|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
NEURONUL
Neuronul este considerat unitatea morfo-functionala a tesutului nervos. Sistemul nervos uman insumeaza un numar de 1010-1011 neuroni, care nu au capacitate de diviziune.
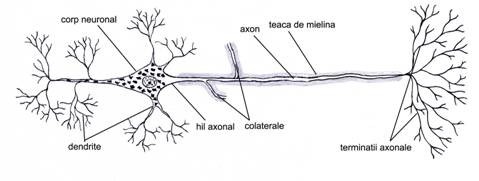
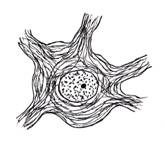
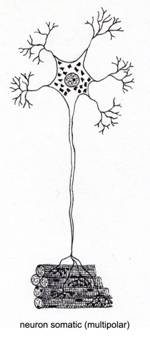
Din punct de vedere morfologic (fig. IV.1.1), neuronul prezinta un corp celular cu forme diferite si prelungiri citoplasmatice, cu lungimi diferite. Aceste prelungiri sunt de doua tipuri:
aferente, numite dendrite - de cele mai multe ori multiple, scurte si ramificate; au rolul de a transmite impulsul nervos de la periferie la corpul celular;
eferente, numite axoni - prelungiri unice lungi ce emit colaterale de-a lungul traiectului si care in portiunea terminala realizeaza frecvent o arborizatie numita telodendron, la nivelul careia se realizeaza sinapsa; au rolul de a conduce impulsul nervos de la corp la sinapsa.
Prelungirile citoplasmatice axonale si dendritice sunt numite generic cilindrax.
CORPUL NEURONULUI
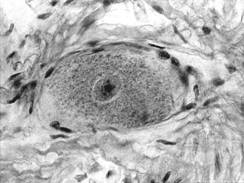
Corpul neuronului (pericarion sau soma) are dimensiuni diferite (50-150 μm) si forme variate si contine un nucleu unic, situat central (exceptie fac unii neuroni din ganglionii autonomi, care sunt binucleati).
Citoplasma este abundenta in neuronii mari si mijlocii si in cantitate redusa in neuronii mici.
|
|
|
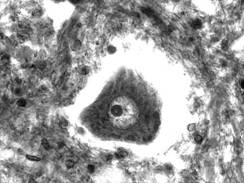
Fig. IV.1.1. Neuron - organizare generala |
|
|
|
Fig.
IV.1.2. Neuron - corp celular, |
|
|
|
Fig.
IV.1.3. Neuron - corp celular - ultrastructura |
In citoplasma sunt prezente uneori mici incluzii lipidice, care fie reflecta un metabolism celular anormal, fie constituie un material energetic de rezerva. Pe masura avansarii in varsta, apar si incluzii de lipofuscina, cu contur neregulat, probabil rezultat al activitatii enzimelor lizozomale; acestea pot deveni din ce in ce mai mari, ocupand un spatiu important in citoplasma, in detrimentul nucleului si al organitelor celulare. In neuronii cu caracter secretor (neurosecretori) citoplasma contine granule secretoare cu ultrastructura caracteristica, in raport cu natura neurosecretiei.
Astfel
au fost identificate vacuole cu miez electron-dens si dimensiuni pana
la 120 nm (catecolamine), vacuole cu continut electron-clar
si dimensiuni pana la 30 nm (vasopresina, oxitocina)
si vacuole electron-dense omogene de dimensiuni medii (peptide biologic
active: VIP,
colecistokinina etc.). In general glicogenul nu este evidentiabil
prin metode histochimice.
In unele zone (locus ceruleus, substanta neagra, nucleul motor dorsal al nervului vag, maduva spinarii, ganglionii simpatici) citoplasma contine granule de melanina (culoare brun-negru), a caror semnificatie nu este elucidata; in aceste zone au fost evidentiate, de asemenea, mici cantitati de feritina.
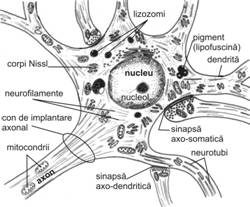
Nucleul (fig. IV.1.2, fig. IV.1.3) este rotund si palid, contine cromatina in cea mai mare parte fin dispersata, iar la periferia lui exista mici cantitati de heterocromatina. In unele cazuri, cromatina sexuala apare evidenta, langa membrana nucleara. Nucleolul (fig. IV.1.2, fig. IV.1.3) este bine reprezentat, aproximativ in centrul nucleului, motiv pentru care in histologia clasica apare comparatia cu "ochiul de bufnita". Aceasta organizare a nucleului este prezenta in neuronii in care se desfasoara o activitate de sinteza.
Exista insa si neuroni cu nuclei mai mici, cu cromatina condensata (de exemplu in stratul granular din cerebel).
In cazul unor leziuni la nivelul prelungirii axonale, suferinta corpului neuronului se manifesta initial prin deplasarea nucleului la periferia acestuia.
Microscopia electronica (fig. IV.1.3) evidentiaza organite celulare, in general bine reprezentate si dispersate. In mod traditional ele au fost impartite in organite comune si organite specifice.
Organitele comune sunt reprezentate de:
complex Golgi (demonstrabil in microscopia optica prin impregnare cu saruri de osmiu), localizat perinuclear, sub forma de retea, este alcatuit din grupe de saci, cisterne si vacuole golgiene; cisternele sunt dilatate periferic, fapt ce indica o intensa activitate de sinteza;
mitocondrii numeroase, dispersate aproximativ omogen, alungite, filamentoase, bogate in creste mitocondriale (care frecvent sunt aranjate longitudinal); prezinta uneori granule dense de matrice; ele au capacitatea de a se misca de-a lungul citoscheletului;
centrozom, prezent mai frecvent sub forma unui singur centriol, asociat unui corpuscul bazal ciliar (considerat structura vestigiala fara motilitate);
peroxizomi si lizozomi;
reticul endoplasmatic neted relativ abundent, reprezentat prin cisterne anastomozate care formeaza adesea o retea; uneori aceste cisterne se gasesc la periferie, sub membrana plasmatica (cisterne hipolemale); contine in interior calciu si material proteic, fiind considerat un sistem de conducere a substantelor sintetizate.
Organitele considerate in mod traditional specifice sunt corpii Nissl si neurofibrilele.
Corpii Nissl sau substanta tigroida (fig. IV.1.4, fig. IV.1.5) apar ca zone de material cromofil, pus in evidenta cu coloratii speciale (intens bazofil cu albastru de toluidina si metacromatic cu tionina), rezultand astfel un aspect patat care lipseste in zona de emergenta a axonului (hilul axonal). In contrast de faza, corpii Nissl pot fi observati fara colorare. Tratarea sectiunilor histologice, inaintea colorarii, cu ribonucleaza, impiedica aparitia corpilor Nissl, ceea ce a dus la concluzia ca aceste structuri constituie arii citoplasmatice bogate in ARN. Microscopia electronica a pus in evidenta faptul ca in zonele corespunzatoare corpilor Nissl exista o mare densitate de cisterne de reticul endoplasmatic rugos, ribozomi liberi si polizomi; printre aceste zone se gasesc reticul endoplasmatic
|
|
|
Fig. IV.1.4. Corpi Nissl - schema in MO |
|
|
|
Fig.
IV.1.5. Neuron cu corpi Nissl evidenti |
|
|
|
Fig. IV.1.6. Neurofibrile - schema in MO |
neted si alte organite. Corpii Nissl nu reprezinta deci organite specifice, ci numai o forma particulara de organizare intracelulara a RER.
Fara a exista un tipar riguros, dimensiunile corpilor Nissl difera in raport cu tipul de neuroni, putand fi mici si granulari, sau mai mari; aceste dimensiuni nu se coreleaza direct cu dimensiunile neuronilor.
Nucleolul evident, cromatina nucleara in cea mai mare parte sub forma dispersata si abundenta RER atrag atentia asupra importantei activitati de sinteza proteica pe care o desfasoara neuronii; cu cat un neuron este mai implicat in sinteza proteica, cu atat corpii Nissl sunt mai evidenti. Stimulari intense si prelungite la nivelul neuronului sau leziuni ale prelungirilor axonale (de exemplu sectionari) sunt urmate de stergerea pana la disparitie a corpilor Nissl, fenomen numit cromatoliza sau tigroliza. Reaparitia progresiva a corpilor Nissl traduce un proces de regenerare axonala.
Neurofibrilele
(fig. IV.1.6) au fost identificate pe preparate histologice tratate prin
impregnari metalice, sub forma unor structuri colorate in negru-cafeniu,
dispuse sub forma unei retele care ocoleste nucleul si cu un
aranjament ordonat, de tip paralel, in lungul prelungirilor neuronale. Aceste
structuri au diametrul intre 1-2 μm.
In microscopia electronica nu sunt evidentiate fibrile omogene
si compacte, de dimensiunile celor decelabile in microscopia optica;
in schimb, apare evidenta prezenta unui citoschelet neuronal foarte
bine organizat, format din trei tipuri de elemente: microtubuli, microfilamente
de actina, neurofilamente.
Microtubulii au diametrul intre 20-30 nm si, din punct de vedere molecular, sunt organizati similar cu microtubulii din alte celule; ei sunt constituiti prin polimerizarea moleculelor de tubulina (α si β in succesiune) cu molecule de dineina. La nivelul neuronului au fost descrise si trei tipuri de proteine asociate (MAP-1, 2, 3), cu rol in asamblarea si stabilizarea microtubulilor; dintre acestea, MAP-2 este localizata in citoplasma.
Microfilamentele de
actina, cu diametru de
6 nm, rezulta prin polimerizarea actinei G. Unele dintre aceste filamente
sunt ancorate pe fata interna a membranei plasmatice, prin molecule
de fodrina.
Neurofilamentele (filamente intermediare) sunt formate prin polimerizarea unor heterodimeri, organizati helicoidal, care determina existenta unei zone centrale de tipul unui lumen de circa 3 nm diametru.
Se accepta astazi ca neurofibrilele din microscopia optica reprezinta o conglomerare artefactuala a acestor elemente de citoschelet, in special a neurofilamentelor care au capacitatea de a fixa sarurile de argint. Citoscheletul neuronului are rol in mentinerea si modificarea formei celulei, in suportul organitelor si distributia lor, precum si in transportul unor componente moleculare sau chiar al unor organite de-a lungul prelungirilor.
Se poate deci conchide ca organitele considerate "specifice" in histologia clasica reprezinta, de fapt, modalitati particulare de organizare a anumitor componente celulare, perfect adaptate functionalitatii diferitilor neuroni.
|
Observatie |
DENDRITELE
Dendritele reprezinta procese sau expansiuni celulare aferente, cu rol receptor pentru informatia provenita din mediul extern sau intern. Astfel, dendritele preiau impulsul nervos de la celule senzoriale periferice, axoni sau dendrite apartinand altor neuroni, pe care il transmit inspre corpul neuronului. Uneori, impulsul nervos poate fi directionat spre alte dendrite.
In majoritatea cazurilor (fig. IV.1.1) ele sunt dispuse in apropiere de corpul celular si sunt multiple, marind substantial suprafata receptorie neuronala. In punctul de emergenta diametrul dendritei este mai mare decat al unui axon; ulterior, dendrita se ramifica intens, devenind arborizata; prin bifurcatii repetate, diametrul transversal scade progresiv inspre terminatia dendritei.
Nu exista un tipar comun dupa care se realizeaza arborizarea, fiecare tip de neuron avand elemente particulare. Aceasta morfologie permite ca, prin intermediul dendritelor, un neuron sa receptioneze concomitent stimuli multipli. Astfel, potentialul de membrana creste peste valoarea de prag.
Prin
tehnici de impregnatii metalice, de-a lungul dendritelor se
evidentiaza mici structuri denumite "spini dendritici", considerate a fi expresia morfologica a sinapselor. Numarul acestora poate
fi deosebit de mare (circa 20.000
la o celula piramidala); se remarca o scadere relativa
odata cu varsta si in unele boli genetice
(trisomia 13, trisomia 21).
Electrono-microscopic, dendritele contin toate elementele prezente in pericarion, cu exceptia aparatului Golgi, dar numarul acestora diminueaza pe masura subtierii dendritei. RER nu este evidentiabil sub forma corpilor Nissl, fiind dispus ca cisterne dispersate, scurte sau ramificate. Elementele de citoschelet capata progresiv o organizare paralela in lungul dendritei (fiind prezenta si proteina asociata MAP-2); neurofilamentele sunt reduse, apar in fascicule mici sau izolate, cuplandu-se in maniera incrucisata cu microtubulii. Dendritele nu prezinta teaca de mielina.
AXONUL
Axonul reprezinta prelungirea unica eferenta a neuronilor (fig. IV.1.1), cu rol efector, conducand impulsul nervos de la corpul celular spre alti neuroni sau spre celule efectorii.
In
general, axonii au diametrul intre
0,02-0,20 μm si lungimi variabile (de la cativa milimetri
pana la un metru); spre deosebire de dendrite, axonul isi
pastreaza dimensiunile diametrului pe toata lungimea sa,
realizand o structura cilindrica. Membrana plasmatica a axonului
este denumita axolema, iar citoplasma - axoplasma.
Zona de emergenta a axonului, numita si hilul axonului sau conul de implantare, este lipsita de corpi Nissl. De asemenea, de-a lungul axonului nu exista corpi Nissl, dar sunt prezente neurofibrile dispuse paralel si longitudinal.
Electrono-microscopic
se constata prezenta de mitocondrii alungite, scurte cisterne de
reticul endoplasmatic neted si vacuole. Elementele de citoschelet au
organizare longitudinala (fiind prezenta si proteina
asociata MAP-3) si, posibil, sunt implicate in reglarea diametrului
axonal; initial, la punctul de origine al axonului, microtubulii
formeaza fascicule mici, pentru ca
de-a lungul axonului sa se dispuna individualizat, spatiati
la distante egale si separati de neurofilamente.
Segmentul initial al axonului, localizat subjacent apexului hilului, este ceva mai ingust si lipsit de teaca de mielina (care va fi descrisa ulterior).
La acest nivel, peste axolema se constata un material fin granulos electron-dens, a carui functie nu este inca stabilita si al carui corespondent apare in zona nodurilor Ranvier (care vor fi descrise ulterior).
In aceasta zona ia nastere potentialul de actiune, ca urmare a stimularii membranei pericarionului prin impulsuri transmise de la nivelul dendritelor sau descarcate direct la nivelul pericarionului.
Potentialul de actiune va avansa centrifug de-a lungul axonului, axolema reprezentand astfel suportul morfologic al propagarii impulsului nervos spre alti neuroni, celule musculare sau elemente glandulare.
De-a lungul axonului pot aparea ocazional ramificatii colaterale, desprinse in unghi drept, care ulterior se orienteaza fie in directia terminatiei axonale, fie in sens invers (colaterale recurente).
Portiunea terminala a axonului se ramifica frecvent, formand telodendronul, iar extremitatea fiecarei terminatii este marcata de o mica dilatatie - butonul terminal, structura implicata in organizarea unei sinapse. (Exceptie fac celulele amacrine din retina - neuroni de asociatie - lipsite de axon.)
Axolema poate fi inconjurata de o structura elaborata prin participarea anumitor celule gliale - teaca de mielina (vezi sectiunea 2.1).
Neuronii care prezinta o astfel de teaca se numesc neuroni mielinizati (caracterizati printr-o viteza mare de conducere a impulsului nervos), iar cei lipsiti de aceasta teaca - neuroni amielinizati (la nivelul carora viteza de conducere a impulsului nervos este mult mai mica).
Fluxul axonal sau transportul axonal
Mentinerea integritatii morfo-functionale si a organizarii moleculare, precum si asigurarea comunicarii interneuronale presupun un transport continuu de-a lungul axonilor.
Acest transport este necesar deoarece pericarionul, prin prezenta RER si a ribozomilor, reprezinta sediul sintezei moleculelor responsabile de comunicarea intercelulara - axonul fiind lipsit de capacitate de sinteza.
Situatia este valabila in special la nivelul prelungirilor foarte lungi, exemplu fiind motoneuronii lombosacrati, ai caror axoni pot atinge un metru.
|
Observatie |
Dupa directia miscarii (a transportului moleculelor) se distinge un flux axonal anterograd (spre terminatia axonului) si un flux axonal retrograd (in sens invers). Dupa viteza transportului se diferentiaza un flux axonal rapid si un flux axonal lent; aceste doua tipuri de flux axonal sunt corelate cu diametrul axonului. Fluxul axonal rapid are o viteza intre 20 si 400 mm / zi si este bidirectional (in sens anterograd si retrograd); este caracteristic pentru axonii cu diametru mare. Fluxul axonal lent este unidirectional (in sens anterograd), atingand 0,2-8 mm / zi; este caracteristic pentru axonii cu diametru mic.
Prin fluxul axonal rapid anterograd sunt transportate, in principal, organite celulare (mitocondrii, vezicule, mici segmente de REN), molecule de actina, miozina si clatrina, anumite enzime implicate in sinteza neurotransmitatorilor, precum si substante cu greutate moleculara mica (aminoacizi, glucide, nucleotide, calciu).
Experimental, prezenta acestui flux a putut fi demonstrata prin injectare de aminoacizi radiomarcati si realizarea de autohistoradiografii. Acest transport este asigurat de catre molecule de kinezina care (ca si miozina), posedand proprietati de ATP-aza, interactioneaza ciclic cu microtubulii, atasandu-le elementele pentru transport. Conform datelor din literatura de specialitate, kinezina realizeaza astfel o viteza de transport de 3 μm/s. S-a demonstrat ca microtubulii axonali prezinta o polaritate caracteristica, manifestata prin adaugarea permanenta de dimeri de tubulina la extremitatea lor terminala, in echilibru cu o depolimerizare a tubulinei la extremitatea proximala (corpul celular). Acest fenomen este posibil prin transportul tubulinei din pericarion spre terminatiile axonale.
Prin fluxul axonal rapid retrograd are loc un transport de neurofilamente agregate, fragmente de microtubuli, enzime solubile, lizozomi, factori neurotrofici (produsi de celule tinta), virusuri neurotrope si neurotoxine.
Experimental, prezenta acestui flux a putut fi demonstrata prin injectare de peroxidaza de hrean la periferie si evidentierea ulterioara a acesteia, prin reactii histochimice, in corpurile neuronale. Fluxul retrograd este asigurat prin molecule de dineina care, la fel ca si kinezina, are activitate ATP-azica si ataseaza moleculele transportate pe microtubuli.
Prin
fluxul axonal lent anterograd se
transporta, in principal, componentele moleculare ale citoscheletului
(tubulina, actina, proteinele care formeaza neurofilamentele).
Bazele moleculare ale acestui transport sunt mai putin cunoscute.
O ipoteza se bazeaza pe proprietatea de polarizare prezentata
mai sus, conform careia continua depolimerizare a elementelor de
citoschelet la extremitatea proximala, in paralel cu polimerizarea
extremitatii distale, asigura o deplasare continua a
intregului citoschelet.
Dupa o alta ipoteza, fluxul lent este comparabil cu ceea ce se intampla la nivelul neuronilor, in cadrul procesului de fibrilogeneza. Fibrilogeneza implica cresterea si alungirea axonilor si se realizeaza pe seama microtubulilor. Acestia sunt conectati prin punti proteice formate din dinamina care, in prezenta ATP-ului si probabil a unui cofactor, genereaza energia necesara pentru glisarea lor; consecutiv, are loc alungirea axonului in crestere. Cea de a doua ipoteza nu a fost pe deplin verificata la neuronul matur. Prin intermediul fluxului axonal lent se transporta si elemente componente ale matricei citoplasmatice (actina, calmodulina, alte enzime metabolice).
Existenta unui flux dendritic este acceptata, dar modalitatea de realizare a acestuia inca este controversata.
O teorie sustine aceleasi caracteristici pentru fluxul dendritic, ca in cazul fluxului axonal. O alta teorie considera fluxul dendritic ca flux lent anterograd, in baza constatarii unei polaritati mixte a microtubulilor la nivelul dendritelor, fapt ce asigura transportul centrifug al ribozomilor si al unor mici componente golgiene.
|
|
|
|
| |
CLASIFICAREA NEURONILOR
Structura generala a neuronilor este similara. Exista insa o anumita diversitate, legata in mare masura de distributia si functia neuronilor in cadrul sistemului nervos. In consecinta, neuronii pot fi clasificati pe baza mai multor criterii morfologice si functionale, pe care le prezentam in continuare.
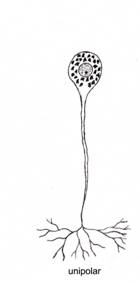
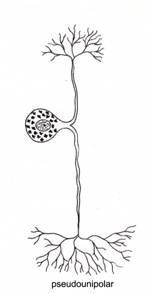
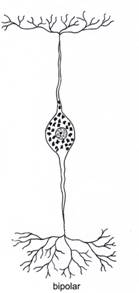
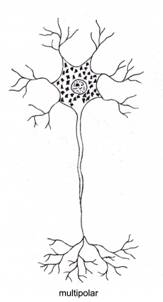
Dupa morfologia generala a neuronului, exista neuroni unipolari, pseudounipolari, bipolari si multipolari (fig. IV.1.7).
Neuronii unipolari prezinta o singura prelungire, de tip dendritic. Sunt prezenti in numar mare in timpul histogenezei tesutului nervos (precoce la embrion), iar in tesutul nervos matur ca celule amacrine, in retina.
Neuronii bipolari au doua prelungiri distincte, una aferenta si una eferenta, care se mentin separate una de cealalta. In aceasta clasa sunt inclusi neuronii de asociatie din retina, neuronii receptori din mucoasa olfactiva (care mentin caracteristici din perioada dezvoltarii precoce a tesutului nervos, avand o localizare superficiala, in contact cu mediul extern, precum si un potential - lent - de reinnoire) si neuronii din ganglionii vestibulari si cohleari (ai nervului VIII vestibulo-cohlear).
Neuronii pseudounipolari au originea in neuroni bipolari embrionari, prezentand initial doua prelungiri (aferenta si eferenta), care in cursul maturarii se apropie una de alta, fuzionand pe o anumita distanta. Ulterior, acestea se despart in unghi drept, rezultand, din nou, doua prelungiri. Prelungirea aferenta se orienteaza spre periferie, iar cea eferenta - spre sistemul nervos central. Ambele prelungiri au rol de conducere a impulsului nervos, astfel incat ele pot fi considerate axoni. Totusi, prelungirea aferenta realizeaza la terminatia ei distala ramificatii arborizate de tip dendritic, cu rol receptor si deci implicit de producere a impulsului. Transmiterea se realizeaza direct intre cele doua prelungiri, fara a traversa corpul celular. Neuronii pseudounipolari sunt localizati in ganglionii cerebrospinali.
Neuronii multipolari
(marea majoritate) prezinta numeroase prelungiri aferente (dendrite), cu
puncte de emergenta diferite si o prelungire eferenta
(axon). Impulsul nervos este transmis de la dendrite la corpul celular si
apoi la axon. Dendritele si corpul celular constituie teritoriile de
receptie a stimulilor, motiv pentru care membrana plasmatica din
aceste zone este adaptata pentru producerea impulsului. Axonul este
teritoriul de conducere a stimulilor, membrana sa plasmatica fiind
adaptata pentru conducerea impulsului.
In mod particular, pentru celulele Purkinje din cerebel, emergenta
dendritica este initial unica si groasa, dupa
care apare o intensa ramificatie
intr-un singur plan, asemanatoare iederei.
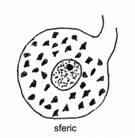
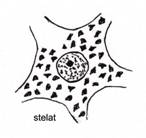
Dupa
morfologia corpului celular,
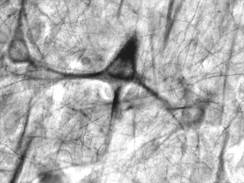
vizibila pe preparate histologice de rutina, sunt descrisi neuroni sferici, neuroni fuziformi, neuroni
stelati, neuroni piramidali (fig. IV.1.8,
fig. IV.1.9, fig. IV.1.10).
Dupa lungimea axonului, exista neuroni tip Golgi I (de exemplu, neuroni de proiectie, localizati in nucleii motori ai sistemului nervos central), care poseda un axon lung (pana la 1 m), si neuroni tip Golgi II (frecvent neuroni de asociatie), cu axon scurt.
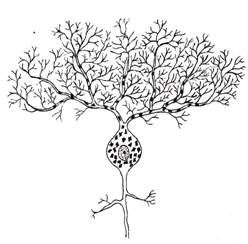
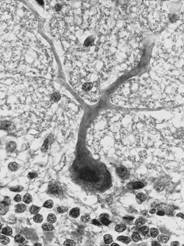
Dupa organizarea dendritelor, clasificarea include neuroni izodendritici, cu organizare spatiala a ramificatiilor dendritice in toate directiile, neuroni alodendritici, cu organizare asimetrica a ramificatiilor dendritice si neuroni idiodendritici, cu organizare particulara a arborizatiei dendritice (exemplu fiind celula Purkinje, mentionata anterior) (fig. IV.1.11, fig. IV.1.12).
|
|
|
|
|
|
|
Fig. IV.1.8. Tipuri de neuroni dupa morfologia corpului celular - schema in MO |
||||
|
|
|
|||
|
Fig.
IV.1.9. Neuron sferic |
Fig.
IV.1.10. Neuron piramidal |
|||
|
|
|
|||
|
Fig.
IV.1.11. Celula Purkinje - schema in MO |
Fig.
IV.1.12. Celula Purkinje |
|||
Dupa criterii functionale, se deosebesc neuroni motori, neuroni senzitivi, neuroni de asociatie, neuroni secretori. Neuronii motori au origine in sistemul nervos central si conduc impulsurile nervoase de la acest nivel la celulele efectoare (musculare, glandulare) sau la alti neuroni. Sunt considerati neuroni eferenti, iar prelungirile lor axonale intra in alcatuirea fibrelor nervoase eferente somatice (trimit impulsuri voluntare la muschii scheletici) si viscerale (transmit impulsuri involuntare la muschiul neted, celulele conducatoare cardiace si glande).
Neuronii senzitivi primesc si conduc impulsurile nervoase preluate prin terminatiile dendritice sau de la receptori spre sistemul nervos central. Sunt considerati neuroni aferenti, iar prelungirile lor axonale formeaza fibrele nervoase aferente somatice ale nervilor senzitivi sau micsti si tracturile ascendente din maduva spinarii (transmit impulsuri nervoase generate de stimuli nociceptivi, termici si presionali, la suprafata corpului sau in organe ale sistemului locomotor - muschi, tendoane, articulatii) si viscerale (transmit impulsuri nervoase generate de modificari de forma sau fizico-chimice la nivelul organelor interne). Neuronii de asociatie, denumiti si neuroni intercalari sau interneuroni, sunt prezenti exclusiv in sistemul nervos central si formeaza o retea vasta intre neuronii senzitivi si motori, responsabila de transmiterea si integrarea informatiei, asigurand functionarea complexa a organismului. Datele din literatura de specialitate estimeaza ca, din numarul total de neuroni, peste 99% sunt neuroni de asociatie. Neuronii motori si neuronii de asociatie sunt, morfologic, neuroni multipolari, iar neuronii senzitivi - neuroni unipolari sau bipolari.
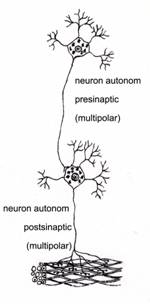
Dupa apartenenta la sistemul nervos somatic sau la cel autonom, exista neuroni somatici si neuroni autonomi (fig. IV.1.13).
|
||||
|
|
||||
|
|
||||
|
Fig. IV.1.13. Tipuri de neuroni dupa apartenenta la sistemul nervos - schema dupa (Ross, Kaye, Pawlina, 2003) |
||||
Copyright © 2025 - Toate drepturile rezervate