|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
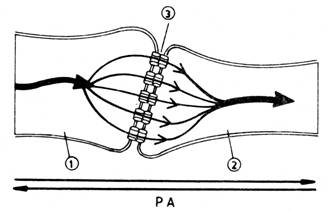
Functionarea sinapselor electrice. Sinapsele electrice sunt regiuni functionale in care
membranele celor doi neuroni sunt foarte apropiate (fig. nr. 9.5.) si legate prin canale
membranare de sodiu, caracteristice jonctiunilor celulare "gap". Aceste
canale permit trecerea libera a
curentului electric, astfel ca un PA
ajuns la o sinapsa electrica trece in neuronul urmator
fara intarziere la nivelul sinapsei. Cu cat suprafata de
contact sinaptic intre membrane este mai mare, cu atat schimburile de curent
electric intre componentele sinapsei vor fi mai mari. Sinapsele electrice, spre
deosebire de cele chimice, permit trecerea
influxului nervos in ambele sensuri, neexistind deosebiri structurale
majore intre cele doua componente sinaptice.
Fig. nr. 9.5. - Sinapsa electrica: 1 - neuron presinaptic; 2 - neuron postsinaptic
Sinapsele electrice sunt caracteristice sistemului nervos al nevertebratelor si cea mai importanta functie a lor este sa sincronizeze neuronii, care in mod normal sunt activati deodata. In increngatura vertebratelor, le intalnim in organele electrice ale unor pesti sau intre neuronii care activeaza in mod sincron aripioarele la pestii zburatori, dar si la vertebratele superioare si chiar la om, in nucleii vestibulari din bulb, unde se cer reactii nervoase foarte rapide, necesare mentinerii sau restabilirii pozitiei corpului in spatiu.
Functionarea sinapselor chimice Presupune traducerea PA in semnal chimic la nivelul componentei presinaptice, transmiterea in aceasta forma la componenta postsmaptica, urmata de regenerarea PA in neuronul postsinaptic (fig. nr. 9.6.).
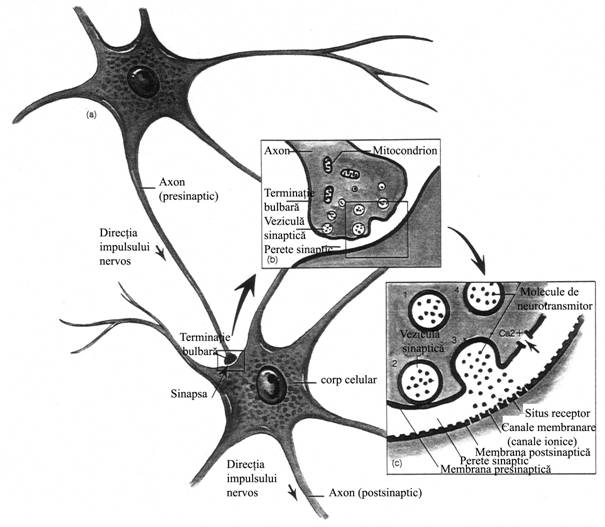
Fig. nr. 9.6. - Transmiterea impulsului nervos in sinapse
Eliberarea mediatorului chimic din componenta presinaptica incepe odata ce la membrana butonului terminal ajunge PA, condus de-a lungul axonului neuronului presinaptic. Depolarizarea membranei butonului terminal produce deschiderea canalelor voltaj-dependente pentru Ca++ si ionii de calciu patrund din spatiul extracelular in buton, in virtutea gradientului electrochimic. Aceasta determina fuzionarea veziculelor ce contin mediator cu membrana presinaptica, in "zonele active" ale acesteia. Membrana veziculei si membrana presinaptica se rup la locul de fuzionare si mediatorul chimic este eliberat in fanta sinaptica, prin exocitoza. Cantitatea de mediator chimic eliberata este proportionala cu marimea influxului de calciu in butonul terminal. Numarul de molecule de mediator chimic eliberate dintr-o singura vezicula sinaptica poarta numele de cuanta de mediator. Succesiunea evenimentelor din componenta presinaptica care au drept rezultat eliberarea mediatorului chimic sunt ilustrate in fig. nr. 9.7.
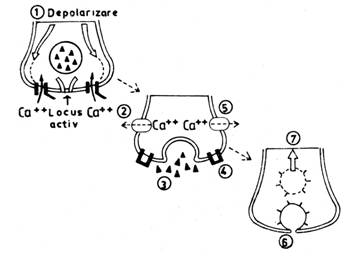
Fig. nr. 9.7. - Evenimente care au loc in membrana presinaptica. Depolarizarea (1) duce la deschiderea canalelor pentru Ca++ (2) care sunt situate in membrana presinaptica, in vecinatatea zonei active. Patrunderea calciului in butonul terminal determina fuzionarea veziculelor cu membrana presinaptica si eliberarea moleculelor de mediator chimic in fanta sinaptica (3). Canalele de calciu se inchid (4) si calciul din citoplasma este pompat in afara celulei (5). membrana veziculei in care s-a aflat mediatorul este recuperata in componenta presinaptica prin endocitoza (6) si primeste o noua cantitate de mediator, refacand vezicula sinaptica sau calatoreste spre soma prin transport axoplasmic retrograd (7).
Nu se cunoaste cu precizie mecanismul prin care patrunderea Ca++ in butonul terminal duce la fuzionarea veziculelor cu membrana postsinaptica. O ipoteza considera ca veziculele sunt ancorate de componente ale citoscheletului prin sinapsina, o proteina care inconjura veziculele. Patrunderea calciului in componenta presinaptica ar duce la fosforilarea acestei proteine si desprinderea ei de citoschelet, permitand veziculelor sa fuzioneze cu membrana presinaptica.
Moleculele de mediator chimic difuzate in fanta sinaptica sunt recunoscute de receptorii membranei postsinaptice si se fixeaza (o parte din ele) pe acestia. Receptorii, specifici pentru un anumit mediator, se pot incadra in una din urmatoarele doua categorii:
- receptori care fac parte din insasi structura canalelor ionice si
- receptori care sunt cuplati cu canalul ionic, printr-o proteina G si un sistem al celui de-al doilea mesager.
In receptorii asociati cu proteina G, prin fixarea mediatorului chimic pe receptor, este initiata o cascada de reactii enzimatice care da nastere unui al doilea mesager, ce modifica functiile celulare pentru o perioada mai lunga de timp decat o face o simpla deschidere a canalelor ionice. Aceste enzime legate de membrana postsinaptica si mesagerii secundari pe care ele ii produc in interiorul celulei sunt adenilatciclaza ce produce AMPc, guanilatciclaza ce produce GMPc si fosfolipaza C care poate produce doi mesageri secundari: diacilglicerolul (DAG) si inozitol trifosfatul (IP3).
In tabelul nr. 9.1. sunt prezentati cei mai frecventi mediatori chimici, impreuna cu receptorii lor, care actioneaza prin deschiderea directa a canalelor ionice sau prin formarea unui mesager secundar ori prin ambele mecanisme.
Tabelul nr. 9.1. - Tipuri de mediatori chimici si de receptori in sinapsele chimice
|
Mediatorul chimic |
Receptorul |
Modificari ale conductantei (g) membranar |
Modificari ale potentialului membranar |
Mesagerul secundar |
|
Acetilcolina |
Nicotinic Muscarinic M1 |
Creste gNa, gK Scade gK Creste gK |
PPSE PPSE PPSI |
IP3 si DAG AMPc |
|
Noradrenalina |
a1 a2 b2 |
PPSI (SNC) PPSI ? |
AMPc AMPc |
|
|
Dopamina |
D1 D2 |
PPSE ? PPSI |
AMPc AMPc |
|
|
Serotonina |
5-HT-IA 5-HT-IB 5-HT-IC 5-HT-ID 5-HT-2 5-HT-3 |
Creste gK Creste gCl Scade gx Creste gNa-gK |
PPSI PPSI PPSE PPSE |
AMPc IP3 IP3 |
|
Glutamat |
Kainat Quisqualat NMDA |
Creste gNa-gK Creste gNa, gK Creste gCa |
PPSE PPSE PPSE |
IP3 si DAG |
|
GABA |
GABA-A GABA-B |
Creste gCl Creste gK |
PPSI PPSI |
AMPc ? |
Cand un mediator chimic se fixeaza pe receptor, formand complexul ligand-receptor [MCh-R], apar modificari in conductanta membranei postsinaptice, care duc la depolarizarea sau la hiperpolarizarea acesteia. O crestere a conductantei pentru Na+ depolarizeaza membrana, in timp ce o crestere a conductantei care permite efluxul K+ sau influxul Cl- o hiperpolarizeaza. In anumite cazuri, hiperpolarizarea membranei postsinaptice poate apare si ca urmare a descresterii conductantei pentru Na+, ceea ce reduce influxul de ioni pozitivi.
Intre sosirea unui PA la componenta presinaptica si modificarea de potential in membrana postsinaptica se scurge o perioada de 0,5 - 0,7 ms, cunoscuta sub numele de intarziere sinaptica. Aceasta intarziere este data de timpul necesar pentru patrunderea Ca++ in butonul terminal si fuzionarea veziculelor avand mediator cu membrana presinaptica. Timpul necesar mediatorului pentru a difuza in spatiul sinaptic este neglijabil.
Odata depolarizata sau hiperpolarizata, membrana postsinaptica, canalele ionice din aceasta, se inactiveaza, inchiderea lor fiind determinata de incetarea actiunii mediatorului chimic. Actiunea mediatorului se opreste cand acesta este:
indepartat de pe receptor prin desfacerea enzimatica a complexului [MCh-R];
transformat chimic intr-o substanta inactiva;
transportat, prin mecanisme active, inapoi in butonul terminal sau, in unele cazuri, in celulele gliale invecinate acestuia.
Succesiunea acestor evenimente se desfasoara cu ajutorul unor enzime specifice; in final mediatorul chimic sau substanta rezultata din metabolizarea acestuia fiind recuperata prin endocitoza si transporta retrograd de neuronul presinaptic, iar apoi reutilizata pentru sinteza a noi cantitati de mediator.
Moleculele de mediator chimic care, dupa difuzarea in fanta sinaptica, nu au apucat sa se lege de receptorii postsinaptici vor avea aceeasi soarta ca si moleculele care s-au combinat cu receptorii.
Cele doua tipuri de sinapse chimice, excitatoare si inhibitoare se diferentiaza prin efectul mediatorului chimic asupra neuronului postsinaptic. Efectul excitator sau inhibitor depinde de tipul de receptori postsinaptici si de tipul de canale ionice pe care acesti receptori le controleaza. De aici rezulta ca este posibil ca acelasi mediator chimic sa produca efecte diferite, in functie de tipul de receptor cu care interactioneaza (deci de tipul de sinapsa), la o sinapsa avand rol de excitator, iar la alta rol de inhibitor. Astfel, neurotransmitatorul este doar un mesager chimic care declanseaza un raspuns preexistent in structura functionala a membranei postsinaptice (tipul de receptori si de canale ionice).
Sinapse chimice excitatoare. La o sinapsa excitatoare, raspunsul postsinaptic la mediatorul chimic este o depolarizare, care aduce potentialul membranar mai aproape de pragul de depolarizare. Formarea complexului [MCh-R] deschide canalele ionice permeabile pentru Na+, K+ si alti ioni mici pozitivi, care astfel devin liberi sa se miste prin membrana, in functie de gradientul lor chimic si electric.
Datorita
distributiei sarcinilor de o parte si de alta a membranei in stare de
repaus, in cazul ionilor de sodiu gradientul chimic si electric vor
actiona sinergic, in sensul influxului de sodiu, in timp ce in cazul
potasiului gradientul electric este opus gradientului de concentratie.
Deschiderea canalelor de sodiu si de potasiu va duce la un influx masiv de
ioni de sodiu, si concomitent, la un eflux, mult mai redus, de ioni de
potasiu. Ca rezultanta, deplasarea neta a sarcinilor pozitive va fi
de la exteriorul spre interiorul neuronului postsinaptic, cu o usoara
depolarizare a membranei postsinaptice (fig. nr. 9.8.). Aceasta modificare de potential electric este un potential postsinaptic excitator
(PPSE), un PL care 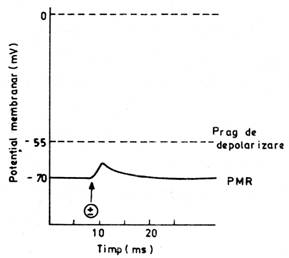 se conduce decremental de o parte si de alta a sinapsei, prin
membrana neuronului postsinaptic. Singura lui functie este aceea de a
aduce potentialul membranei post-sinaptice mai aproape de pragul de
depolarizare (la care se va putea initia un PA).
se conduce decremental de o parte si de alta a sinapsei, prin
membrana neuronului postsinaptic. Singura lui functie este aceea de a
aduce potentialul membranei post-sinaptice mai aproape de pragul de
depolarizare (la care se va putea initia un PA).
 Sinapse chimice inhibitoare. La sinapsele inhibitoare, modificarea de potential in neuronul
postsinaptic este de tip hiperpolarizant
sau de tip stabilizator. Deci, activarea unei sinapse inhibitoare scade
probabilitatea producerii unui PA in neuronul postsinaptic.
Sinapse chimice inhibitoare. La sinapsele inhibitoare, modificarea de potential in neuronul
postsinaptic este de tip hiperpolarizant
sau de tip stabilizator. Deci, activarea unei sinapse inhibitoare scade
probabilitatea producerii unui PA in neuronul postsinaptic.
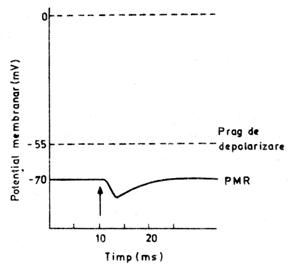 Formarea complexului [MCh-R] la nivelul membranei postsinaptice
deschide canalele de clor sau pe cele de potasiu, ori ambele tipuri; canalele
de sodiu nu sunt afectate. Asa cum s-a aratat intr-un alt capitol,
daca membrana celulara ar fi permeabila numai pentru potasiu, PMR
ar echivala potentialul de echilibru al acestui ion, adica ar fi de -90 mV in loc de -70 mV. Deci,
cresterea permeabilitatii membranei postsinaptice pentru K+
va duce la cresterea PMR si apropierea lui de potentialul de
echilibru al potasiului. Aceasta hiperpolarizare este denumita potential postsinaptic inhibitor (PPSI)
(fig. nr. 9.9.).
Formarea complexului [MCh-R] la nivelul membranei postsinaptice
deschide canalele de clor sau pe cele de potasiu, ori ambele tipuri; canalele
de sodiu nu sunt afectate. Asa cum s-a aratat intr-un alt capitol,
daca membrana celulara ar fi permeabila numai pentru potasiu, PMR
ar echivala potentialul de echilibru al acestui ion, adica ar fi de -90 mV in loc de -70 mV. Deci,
cresterea permeabilitatii membranei postsinaptice pentru K+
va duce la cresterea PMR si apropierea lui de potentialul de
echilibru al potasiului. Aceasta hiperpolarizare este denumita potential postsinaptic inhibitor (PPSI)
(fig. nr. 9.9.).
 In celulele care in stare de repaus exporta ioni de
clor prin mecanisme active, potentialul de echilibru pentru Cl-
(- 80 mV) este mai negativ decat PMR, deci cresterea
permeabilitatii pentru clor prin deschiderea canalelor sale sub
actiunea mediatorului chimic are un efect hiperpolarizant, similar cu cel
obtinut prin cresterea permeabilitatii pentru potasiu, se
produce de asemenea un PPSI. In celulele in care potentialul de echilibru
pentru clor este identic cu PMR, cresterea permeabilitatii
pentru clor stabilizeaza membrana la nivelul potentialului de repaus
si scade probabilitatea producerii unui PA (cu alte cuvinte,
initierea unui PA ar necesita un influx mai mare decat in mod normal de
ioni
In celulele care in stare de repaus exporta ioni de
clor prin mecanisme active, potentialul de echilibru pentru Cl-
(- 80 mV) este mai negativ decat PMR, deci cresterea
permeabilitatii pentru clor prin deschiderea canalelor sale sub
actiunea mediatorului chimic are un efect hiperpolarizant, similar cu cel
obtinut prin cresterea permeabilitatii pentru potasiu, se
produce de asemenea un PPSI. In celulele in care potentialul de echilibru
pentru clor este identic cu PMR, cresterea permeabilitatii
pentru clor stabilizeaza membrana la nivelul potentialului de repaus
si scade probabilitatea producerii unui PA (cu alte cuvinte,
initierea unui PA ar necesita un influx mai mare decat in mod normal de
ioni
de sodiu).
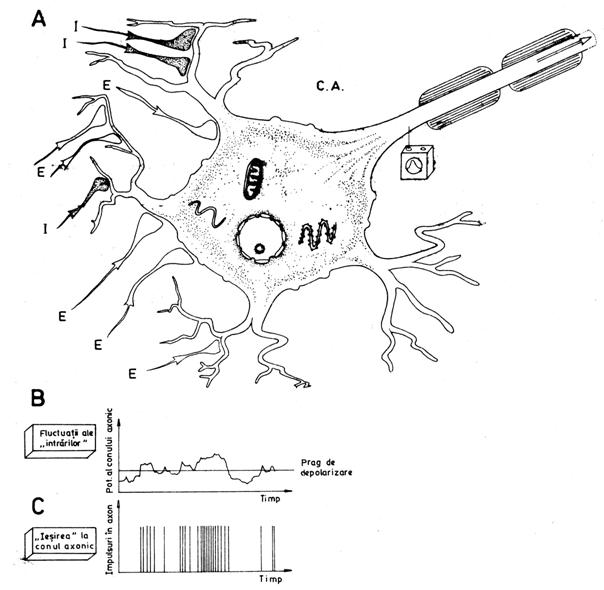
Fig. nr. 9.10. - A - Sumare spatiala si temporala a PPSE (sinapse excitatoare - E) si PPSI (sinapse inhibitoare - I). "Iesirea", care este rezultanta sumatiei, depinde de ponderea PPSE si PPSI. CA = con axonal. B - Un rezultat ipotetic al sumatiei. Fluctuatiile de potential ale membranei postsinaptice se datoresc impulsurilor excitatoare si inhibitoare pe care le primeste. Cand predomina impulsurile excitatoare, pragul de depolarizare este trecut si apar PA (reeprezentate ca linii verticale in C)
Activarea celulei postsinaptice. O caracteristica ce face posibila integrarea postsinaptica este faptul ca intrarea in activitate a unei singure sinapse excitatoare nu este suficienta pentru ca neuronul postsinaptic sa atinga pragul de depolarizare necesar initierii unui PA. Intr-un neuron postsinaptic motor, de exemplu, un PPSE are valoarea de 0,5 mV, dar pragul de depolarizare este de 15-25 mV. Astfel stand lucrurile, un PA poate fi initiat in neuronul postsinaptic doar prin efectul combinat al mai multor sinapse excitatoare. Dintre miile de terminatii sinaptice care vin la un neuron, cateva sute sunt active simultan (sau la intervale de timp suficient de apropiate ca efectele lor sa se poata insuma), iar potentialul membranar al neuronului postsinaptic este, in fiecare moment, rezultanta activitatii tuturor sinapselor care lucreaza in acel moment (fig. nr. 9.10.). Cand printre sinapsele active predomina cele excitatoare, are loc o depolarizare a membranei, numita facilitare, iar cand predomina activitatea sinapselor inhibitoare are loc o hiperpolarizare.
Pentru a vedea cum pot interactiona 2 PPSE, 2 PPSI sau un PPSE si un PPSI, sa ne imaginam un model teoretic in care un neuron postsinaptic poate primi influxuri nervoase de la 3 neuroni presinaptici (fig. nr. 9.11.); sinapsele A si B sunt excitatoare, iar sinapsa C este inhibitoare.
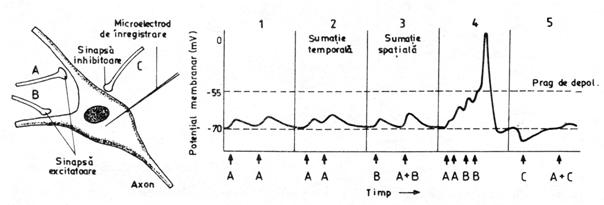
Fig. nr. 9.11. - Interactiunea PPSE si PPSI in neuronul postsinaptic. Sagetile reprezinta momentele de stimulare
(1) Vom testa interactiunea a 2 PPSE, produse prin stimularea axonului A, si apoi stimularea lui din nou, dupa un scurt interval de timp. Compartimentul 1 din figura arata ca nu apare nici o interactiune intre cele doua PPSE, deoarece slaba modificare a potentialului membranar produsa de prima stimulare se stinge in intervalul de timp care se scurge pana la a doua stimulare si cele 2 PPSE nu se pot insuma.
(2) Daca a doua stimulare a axonului A are loc inainte ca primul PPSE sa se stinga, al doilea PPSE se suprapune peste primul si depolarizarea rezultata in membrana postsinaptica este mai mare - se produce o sumatie temporala, stimulii ajungand in acelasi loc la timpi diferiti.
(3) Este stimulat axonul B pentru a determina amplitudinea PPSE produs de el, apoi sunt stimulati simultan axonii A si B. Cele doua PPSE care rezulta vor determina o depolarizare mai mare decat stimularea numai a lui B; are loc si in acest caz o sumatie, numita sumatie spatiala, deoarece cei doi stimuli actioneaza in pozitii diferite pe membrana postsinaptica a aceleiasi celule.
(4) Sumatia unor PPSE multiple poate duce membrana postsinaptica la pragul de depolarizare si aceasta permite initierea unui PA.
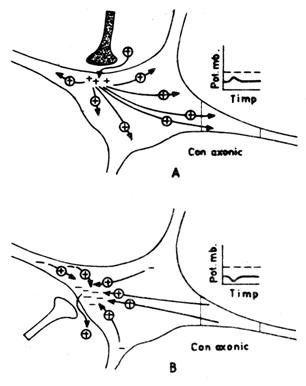 (5) Daca o sinapsa excitatoare si una inhibitoare
sunt activate in acelasi timp, PPSE si PPSI tind sa se anuleze
reciproc si modificarea de potential membranar este
nesemnificativa.
(5) Daca o sinapsa excitatoare si una inhibitoare
sunt activate in acelasi timp, PPSE si PPSI tind sa se anuleze
reciproc si modificarea de potential membranar este
nesemnificativa.
 PPSI se pot insuma,
spatial sau temporal, la fel ca si PPSE. Depolarizarea sau
hiperpolarizarea produsa in membrana postsinaptica se transmite, prin
mecanisme locale ("pas cu pas"), la intreaga membrana a somei neuronului
postsinaptic si pana la conul axonului (fig. nr. 9.12.). Ajunsi in acest punct, sa ne reamintim ca
pragul de depolarizare nu are aceeasi valoare peste tot de-a lungul
membranei neuronale; corpul celular si dendritele au un prag de
depolarizare de aproximativ 25 mV, iar conul axonic are cel mai scazut
prag, de 15 mV. De aceea, conul axonic atinge primul pragul de depolarizare,
cand are loc procesul de insumare al PPSE si PA care rezulta se
propaga de-a lungul axonului. In acest context, devine deosebit de
importanta pozitia sinapselor de-a lungul membranei neuronului
postsinaptic. PPSE produse la sinapsele localizate mai aproape de conul axonal
vor avea un impact mai mare in generarea PA, decat PPSE al sinapselor de pe
dendritele indepartate.
PPSI se pot insuma,
spatial sau temporal, la fel ca si PPSE. Depolarizarea sau
hiperpolarizarea produsa in membrana postsinaptica se transmite, prin
mecanisme locale ("pas cu pas"), la intreaga membrana a somei neuronului
postsinaptic si pana la conul axonului (fig. nr. 9.12.). Ajunsi in acest punct, sa ne reamintim ca
pragul de depolarizare nu are aceeasi valoare peste tot de-a lungul
membranei neuronale; corpul celular si dendritele au un prag de
depolarizare de aproximativ 25 mV, iar conul axonic are cel mai scazut
prag, de 15 mV. De aceea, conul axonic atinge primul pragul de depolarizare,
cand are loc procesul de insumare al PPSE si PA care rezulta se
propaga de-a lungul axonului. In acest context, devine deosebit de
importanta pozitia sinapselor de-a lungul membranei neuronului
postsinaptic. PPSE produse la sinapsele localizate mai aproape de conul axonal
vor avea un impact mai mare in generarea PA, decat PPSE al sinapselor de pe
dendritele indepartate.
PPSE dureaza mai mult decat PA. Daca insumarea PPSE poate mentine conul axonal in stare depolarizata prelungita dupa initierea unui PA si trecerea PRA a acestuia, atunci un nou PA este generat. Cu cat depolarizarea produsa prin insumarea PPSE este mai mare, cu atat frecventa PA ce au sansa de a se forma este si ea mai mare. De altfel, activitatea neuronala se desfasoara mai ales sub forma de trenuri (serii) de PA si mai putin prin PA izolate.
Din cele descrise pana acum, reiese ca un neuron poate afecta activitatea altui neuron numai la nivelul sinapselor. In anumite conditii (vezi si fig. nr. 9.1.), PL formate intr-un neuron pot influenta direct PMR ale neuronilor invecinati. Este cazul unor regiuni din SNC care contin un numar mare de prelungiri nervoase amielinice.
Eficienta sinapselor. Inhibitia sau facilitarea presinaptica. Eficienta cu care lucreaza o sinapsa in transmiterea PA cunoaste o mare variabilitate, care depinde atat de mecanismele presinaptice cat si de mecanismele postsinaptice.
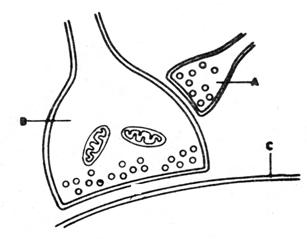 Mecanismele presinaptice
includ o mare varietate de factori care
modifica cantitatea de mediator eliberata de neuronul presinaptic (cu
cat cantitatea de mediator eliberat este mai mare, se deschid mai multe canale
in membrana postsinaptica si amplitudinea PPSE sau PPSI ce ia
nastere in neuronul postsinaptic este de asemenea mai mare).
Mecanismele presinaptice
includ o mare varietate de factori care
modifica cantitatea de mediator eliberata de neuronul presinaptic (cu
cat cantitatea de mediator eliberat este mai mare, se deschid mai multe canale
in membrana postsinaptica si amplitudinea PPSE sau PPSI ce ia
nastere in neuronul postsinaptic este de asemenea mai mare).
 De exemplu, Ca++
care a intrat in butonul terminal in timpul unui PA este indepartat prin
pomparea afara din celula sau (temporar) in unele organite celulare
(reticul endoplasmatic). Daca sistemele care au rolul de a indeparta
calciul nu sunt in stare sa tina pasul cu patrunderea
acestuia in celula, concentratia calciului in butonul terminal
si in consecinta, cantitatea de mediator chimic eliberat vor fi
mai mari decat in mod normal.
De exemplu, Ca++
care a intrat in butonul terminal in timpul unui PA este indepartat prin
pomparea afara din celula sau (temporar) in unele organite celulare
(reticul endoplasmatic). Daca sistemele care au rolul de a indeparta
calciul nu sunt in stare sa tina pasul cu patrunderea
acestuia in celula, concentratia calciului in butonul terminal
si in consecinta, cantitatea de mediator chimic eliberat vor fi
mai mari decat in mod normal.
Cantitatea de mediator chimic eliberat in fanta sinaptica este, de asemenea, modificata de activitatea receptorilor membranari ai membranei presinaptice (a butonului terminal). Acesti receptori pot sa fie sau sa nu fie asociati cu o a doua sinapsa, care se termina pe butonul terminal (sinapsa axo-axonica sau sinpsa presinaptica) (fig. nr. 9.13.). In cazul acestei sinapse, membrana butonului terminal B prezinta o regiune cu receptori, care joaca rol de membrana postsinaptica pe care se termina axonul A, si o regiune prin care elibereaza mediatorul chimic cu rol de membrana presinaptica in cadrul sinapsei B-C. Mediatorul chimic eliberat de butonul A se combina cu receptorii butonului B, generand potentiale in B si modificand astfel cantitatea de mediator chimic eliberat de butonul B ca raspuns la PA transmis prin axonul sau. Deci, neuronul A nu are un efect direct asupra lui C, dar poate influenta efectul neuronului B asupra lui C.
Spunem ca neuronul A exercita un efect presinaptic asupra sinapsei dintre neuronii B si C, putand diminua sau accentua eficienta acestei sinapse. In functie de natura mediatorului chimic eliberat de neuronul A si de natura receptorilor prezenti pe butonul terminal al lui B, putem vorbi de inhibitie presinaptica sau de facilitare presinaptica.
Un exemplu
clasic de inhibitie presinaptica il reprezinta celulele Renshaw din coarnele anterioare
medulare, prin care este modulata activitatea nervoasa a
motoneuronilor a
medulari (fig. nr. 9.14.). In coarnele anterioare ale
maduvei spinarii, neuronii senzitivi din ganglionul spinal, ce aduc
informatii de la receptorii periferici, stabilesc sinapse chimice
excitatoare cu motoneuronii a, ai caror axoni au drept organe tinta efectorii
musculari. Activitatea acestei sinapse excitatoare este controlata prin
intermediul celulei Renshaw (fenomen de
inhibitie presinaptica). Astfel, a-neuronul motor, printr-o colaterala
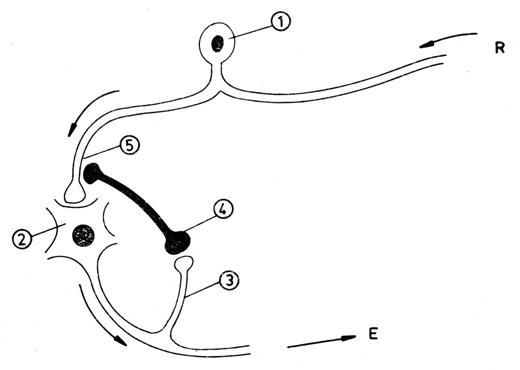
a axonului sau, sinapseaza cu neuronul Renshaw, al carui buton
terminal se leaga sinaptic de axonul neuronului senzitiv, printr-o
sinapsa axo-axonica. O activitate nervoasa sporita a
neuronului motor activeaza neuronul Renshaw, care emite mai multe cuante
de mediator chimic inhibitor (acidul gama-aminobutiric - GABA), ceea ce
genereaza hiperpolarizare si hipoexcitabilitate in axonul neuronului
senzitiv. Acesta va transmite mai putine PA neuronului motor, a carui
activitate va fi diminuata.
Fig. nr. 9.14. - Mecanismul inhibitiei prin intermediul celulei Renshaw. In coarnele anterioare ale maduvei spinarii, axonul neuronului senzitiv (1) din ganglionul spinal se afla in legatura sinaptica cu motoneuronul (2). Acesta, printr-o colaterala (3) a axonului sau, sinapseaza cu neuronul Renshaw (4), al carui axon elibereaza la sinapsa (5) cu neuronul senzitiv un mediator chimic inhibitor (GABA). R = receptor; E = efector.
Mecanismul inhibitiei presinaptice la nivel medular are o importanta functionala deosebita; din numarul mare de excitatii care sosesc la un neuron motor, prin inhibitie presinaptica sunt selectate informatiile importante si inhibate cele cu semnificatie redusa, evitandu-se astfel suprasolicitarea neuronului motor si pierderea distinctiei informatiei.
Copyright © 2025 - Toate drepturile rezervate