinta in trei modele:
modelul α helix;
modelul in foaie pliata;
modelul de tip colagen.
a) Modelul α-helix reprezinta cel mai simplu aranjament spatial rezultat prin spiralarea spre dreapta a catenei polipeptidice in jurul unui cilindru imaginar. Se presupune ca:
aminoacizii constituenti prezinta configuratie L;
distantele interatomice si unghiurile de valenta prezinta aceleasi valori, indiferent de marimea catenei polipeptidice;
legatura peptidica este coplanara, aceasta conformatie fiind cea mai favorabila din punct de vedere energetic;
in macromolecula proteica se formeaza un numar maxim de legaturi de hidrogen.
![]() Modelul α helix este caracterizat prin pasul elicei (distanta
masurata intre doua pozitii identice succesive pe helix),
care are valoarea 5,4 Å. Pe fiecare spira a helixului revin cate 3,6
resturi de aminoacizi. Astfel in spirele invecinate se gasesc
fata in fata gruparile CO si - NH ale legaturii
peptidice, intre care se stabilesc punti de hidrogen.
Modelul α helix este caracterizat prin pasul elicei (distanta
masurata intre doua pozitii identice succesive pe helix),
care are valoarea 5,4 Å. Pe fiecare spira a helixului revin cate 3,6
resturi de aminoacizi. Astfel in spirele invecinate se gasesc
fata in fata gruparile CO si - NH ale legaturii
peptidice, intre care se stabilesc punti de hidrogen.
Structura α helix apare atat in proteinele fibrilare (α keratina din par si lana), cat si in cele globulare, in care predomina aminoacizii cu catene laterale R, voluminoase. Acestia sunt orientati spre exteriorul spiralei.
Proteinele native nu sunt insa alcatuite din conformatii α helix regulate, in structura lor apar si regiuni neelicoidale. Existenta lor este determinata de natura radicalilor R a unor aminoacizi, intre care se exercita atractii sau respingeri. Procentul de structura α helix a proteinelor globulare variaza de la o proteina la alta: 0-10 % in cazeina, actina, γ-globulina, intre 10-20 % in ribonucleaza, intre 20-30 % in pepsina si histone, 30-45 % in ovalbumina si fibrinogen, 60-80 % in mioglobina si hemoglobina.
b) Modelul de foaie pliata reprezinta aranjamentul spatial al macromoleculelor proteice, rezultat in urma interactiunii prin legaturi de hidrogen dintre diferitele catene polipeptidice. Acest tip de structura apare la proteinele fibrilare din firul de par (β-keratina) si din firul produs de viermele de matase (fibroina). Lanturile polipeptidice ale proteinelor pot fi dispuse paralel sau antiparalel, adica cu gruparile R din diferitele catene polipeptidice orientate in acelasi sens (β-keratina) sau in sensuri diferite. In acest fel, alternanta gruparilor NH si CO apare inversa in cele doua lanturi.
Aranjamentul spatial sub forma unei foi pliate confera moleculei o mare stabilitate.
Proteinele cu structura α-helix pot trece in anumite conditii in proteine cu structuri de foaie pliata. Astfel, α-keratina, proteina dinpar supusa actiunii apei calde sau alcaliilor diluate si a tractiunii, se poate transforma in β-keratina.
![]() α-keratina β-keratina
α-keratina β-keratina
(α-helix) (foaie pliata)
La trecerea din structura α-helix in structura de foaie pliata legaturile de hidrogen intracatenare din α-helix se desfac, iar catenele polipeptidice se grupeaza paralel, cu formare de legaturi de hidrogen intercatenare, caracteristice structurii in foaie pliata. Reactia este reversibila, iar incetarea tractiunii firului duce la refacerea formei initiale (α-keratina).
c) Modelul tip colagen. Proteinele fibrilare din colagen reprezinta circa 30 % din totalul proteinelor organismului animal. Ele sunt prezente in tesuturile conjunctive. Colagenul se prezinta sub forma unor fibre liniare foarte extensibile. Colagenul contine in proportie ridicata glicocol, prolina si hidroxiprolina. Acesti aminoacizi fac imposibila structura α-helix sau de foaie pliata, intrucat acesti aminoacizi nu pot participa la formarea legaturilor de hidrogen.
Colagenul este alcatuit dintr-un suprahelix format din trei catene polipeptidice spiralate spre stanga si infasurate spre dreapta in jurul axei comune. In figura urmatoare este prezentat un fragment din catena polipeptidica a colagenului:
Structura tertiara a proteinelor reprezinta un grad avansat de organizare spatiala a acestora. Ea reprezinta rezultatul interactiunilor dintre resturile R ale aminoacizilor din catenele polipeptidice, care pot stabili diferite tipuri de legaturi intramoleculare. Acestea sunt:
legaturi de hidrogen nepeptidice ce se stabilesc intre gruparea - OH a tirozinei si un rest carboxilic apartinand aminoacizilor cu caracter acid (aspartic si glutamic) sau intre restul imidazolic al histidinei si gruparea - OH a serinei;
legaturi ionice intre resturile - COOH ale aminoacizilor dicarboxilici si resturile - NH2 ale aminoacizilor bazici, ele fiind situate spre exteriorul moleculei si intra in interactiune cu solventul;
legaturi covalente formate prin punti disulfidice ( - S - S - ) intre resturile de cisteina;
forte van der Waals intre radicalii hidrofobi ai alaninei, valinei, leucinei. Acest tip de atractie are ca rezultat excluderea moleculelor de solvent polar in regiunea respectiva si sunt situate spre interiorul moleculei;
interactiuni dipol-dipol, ce se stabilesc intre gruparile - OH din serina.
Acest tip de structura este caracteristic proteinelor globulare: hemoglobina, mioglobina, ribonucleaza, tripsinogen, etc.
Legaturile si interactiunile caracteristice structurii tertiare sunt destul de labile si pot fi usor desfacute prin actiunea diferitilor factori fizici si chimici (pH, temperatura, presiune, reactivi chimici). Astfel denaturate, proteinele vor pierde proprietatile lor biologice.
Acest tip de structura reprezinta nivelul cel mai inalt de organizare a proteinelor si se datoreste interactiunii dintre catenele polipeptidice independente, care sunt caracterizate prin structuri primare, secundare si tertiare bine definite. Ea apare in cazul proteinelor globulare cu mase moleculare mari. Acestea exista sub forma unor agregate moleculare si se numesc proteine oligomere. Ele sunt alcatuite din doua sau mai multe catene polipeptidice (protomeri). In functie de numarul acestora putem vorbi de dimeri, trimeri, tetrameri, etc. Agregarea protomerilor se datoreste unor interactiuni de natura electrostatica (legaturi ionice si legaturi de hidrogen) care sunt localizate la suprafata fiecarei catene.
Hemoglobina din sange prezinta o structura care se gaseste sub forma unui tetramer alcatuit din patru catene polipeptidice, identice doua cate doua. Aceste catene alcatuiesc un agregat molecular in care sunt prezente si patru grupari neproteice, numite hem si care sunt implicate in transportul oxigenului.
Structura cuaternara poate fi dezorganizata cu usurinta prin diluarea solutiilor, modificarea pH-ului sau in prezenta unor saruri, obtinandu-se protomerii constituenti. Prin indepartarea agentului ce a produs disocierea, se constata reasamblarea rapida a catenelor cu formarea suprastructurilor moleculare.
Masa moleculara a proteinelor variaza in limite foarte largi si anume de la zeci de mii pana la zeci de milioane. Metodele utilizate pentru determinarea masei moleculare, se bazeaza pe:
masurarea presiunii osmotice (Sörensen);
masurarea vitezei de sedimentare (Svedberg);
masurarea vitezei de difuziune;
masurarea difractiei luminii;
analiza röntgenografica;
gel - filtrare.
Greutatile moleculare a catorva proteine sunt cuprinse in tabelul de mai jos. Se constata ca proteinele cu masa moleculara mare si foarte mare, sunt formate din mai multe subunitati, iar catenele polipeptidice contin 300 - 600 resturi de aminoacizi, pe cand cele cu masa moleculara mica sunt formate dintr-un singur lant polipeptidic, ce contine 110 - 120 aminoacizi. S-a constatat ca mase moleculare foarte mari se intalnesc adesea la enzime, datorita agregarii lanturilor polipeptidice.
Proteinele sunt substante care prezinta o serie de proprietati fizico-chimice, chimice si biologice. Aceste proprietati se datoresc structurii macromoleculare a proteinelor, precum si gruparilor polare si nepolare care se gasesc la suprafata acestor molecule.
1. Solubilitatea proteinelor. Aceasta depinde de existenta gruparilor polare hidrofile de la suprafata macromoleculelor ( - COOH, - NH2, - OH, - SH ) care stabilesc legaturi cu moleculele solventilor polari (apa, glicerolul). Majoritatea proteinelor sunt solubile in apa, cu care formeaza solutii coloidale, precum si in solutii saline. Proteinele greu solubile sufera in contact cu apa un fenomen de gonflare, care se datoreste hidratarii gruparilor polare prezente.
Macromoleculele proteice se comporta in solutie apoasa ca macroamfioni datorita prezentei gruparilor ionizabile - COOH si - NH2 la suprafata acestora. In functie de pH-ul solutiei, proteinele se comporta fie ca anioni (in mediu bazic) sau cationi (in mediu acid), la fel ca aminoacizii. Exista o valoare a pH-ului la care numarul sarcinilor pozitive a moleculei este egal cu numarul sarcinilor negative, deci sarcina electrica devine nula. Acest pH poarta denumirea de punct izoelectric al proteinei si se noteaza cu pHi. Punctul izoelectric al proteinelor variaza in limite foarte largi (1 - 12,5) in functie de numarul gruparilor acide sau bazice care predomina pe suprafata macromoleculei. La pHi solubilitatea proteinelor este minima, datorita atractiei sarcinilor electrice de semn contrar, fapt ce favorizeaza agregarea moleculelor. In tabelul de mai jos sunt prezentate valorile pHi a unor proteine. Datorita caracterului amfoter proteinele se comporta ca sisteme tampon, care mentin la valori constante pH-ul mediului biologic in care se gasesc.
Valorile pHi a unor proteine
= = = = = = = = = = = = = = = = = =
Proteina pHi Proteina pHi
__________ ______ ____ __________ ______ ____
Pepsina 1,0 Miozina 5,4
Cazeina 4,6 Fibrinogen 5,5
Ovalbumina 4,6 Gliadina 6,5
Serumalbumina 4,7 Hemoglobina 6,9
Ureaza 5,1 Globina 8,1
Insulina 5,4 Citocrom C 10,5
= = = = = = = = = = = = = = = = = =
Datorita gruparilor ionizabile si a aparitiei in solutie proteica a macroanionilor sau macrocationilor, proteinele au proprietatea de a migra in camp electric. Fenomenul se numeste electroforeza. Daca macromolecula va fi incarcata negativ ea va migra spre anod (anaforeza), daca insa vor predomina sarcinile pozitive, ea se va deplasa spre catod (cataforeza). Viteza de migrare in camp electric a diferitelor proteine este diferita, ceea ce permite separarea prin electroforeza a amestecurilor de proteine.
2. Activitate optica. Proteinele prezinta activitate optica levogira, datorita prezentei in molecula a aminoacizilor din seria L. In lumina ultravioleta (λ = 280 nm), proteinele prezinta un maxim de absorbtie, datorat aminoacizilor aromatici. Pe aceasta proprietate se bazeaza dozarea proteinelor prin metoda spectrofotometrica.
3. Denaturarea. Proteinele native pot sa sufere transformari sub actiunea unor agenti fizico-chimici care duc la dezorganizarea structurii spatiale, fara a fi afectata structura primara si masa moleculara a acestora. Fenomenul se numeste denaturare si este rezultatul desfacerii legaturilor ce caracterizeaza structura secundara si tertiara.
Aceste transformari sunt, de obicei, ireversibile si au drept consecinta modificarea proprietatilor fizice ale proteinelor (scaderea solubilitatii, cresterea activitatii optice, modificarea vascozitatii si a presiunii osmotice, cresterea valorii pHi, pierderea tendintei de cristalizare) precum si pierderea activitatii biologice.
Agentii care provoaca denaturarea sunt de natura fizica sau chimica (acizi, baze, sarurile metalelor grele, solventi organici, detergenti). Daca actiunea agentului denaturant este limitata si moleculele proteice sunt numai partial denaturate, procesul poate fi reversibil, iar prin indepartarea cauzei denaturarii, moleculele proteice se pot renatura dobandind structura si proprietatile biologice initiale.
Proteinele au, in general, o reactivitate chimica ridicata, datorita gruparilor functionale libere ale aminoacizilor constituenti, precum si a nucleelor aromatice sau a resturilor hidrocarburice. Legaturile peptidice imprima si ele proprietati chimice specifice.
a) Hidroliza. Proteinele, in functie de natura lor, hidrolizeaza mai usor sau mai greu in prezenta acizilor, bazelor sau a enzimelor specifice. Hidroliza consta in desfacerea legaturilor peptidice in prezenta apei, rezultand aminoacizi.
Hidroliza partiala se desfasoara in conditii blande, cu acizi diluati sau cu enzime proteolitice si se pot obtine polipeptide si peptide cu masa moleculara tot mai mica. Acest tip de hidroliza este utilizat pentru stabilirea structurii primare a proteinelor.
b) Reactii de culoare. Aceste reactii se datoresc aminoacizilor constituenti ai proteinelor care reactioneaza specific cu anumiti reactivi, dand reactii de culoare. Principalele reactii de culoare sunt reactia ninhidrinei (caracteristica gruparii - NH2 din aminoacizi), reactia xantoproteica si Millon (datorita nucleelor aromatice) si reactia biuretului (datorita legaturilor peptidice - CO - NH - ). Reactia biuretului este utilizata curent pentru dozarea colorimetrica a proteinelor, deoarece intensitatea culorii este direct proportionala cu concentratia acestora in solutie.
c) Reactii de precipitare. Proteinele existente in solutii pot fi precipitate sub actiunea unor reactivi. In functie de natura reactivilor, precipitarile pot fi reversibile sau ireversibile.
Precipitarea reversibila se realizeaza sub actiunea sarurilor [Na2SO4, (NH4)2SO4] sau a unor alcooli. Acestia nu produc modificarea structurii, ci doar deshidratarea partiala a macromoleculelor care se aglomereaza si precipita. Proteinele precipitate prin salifiere se pot redizolva dupa indepartarea agentilor chimici ce au produs precipitarea.
Precipitarea ireversibila are loc sub actiunea unor reactivi chimici, care produc modificari profunde ale structurii proteinelor, respectiv denaturarea lor. Dintre reactivii ce produc precipitari ireversibile, amintim acizii anorganici (HCl, HNO3), acizii organici (acidul tricloracetic, acidul picric si acidul sulfosalicilic) si sarurile metalelor grele (Cu2+, Pb2+, Hg2+, Ag+).
Coagularea este precipitarea ireversibila a proteinelor sub actiunea caldurii. Temperatura de coagulare depinde de natura si concentratia sarurilor prezente in solutie. In absenta sarurilor unele proteine nu precipita nici la fierbere.
Proteinele prezinta proprietati biochimice specifice:
hormoni;
enzime;
contractie;
transport;
imunologice.
Holoproteidele (proteinele) sunt formate, dupa cum s-a mai aratat, numai din aminoacizi. Dupa rolul lor fiziologic proteinele se grupeaza in doua clase, si anume;
proteine vii (netransformate) solubile si active din punct de vedere biologic, care prezinta toate proprietatile fizico-chimice si chimice descrise anterior;
proteine transformate (scleroproteine) care au suferit un proces de transformare in organismele vii, in urma caruia si-au pierdut activitatea biologica si nu mai prezinta proprietatile caracteristice proteinelor vii, ele fiid insolubile.
Acestea au structura globulara si sunt prezente in toate organismele vii, vegetale si animale. Ele pot sa aiba caracter neutru, acid sau bazic, in functie de raportul diferitelor tipuri de aminoacizi din molecula. Proteinele globulare se impart in sase grupe, pe baza provenientei si a proprietatilor fizice si chimice:
![]()
- albumine
universal raspandite
- globuline
- histone
regnul animal
- protamine
- prolamine
regnul vegetal
- gluteline
In majoritatea cazurilor proteinele, apartinand diferitelor grupe, apar impreuna, iar separarea si caracterizarea lor este deosebit de dificila.
Albumine. Sunt proteine globulare cu caracter neutru sau slab acid. In constitutia lor intra aminoacizii cu caracter neutru. Nu contin glicocol. Ele sunt solubile in apa si solutii diluate de electroliti. Precipitarea lor necesita solutii concentrate de saruri [(NH4)2SO4 70 - 100%)]. Albuminele coaguleaza prin incalzire la 60 - 70s C. Datorita faptului ca au masa moleculara relativ mica, ele manifesta in campul electric o mobilitate deosebita si pot patrunde prin membrane subtiri.
Albuminele sunt raspandite in citoplasma si in lichidele biologice ale organismelor animale. Cele mai importante sunt: seralbuminele, care reprezinta 52-62 % din totalul proteinelor din plasma sanguina, lactoalbumina din lapte, ovalbumina din albusul de ou si mioalbumina din tesutul muscular. Albuminele animale indeplinesc roluri multiple:
-regleaza presiunea osmotica a sangelui;
-constituie o rezerva de proteine pentru organism;
-contribuie la tamponarea pH-ului sanguin;
-indeplinesc functia de transport a anionilor acizilor grasi.
In regnul vegetal albuminele apar indeosebi in seminte si poarta denumiri specifice, dupa originea lor: legumelina (in semintele de leguminoase), faseolina (in fasole), leucozina (in grau), ricina (in ricin), etc.
Unele plante contin albumine toxice: crotina din Croton tiglium, falina din Amanita phaloides, etc.
Globuline. Sunt holoproteide cu caracter slab acid, datorita continutului ridicat in acid aspartic si glutamic. Ele sunt greu solubile in apa, insa sunt solubile in solutii diluate de saruri neutre (NaCl, Na2SO4) si de baze. Ele au tendinta de precipitare atat la diluarea solutiei cu apa cat si la salifiere (NH4)2SO4 (50 %). Globulinele au mase moleculare mai mari decat albuminele (de ordinul 105 - 106) dar cu toate acestea, solutiile lor coaguleaza mai greu sau deloc la fierbere.
Globulinele sunt cele mai raspandite proteine globulare din organismul animal. In plasma sanguina se gaseste serumglobulina si fibrinogenul, in lapte lactoglobulina, in ou ovoglobulina, in muschi miozina.
In regnul vegetal globulinele apar mai ales in seminte, alaturi de albumine. Dupa originea lor, globulinele vegetale poarta denumiri specifice: glicina din soia, edestina din canepa sau legumina din mazare.
Histone. Sunt holoproteide cu caracter slab bazic, datorita prezentei in molecula a aminoacizilor cu caracter bazic (arginina si lizina). Ele sunt solubile in solutii acide. Coaguleaza prin incalzire si pot fi hidrolizate in prezenta pepsinei.
Histonele apar sub forma libera in leucocite si sperma pestilor, dar indeosebi asociate cu acizi nucleici sub forma de nucleoproteide (nucleohistone) in nucleul celulelor glandulare (pancreas, timus), celule spermatice, eritrocite nucleate, etc.
Protamine. Sunt holoproteide cu caracter bazic accentuat datorita unui continut de aminoacizi cu caracter bazic (arginina, lizina, histidina). In molecula lor lipsesc aminoacizii cu sulf, tirozina si triptofanul. Protaminele au masa moleculara mica (2.103 - 3.103), usor solubile in apa, nu coaguleaza la incalzire si sunt greu hidrolizabile. Ele apar in regnul animal, in special in sperma pestilor, cuplate cu acizi nucleici sub forma de nucleoproteide. Protaminele izolate din diferitele celule spermatice ale pestilor poarta denumiri diferite: salmina (somon), clupeina (scrumbie), ciprinina (crap), etc.
Prolamine si gluteline sunt holoproteide caracteristice regnului vegetal. Sunt proteine insolubile in apa, dar solubile in solutii diluate de acizi sau baze. Prolaminele sunt solubile si in alcool etilic 70 %, proprietate prin care se deosebesc de toate celelalte proteine.
In cantitati mai mari, prolaminele si glutelinele se gasesc in semintele de cereale. Astfel, in semintele de grau se gaseste prolamina numita gliadina si glutelina numita glutenina. Aceste doua proteine insotite de cantitati mici de albumine si globuline, alcatuiesc glutenul, care confera fainei de grau calitati de panificatie. Semintele de porumb contin prolamina numita zeina. Valoarea alimentara a acestor proteine este diferita in functie de continutul lor in aminoacizi esentiali. Astfel, protaminele sunt bogate in acid glutamic, glutamina si prolina, insa sunt sarace in triptofan si lizina, iar glutelinele sunt bogate atat in acid glutamic, cat si in lizina. Triptofanul si lizina sunt aminoacizi esentiali, care nu pot fi sintetizati, dupa cum am mai vazut, de catre organismul animal, insa sunt indispensabili. O alimentatie a omului si a animalelor, bazata exclusiv pe porumb, duce in scurt timp la tulburari de metabolism, care sunt agravate si din cauza continutului insuficient al boabelor de porumb in unele vitamine, in special in vitamina PP.
Scleroproteinele sunt holoproteide de natura fibrilara, caracteristice organismului animal. Ele se caracterizeaza prin rezistenta lor deosebita fata de diferiti agenti fizici sau chimici. Sunt insolubile in apa, in solutii de saruri, de acizi sau baze si nu sunt hidrolizabile. Datorita acestor proprietati, scleroproteinele exercita in organismul animal un rol de sustinere, de protectie si de rezistenta mecanica. Sunt constituentele principale a tesuturilor conjunctive si a epidermei.
Scleroproteinele au un continut relativ scazut in azot (cca. 12 %) si ridicat in oxigen. Aminoacizii care predomina sunt: cisteina, prolina si hidroxiprolina. Din categoria scleroproteinelor fac parte colagenele, keratinele, elastinele, fibroina, miozina si fibrinogenul.
Colagenele sunt proteinele principale din tesuturile conjunctive, tendoane, cartilagii, piele, oase si solzi de peste si reprezinta cca. 50 % din totalul proteinelor organismului animal. Sunt substante insolubile in apa si rezistente la actiunea enzimelor proteolitice. Prin fierbere cu apa, colagenele sufera o hidroliza partiala, care are loc cu modificari structurale, datorita desfacerii legaturilor intercatenare, transformandu-se intr-o masa elastica, numita gelatina.
Gelatina este solubila in apa calda, dand solutii vascoase, care prin racire formeaza geluri. Este hidrolizata de catre enzimele proteolitice.
Colagenele sub actiunea taninurilor se transforma in produse rezistente, neputrescibile, datorita supercontractiei ireversibile a fibrelor. Acest proces joaca un rol fundamental in tabacirea pieilor.
Keratinele sunt proteinele principale din epiderma, par, pene si din formatiunile cornoase (unghii, copite, coarne) si indeplinesc un rol protector. Ele sunt insolubile si rezistente fata de agentii chimici. Continutul ridicat in aminoacizi cu sulf (3-5 % S) cu aparitia unui numar mare de punti disulfidice - S - S - inter- si intracatenare, le confera rezistenta mecanica si elasticitate deosebita. Astfel, firul de par tratat cu apa calda si supus tractiunii sufera o alungire pana la 100 %. Keratinele nu sunt hidrolizate de catre enzimele proteolitice.
Elastinele sunt scleroproteide, ce participa la formarea fibrelor elastice din artere si tendoane. Se aseamana cu colagenele, insa nu se pot transforma in gelatine si se caracterizeaza printr-un procent ridicat de glicocol si leucina in detrimentul prolinei si hidroxiprolinei.
Fibroina este proteina principala din matasea naturala produsa de viermele de matase (Bombix mori) unde este asociata cu o alta proteina - sericina. In glandele viermelui de matase, proteinele se gasesc sub forma unei mase vascoase, care la trecerea prin orificiul ingust al glandei, ia aspectul de fir care este si rasucit de catre labutele viermelui
Fibroina este o proteina fibrilara tipica, structura fiind asemanatoare cu a β - keratinei (structura in planuri pliate), insa catenele polipeptidice sunt orientate antiparalel.
Miozina este principala proteina a tesutului muscular (35 %), avand proprietati contractile si enzimatice (fata de acidul adenozintrifosforic - ATP). Miozina este insotita de actina (17 %) cu care formeaza un complex numit actomiozina. Aceste constituie substanta de baza a fibrilelor musculare si suportul biochimic al contractiei musculare. Miozina prezinta elasticitate deosebita, molecula ei fiind alcatuita din trei catene polipeptidice.
Fibrinogenul este o proteina fibrilara solubila de tipul α - keratinei prezenta in plasma sanguina, avand rol esential in procesul de coagulare a sangelui. In acest proces, fibrinogenul se transforma in fibrina care este insolubila. Transformarea fibrinogenului in fibrina are loc in prezenta unor enzime (trombina, fibrinaza) si a ionilor de Ca2+.
Copyright © 2025 - Toate drepturile rezervate