|







 |  |
|
| |
 | Alimentatie | Asistenta sociala | Frumusete | Medicina | Medicina veterinara | Retete |
Drojdia de panificatie reprezinta o biomasa de celule din specia Saccharomyces cerevisiae (drojdie de fermentatie superioara), biomasa formata din celule vii, capabile sa produca fermentarea zaharurilor din aluat cu formarea de alcool etilic si dioxid de carbon, agentul de afanare al aluatului si alte produse secundare, cu rol in formarea painii.
Cultivarea drojdiei Saccharomyces cerevisiae in scopul obtinerii de biomasa destinata industriei de panificatie este un complex de procese fizico-chimice, biochimice, termoenergetice si microbiologice.
Din productia mondiala de drojdie comprimata aproximativ 88% este folosita in industria panificatiei, iar restul pentru obtinerea de izolate proteice, vitamine (grupul B) sau enzime (invertaza, dehidrogenaza, enzime ale complexului zimazic), incat in diferite tari consumul mediu de drojdie este de 1,4÷2,5 kg/locuitor si an.
Scopul principal al tehnologiei de fabricatie a drojdiei de panificatie il reprezinta obtinerea unei cantitati maxime de biomasa de drojdie de calitate superioara cu consum minim de medii nutritive si de utilitati. Se urmareste realizarea unor multiplicari optime a celulelor prin inmugurire, folosind culturi periodic innoite, cu mentinerea conditiilor prescrise de dezvoltare si luarea in considerare a starii fiziologice, a cantitatii de drojdie cuib si a tuturor factorilor limitativi.
Industria drojdiei de panificatie din tara noastra a cunoscut o dezvoltare ampla pana in anul 1989, atat prin modernizarea fabricilor existente, imbunatatirea indicilor intensivi si extensivi de utilizare a utilajelor, cat si prin infiintarea a noi capacitati de productie.
In anul 1989 existau 6 fabrici de drojdie la Arad, Bucuresti, Oradea, Seini, Tandarei si Bacau. Dupa anul 1989, trei din aceste fabrici si-au incetat activitatea, fabrica de la Tandarei, definitiv prin scoaterea la licitatie a utilajelor proprii, iar cele de la Oradea si Seini, temporar, prin intrarea in conservare.
Incepand cu anul 1992, piata romaneasca a fost invadata de drojdie de panificatie importata din Turcia, Franta, Iugoslavia, Grecia, Bulgaria, astfel ca, datorita unei concurente neloiale si in lipsa unei protectii din partea statului productia de drojdie indigena a scazut foarte mult. In prezent, nici o fabrica din tara, din cele vechi, nu lucreaza la capacitatea ei zilnica.
In anul 1999 a fost data in functiune o noua fabrica de drojdie, la Pascani, cu capital integral turcesc, fabrica ce apartine concernului PAKMAYA - S.C. "ROMPAK" S.A.
Pe plan mondial, dintre firmele renumite din industria drojdiei de panificatie se pot enumera Pressindustrie - Italia, Vogelbusch si Andritz - Austria, Pasilac - Danemarca, Mauri - Australia, Pakmaya, Akmaya, Safmaya, Ozmaya - Turcia, Liko - Cehia, Lesaffre - Franta.
In industria drojdiei de panificatie se folosesc cateva scheme de obtinere a drojdiei de panificatie care se deosebesc prin procedeul tehnologic aplicat (clasic - discontinuu, semicontinuu, continuu), modul de folosire a materiei prime (cu plamezi diluate sau concentrate), numarul stadiilor de multiplicare, viteza de crestere, parametrii tehnologici utilizati (temperatura, pH, cantitatea de drojdie de insamantare), s.a.
Toate schemele existente prevad acumularea continua de biomasa.
Pe plan mondial, la baza schemelor tehnologice existente se afla aceleasi metode de cultivare, firmele producatoare de drojdie de panificatie introducand diferentele lor specifice in tehnologie sau sub aspectul utilajelor folosite. Aceste diferentieri conduc si la obtinerea de drojdii cu indici fizico-chimici diferiti, caracteristic fiecarei firme producatoare (tabelul 19).


Receptie
![]() Multiplicare
Multiplicare
in laborator
![]() Depozitare
Depozitare
![]()
![]() Multiplicare
Multiplicare
![]() Cantarire in faza I
Cantarire in faza I
![]()
![]()
![]() Diluare Multiplicare
Diluare Multiplicare
![]()
![]() in faza a II-a
in faza a II-a
![]() Acidulare
Acidulare
![]() Sterilizare
Sterilizare
Limpezire
![]()
Multiplicare
![]()
![]()
![]() in faza a
III-a
in faza a
III-a
![]()
![]()
![]() Multiplicare
Separare
Multiplicare
Separare
![]()
![]() in faza a
IV-a maia
in faza a
IV-a maia
![]()
![]()
![]()
![]() Multiplicare
Purificare
Multiplicare
Purificare
in faza a V-a cu H2SO4
![]()
![]() Separare
si spalare
Separare
si spalare

![]()
Racire
![]()
![]()
![]() Filtrare-presare Granulare
Filtrare-presare Granulare
![]()
![]()
![]() Malaxare Uscare
Malaxare Uscare
Modelare si ambalare Ambalare
![]()
![]()
Depozitare
COMPRIMATA

DROJDIE
Tabelul 19
Indicatorii masici si energetici ai procesului de obtinere a drojdiei de panificatie utilizati de firme reprezentative din Europa
|
Consum productie /1tona drojdie(27% s.u.) |
Firma producatoare |
||||
|
Pressindustria |
Vogelbush |
Andritz |
Pasilac |
||
|
Melasa(M50), kg | |||||
|
Sulfat de amoniu, kg | |||||
|
Apa amoniacala, kg | |||||
|
Diamoniufosfat, kg | |||||
|
Acid sulfuric, kg | |||||
|
Apa,m3 |
tehnologica | ||||
|
de racire | |||||
|
consum general | |||||
|
Energie electrica, kW/h | |||||
|
Abur, Gcal | |||||
In tara noastra, in majoritatea fabricilor de drojdie, drojdia de panificatie se obtine dupa schema tehnologica prezentata in figura 18 si cuprinde ca etape principale:
pregatirea melasei in vederea cultivarii drojdiei;
multiplicarea drojdiilor in cele cinci faze;
separarea drojdiilor din mediul de cultura;
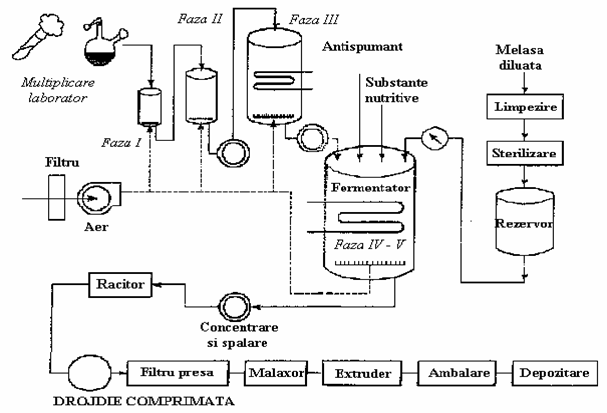
filtrarea-
presarea drojdiei;
modelarea si ambalarea drojdiei de panificatie - produs finit (fig.19).
Drojdia de panificatie se prezinta astazi, in comert, in mai multe forme diferite: drojdia comprimata (proaspata), drojdie uscata activa (ADY), drojdie uscata activa protejata (PADY) si drojdie uscata instant (IDY).
In prezent necesarul zilnic de paine in tara noastra este de circa 1000 tone. Pentru aceasta sunt necesare 180 tone de drojdie de panificatie. Capacitatea interna pentru productie este de 120 tone zilnic (24000 tone anual) data de fabricile producatoare de drojdie Arad, Bucuresti, Oradea, Seini si Bacau.
In S.U.A. productia drojdie de panificatie a fost organizata inca din 1868 si este disponibila astazi in mai multe forme diferite.
Cea mai populara forma este drojdia comprimata (proaspata), care se comercializeaza in pachete vrac ca drojdie sfaramata si ca drojdie pentru prajituri ambalata in hartie ceruita. Drojdia in calupuri pentru prajituri este folosita acum indeosebi in instalatii de coacere mai mici, pentru ca aceasta este in pachete de 0,5 kg sau de 2,5 kg, ori in cutii, pentru a usura procesul de cantarire in aceste mici brutarii (laboratoare). In timp ce drojdia comprimata este folosita in general intr-o suspensie de apa rece pentru a usura masurarea in sisteme automate de dozare, cea mai mare parte a drojdiei in calupuri se adauga direct in amestec fie sfaramata, fie in suspensie, sub forma de maia. Acest tip de drojdie este usor incorporata in aluat si incepe imediat fermentarea zaharurilor, chiar inainte ca aluatul sa fie complet framantat.
Drojdia pasta cu un continut de substanta uscata de 18% a fost disponibila in industria de panificatie din S.U.A. dupa anul 1980. Instalarea initiala a unui sistem pentru fabricarea drojdiei pasta poate costa de la 200.000 la 500.000$. Depozitata la temperatura recomandata de 20C cu o usoara agitare in tanc, drojdia pasta are o durata de pastrare de pana la 3 saptamani.
Drojdia uscata activa (ADY) a fost realizata in anul 1940 ca raspuns la nevoile speciale din timpul celui de-al doilea razboi mondial. Drojdia uscata activa nu necesita refrigerare in afara de cazul cand trebuie depozitata o perioada mai lunga. Daca este ambalata sub vid sau intr-o atmosfera inerta, drojdia uscata activa are o durata de pastrare de pana la 2 ani. In producerea sa se combina o tulpina speciala de Saccharomyces cerevisiae cu conditii specifice de crestere si un procedeu de uscare atent controlat. Drojdia uscata activa are un continut de proteine relativ scazut (38÷42%) si un continut ridicat de zaharuri (39÷47%). Pentru a obtine rezultate bune drojdia uscata activa trebuie rehidratata in apa calduta inainte de a se adauga in aluat. Producatorii de drojdie recomanda ca aceasta sa fie efectuata cu 4÷6 parti apa la 38÷43 C, pentru fiecare parte de drojdie uscata activa timp de 5÷10 minute. Raportat la substanta uscata drojdia uscata activa are o activitate echivalenta cu 65÷75% din cea a drojdiei proaspete. Motivul pentru care drojdia uscata se rehidrateaza in apa calduta este acela ca in timpul procesului de uscare membranele celulelor de drojdie pot deveni foarte poroase. Acestea pot fi refacute mai rapid in apa calda decat in apa rece, care incetineste procesul de rehidratare. Apa rece poate de asemenea sa determine solubilizarea pana la jumatate din componentele solubile din celulele de drojdie, care include glutationul. Glutationul solubilizat este un puternic agent reducator care nu numai ca reduce timpul de framantare, dar slabeste si structura glutenului din aluat. Aceasta poate duce la o reducere semnificativa a volumului specific al painii.
O forma stabila de drojdie uscata activa (ADY) este drojdia uscata activa protejata (PADY). Acest tip de drojdie a fost fabricat in 1960 ca ingredient pentru mixturi cu un continut de umiditate mai scazut si avand adaugati oxidanti si emulgatori. Emulgatorii usureaza rehidratarea drojdiei si astfel ajuta la reducerea solubilizarii componentelor celulare din drojdie. Durata de pastrare a drojdiei uscate protejate poate fi pana la de doua ori mai mare decat forma obisnuita neprotejata.
O alta forma de drojdie uscata cunoscuta ca drojdie uscata instant (IDY) a fost fabricata in 1960. Aceasta drojdie a fost rezultatul unei noi tulpini de Saccharomyces cerevisiae cu conditii de crestere si uscare diferite si al adaugarii de emulgatori. Se ambaleaza sub vid sau in atmosfera inerta si poate avea o durata de pastrare de un an la temperatura camerei. Raportat la substanta uscata, activitatea acesteia variaza de la 80 la 90% din cea a drojdiei proaspete. Drojdia uscata instant are un continut de umiditate de 5%. Un continut de proteine de 43÷44% impreuna cu circa 40% hidrati de carbon, nu numai ca asigura activitatea buna a drojdiei in aluat, dar are, de asemenea, o foarte buna stabilitate in timpul depozitarii in pachete nedeschise. Odata ce pachetul a fost deschis si drojdia uscata instant s-a expus la oxigenul din aer durata de pastrare a drojdiei se reduce substantial. Este foarte important ca drojdia uscata instant sa fie rehidratata, fie in apa calda (30÷430C), fie prin adaugarea sa la faina pe durata procesului de framantare. Particulele foarte fine ale acestei drojdii fac acest lucru posibil in majoritatea aluaturilor. Aluaturile foarte uscate, cum ar fi cele pentru paine bagheta, constituie totusi o exceptie si pot contine o umiditate insuficienta pentru rehidratarea drojdiei pe durata framantarii. Indiferent de metoda de adaugare folosita este important de reamintit ca drojdia uscata instant nu se adauga niciodata in apa rece. Solubilizarea glutationului din celulele de drojdie in timpul unei rehidratari necorespunzatoare poate avea ca efect o slabire semnificativa a structurii de gluten. In acelasi timp, aceasta furnizeaza avantajul potential al reducerii timpului de framantare al aluatului.
Pe langa tipurile de drojdie obtinute, producatorii de drojdie din S.U.A. pun la dispozitie drojdii speciale pentru produse specifice, cum ar fi: aluaturi fara zahar, aluaturi foarte dulci, arome (drojdii inactive)si altele.
In S.U.A. se folosesc si altfel de drojdii pentru prepararea painii. Astfel cultura microbiana folosita pentru producerea painii acide de San Francisco contine o drojdie specifica (Saccharomyces exiguus) care spre deosebire de drojdia de panificatie este incapabila sa fermenteze maltoza. Aceasta drojdie este capabila sa coexiste cu bacteria heterofermentativa Lactobacillus San Francisco care produce cativa acizi organici diferiti, pe langa acidul lactic la un nivel de pH 3,8÷4,5. Aceasta drojdie speciala pentru aluaturi acide este un excelent producator de dioxid de carbon gazos pentru crestere, dar nu tolereaza frigul la fel de bine ca Lactobacillus si ca drojdia de panificatie.
In S.U.A. se folosesc ca materii prime pentru fabricarea drojdiei, in afara de melasa si pseudomelasele care reprezinta siropuri cu compozitii asemanatoare cu cele ale melaselor, dar cu adaos de substante nutritive si de biostimulatori. In decembrie 1981 a fost creata firma Nutrisearch in scopul utilizarii zerului in producerea de drojdie de panificatie si proteine.
Fabricarea drojdiei de panificatie in fosta U.R.S.S. a cunoscut si cunoaste o dezvoltare continua prin aplicarea realizarilor din domeniul bioenergiei si eliminarea deficitelor din fabricile moderne de drojdie. Astfel, fabrica de drojdie din orasul Kurgansk produce 12.500 tone drojdie de panificatie pe an, din care 1.300 tone drojdie uscata. Specialistii fabricii au conceput o instalatie care purifica apa reziduala, reziduul se usuca si se foloseste in hrana animalelor.
Din 1976, functioneaza pe langa fabrica de zahar din Erken-Sahar o sectie de drojdie ce utilizeaza procedeul de multiplicare cu plamezi concentrate si produce 400 tone drojdie pe an.
Fabrica de drojdie din Riga cu o productie de 000 tone pe an prezinta cateva particularitati in procesul tehnologic: foloseste apa racita la temperatura de 50C in procesul tehnologic (pregatirea patului de pornire), iar alimentarea liniilor este automatizata (dozatoare); apa de la prima treapta de separare se foloseste la diluarea melasei, are un continut de 3÷4% substanta uscata, ceea ce duce la scaderea consumului de melasa.
Fabrica de drojdie din Moscova produce 22.500 tone drojdie pe an, in medie 66,5 tone pe zi. Din anul 1975 foloseste tehnologia de fabricare cu plamezi concentrate.
In Germania, printre cele mai mari intreprinderi s-au dezvoltat pana la al doilea razboi mondial, Norddeutsche, Hefeindustrie Berlin (astazi Deutsche Hefewerk GmbH). Prin evenimentele din timpul razboiului si dupa razboi majoritatea fabricilor au suferit din greu. In 1949 existau 20 de fabrici in Germania, iar in 1960, numarul lor a crescut la 27.
In Polonia existau in 1970, 8 fabrici de drojdie de panificatie, din care o fabrica producea si drojdie uscata.
In Suedia, printre fabricile de drojdie se numara si fabrica din Rotebro, construita in anul 1976. Fabrica are o capacitate de productie de 16.500 tone pe an, din care 5% sub forma de drojdie uscata si 95% drojdie comprimata cu 28% substanta uscata.
Drojdiile ocupa un loc unic in lunga istorie a omenirii. Nici un alt grup de microorganisme nu a fost mai intim asociat cu progresul si bunastarea omenirii ca drojdiile.
Din timpuri stravechi au fost folosite microorganisme in dese randuri inconstient, pentru fermentarea aluatului la fabricarea painii. Maiaua acida reprezinta primul preparat de drojdie care a fost folosit ca mijloc de afanare a aluatului si isi are originea la egipteni (anul 6000 i.d.H.). Mult timp prepararea drojdiei de panificatie a ramas la acest stadiu de tehnica. Din Egipt tehnologiile de fabricare a painii au fost preluate in Grecia si de aici in Roma Antica si Imperiul Roman.
Un progres considerabil l-a adus folosirea drojdiilor de bere si vin si a drojdiilor de la fabricarea alcoolului in scopuri de panificatie, drojdii care inca din secolul al XVIII-lea rezultau ca produse reziduale ale acestor mici industrii. Prin introducerea in fabricile de bere a drojdiei de fermentatie inferioara, acestea nu au mai putut fi folosite in panificatie atat datorita unei temperaturi optime mai scazute cat si continutului mai ridicat in enzime proteolitice care ataca glutenul, ceea ce face ca painea sa nu creasca. Astfel fabricile de alcool au ramas singura sursa de drojdie de panificatie, cautandu-se sa se mareasca randamentul in drojdie in dauna celui in alcool, cantitati insa insuficiente pentru satisfacerea cererii crescande. De aceea, s-au cautat cai pentru a mari cantitatile de drojdie de alcool si au aparut procedee mai distincte pentru fabricarea drojdiei in scopuri de panificatie.
Procedeul cel mai vechi, care a avut drept scop fabricarea drojdiei presate, este asa numitul procedeu olandez, utilizat pentru prima data la Schiedam (Olanda) in jurul anului 1800. In anul 1810 procedeul a fost introdus si in Germania.
Primul pas important in dezvoltarea tehnologiei drojdiei de panificatie a fost procedeul vienez, aparut in 1860. Acesta se baza pe fermentatia unor plamezi vascoase obtinute din malt, porumb si secara. Pentru protejarea de contaminari, inainte de fermentarea propriu-zisa a drojdiei, plamada era supusa unei fermentatii lactice timp de 24÷56 ore la temperaturi intre 50 si 600C. Fermentatia in timpul careia se formau aproximativ 15 kg de drojdie din 100 kg materie prima (pe langa 30÷32 l alcool), dura 10 ore la 24÷340C. Separarea drojdiei din plamezi se realiza prin indepartarea spumei care era apoi spalata de mai multe ori si apoi presata in filtre-presa.
Introducerea presei cu parghie de catre Tebhenhaff in anul 1820, a fost un pas inainte pentru formarea drojdiei presate, pentru forma sa de vanzare. In anul 1867 ea a fost inlocuita prin filtrul presa a lui Dohne care se foloseste si in prezent. Prima masina continua de fasonat si portionat drojdia a fost construita de Simmen in 187
Incepand din anul 1880, cand Pasteur a descoperit rolul oxigenului in multiplicarea drojdiei (efectul Pasteur) s-a introdus aerarea plamezilor, obtinandu-se randamente superioare in drojdie, pana la 50÷60%. La imbunatatirea acestora a contribuit si constatarea ca multiplicarea drojdiei are loc mai rapid in plamezi mai diluate (circa 40Bllg) si atunci cand se foloseste o cantitate mai mare de cuib de drojdie.
Howman a folosit in Anglia in anul 1896 un sistem de aerare, la scara industriala, in linul de fermentare la producerea drojdiei. Cerealele erau baza materiei prime, iar randamentul a crescut de la 10÷13 kg la 20 kg drojdie presata din 100 kg materie prima cu o scadere a productiei de alcool de la 28÷30% la 20÷21%. La sfarsitul secolului materia prima pentru producerea drojdiei se compunea din circa 50÷55% porumb, 25÷30% malt verde si 10÷15% germeni de malt. Inca din anul 1895 s-au depus eforturi, in primul rand in Austria, ca sa se inlocuiasca hidratii de carbon scumpi din cereale, prin zaharul mai ieftin, din melasa. Prin dilutii avansate ale melasei, corectarea pH-ului, adaosuri de substante nutritive, cantitati mari de cuib de drojdie, cat si printr-o aerare intensa a mediului s-a ajuns la randamente ridicate de drojdie comprimata de circa 10% fata de melasa, fara ca in mediu sa se mai formeze alcool etilic. Aceste procedeu cunoscut sub denumirea de procedeul prin aerare si alimentare continua este utilizat si astazi la fabricarea drojdiei de panificatie.
Prin folosirea exclusiva a melasei din anul 1925 la fabricarea drojdiei de panificatie au fost ridicate probleme pentru alimentarea cu azot a drojdiei, careia pana atunci nu i se acordase nici o atentie sau numai foarte putin. Procedeul lui Wohl si Scherdel a reprezentat un important progres, deoarece a propus ca la cultivarea drojdiei in plamezi din melasa, 10÷50% din azotul organic sa fie inlocuit prin azot anorganic. Prin aceasta se oferea si posibilitatea sa se faca mai usor bilantul adaosurilor de azot.
Un alt pas important in dezvoltarea tehnologiei drojdiei de panificatie l-a reprezentat introducerea separatoarelor centrifugale pentru separarea economica a drojdiei din mediul sau de cultivare. Mult timp plamada framantata se pompa in asa numitele cazane de limpezire, unde drojdia trebuia sa se depuna si de unde era separata prin decantare de plamada alcoolica sau de apa de spalare, pentru ca de aici sa fie dusa la filtrele presa. La inceput s-au folosit separatoare cu functionare periodica (1892, Copenhaga), inlocuit apoi cu separatoare centrifugale cu functionare continua, produse de firmele Westfalia si Alfa-Laval. De asemeni, si pentru limpezirea melasei, care pana atunci se realiza prin decantare, au fost realizate separatoare pentru limpezire, cu evacuarea continua sau periodica si automata a namolului separat.
Necesitatile pentru marirea stabilitatii in timp a drojdiei au condus la dezvoltarea gradata a drojdiei uscare active. O drojdie uscata de calitate inferioara era cunoscuta inainte de 1900 si drojdia de panificatie uscata numita Florylin se gasea in comert in Germania dupa 1918, dar eforturile concentrate pentru producerea pe scara larga a unui produs acceptabil n-au fost facute decat dupa 1920.
O drojdie uscata buna a fost fabricata in Australia la inceputul anilor 1940, dar cantitati mari au fost fabricate in America de Nord in timpul celui de-al doilea razboi mondial. Mai apoi s-au facut in mod treptat imbunatatiri, dar drojdia uscata n-a inlocuit in totalitate folosirea drojdiei comprimate (umede).
In afara de utilizarea in panificatie, drojdiile sunt folosite pentru producerea pe scara industriala de proteine, aminoacizi, vitamine enzime, introduse in prezent in hrana animalelor,
In multe tari ale lumii drojdiile de panificatie se considera cele mai economice si utile materii prime pentru producerea extractelor proteice cu concentratie mare de proteine. In ultimii ani, s-a observat tendinta sporirii fabricarii drojdiei de panificatie pentru obtinerea de proteine alimentare, deoarece indicatorii sai organoleptici sunt apropiati de indicatorii proteinelor extractelor de carne. Dupa calculele Institutului Alimentar AMH al Rusiei, cerinta unui singur om in drojdie o constituie 2 kg pe an. Conform datelor pe anul 1985 cerintele de drojdie pentru un singur om au fost: in Finlanda - 2,5 kg, Anglia - 1,8 kg, S.U.A. - 1,4 kg, fosta U.R.S.S - 1,49 kg.
In prezent, datorita cunostintelor in domeniul geneticii s-au pus bazele ameliorarii drojdiilor industriale si modalitatilor practice de imbunatatire a calitatilor lor prin inducerea, identificarea, izolarea si caracterizarea unor mutante si linii cu proprietati biologice si economice superioare, obtinerea unor hibrizi si recombinatii prin tehnici de inginerie genetica si fuziune de protoplasti au deschis largi perspective pentru ameliorarea drojdiilor industriale, fuziunea de protoplasti facilitand obtinerea unor hibrizi intraspecifici, interspecifici si intergenetici prin depasirea barierelor sexuale si a eliminarii incompatibilitatii genetice.
In ultimii ani, in industria drojdiei de panificatie, s-au realizat progrese semnificative care au contribuit la imbunatatirea randamentelor de fabricatie, a calitatii drojdiei si implicit la imbunatatirea eficientei economice a productiei de drojdie.
Preocuparile cercetarilor din acest domeniu important al industriei alimentare s-au indreptat, in special, in urmatoarele directii principale:
selectionarea unor tulpini de drojdie cu insusiri calitative superioare;
imbunatatirea mediului de cultivare a drojdiilor;
perfectionarea tehnologiilor de fabricatie;
perfectionarea functionarii utilajelor tehnologice.
In industria panificatiei calitatea drojdiei este dependenta de viteza cu care aceasta se adapteaza la conditiile din aluat si in special pentru a produce maltaza. Printre glucidele existente in aluat sau formate in urma hidrolizei amidonului sub actiunea α si β-amilazei prezente in faina (sau din surse exogene), exista o diferenta in secventa de absorbtie si fermentare. Astfel cel mai rapid sunt absorbite hexozele (glucoza), apoi zaharoza si maltoza. Celula de drojdie absoarbe usor hexozele care trec prin membrana citoplasmatica celulara prin difuzie simpla in cazul in care exista un gradient de concentratie intre concentratia lor in exteriorul celulei si prin translocatie de grup a esterilor fosforici ai hexozelor prin intermediul hexokinazelor.
Zaharoza este hidrolizata in exteriorul membranei citoplasmatice, in regiunea peretelui celular unde este localizata invertaza si este absorbita cu o viteza echivalenta cu cea a hexozelor din care este formata, respectiv glucoza si fructoza, glucide direct fermentescibile. Fermentarea succesiva se explica prin adaptarea treptata a celulei, prin inducerea enzimelor adaptive, pe masura ce in mediu se epuizeaza hexozele.
Maltoza, principalul diglucid prezent in aluat este fermentat numai dupa o perioada de inductie necesara pentru formarea enzimei maltaza (α-glucozidaza). Maltoza si maltotrioza patrund in celula de drojdie sub influenta unor permeaze specifice induse in prezenta lor, enzime care se comporta ca sisteme active de transport. Dupa patrunderea in interiorul celulei, maltoza sub actiunea α-glucozidazei induse este hidrolizata in glucoza (2 moli) si are loc fermentarea rapida.
Harris specifica existenta a cinci gene distincte, responsabile pentru formarea α-glucozidazei si fermentarea maltozei de catre Saccharomyces cerevisiae. Metabolizarea maltozei este sub controlul a 5 gene MAL complex incat pentru a se produce fermentarea maltozei actioneaza o gena reglatoare pentru maltaza si pentru maltozopermeaza. Ambele enzime sunt induse de catre maltoza si sunt represate metabolic de catre glucoza. Astfel, adaosul de glucoza poate conduce la initierea urmatoarelor efecte: o scadere a activitatii enzimatice, inactivarea maltazei sau represia catabolica a sintezei enzimatice.
Controlul fermentarii maltozei este realizat prin doua mecanisme diferite si anume prin transcriptie, cand are loc transmiterea informatiei genetice de catre gena structurala prin intermediul acidului ribonucleic mesager care apoi efectueaza translatia. Ca rezultat se produce reglarea genetica a biosintezei enzimelor adaptive, respectiv a maltozopermeazei si a maltazei(α-glucozidazei).
O drojdie buna de panificatie trebuie sa aiba un timp cat mai scurt de inducere pentru sinteza maltazei si maltozopermeazei. Capacitatea maltazica isi pierde din importanta pentru aluaturile preparate cu adaos de zaharoza sau siropuri de amidon. In timpul depozitarii drojdiei de panificatie, capacitatea ei de a fermenta maltoza descreste mult mai mult decat cea de a fermenta glucoza. Sarea inhiba fermentarea maltozei in mai mare masura decat a altor glucide, precum si multiplicarea drojdiilor din specia Saccharomyces cerevisiae (Dan, V., 1999).
In tara noastra, tehnologia de obtinere a drojdiei de panificatie prevede ca materie prima melasa, bogata in diglucidul zaharoza. Prin utilizarea in panificatie drojdia trece de la un mediu de cultivare bogat in zaharoza, la un mediu specific aluatului bogat in maltoza. De aceea este foarte important de a obtine drojdii cu activitate maltazica superioara, prin reducerea perioadei de inducere si stimularea vitezei de fermentare a maltozei care se formeaza in aluat sub actiunea enzimelor amilolitice ale fainii.
Industrial, se urmareste:
obtinerea de drojdii de panificatie, cu activitate enzimatica superioara, si in special cu potential inalt de fermentare a maltozei, prin imbunatatirea conditiilor de cultivare si de pastrare;
asigurarea unui metabolism activ intracelular, posibil prin biosinteza de enzime constitutive si adaptive, in procesul cultivarii si in mentinerea in stare activa a enzimelor complexului zimazic prin prevenirea proceselor de degradare ireversibila si inactivarea lor.
1. PREGATIREA MELASEI IN VEDEREA MULTIPLICARII DROJDIEI
In vederea transformarii melasei intr-un mediu favorabil pentru multiplicarea drojdiei sunt necesare aceleasi operatii pregatitoare de la fabricarea alcoolului din melasa si anume:
diluarea melasei;
acidularea;
limpezirea si sterilizarea.
1.1. Diluarea melasei
Melasa introdusa in fabricatie este mai intai cantarita in vederea stabilirii consumurilor specifice si a randamentelor in drojdie, dupa care se efectueaza operatia de diluare.
Diluarea melasei la fabricarea drojdiei se realizeaza in doua etape:
diluarea initiala pana la 600Bllg in scopul cresterii fluiditatii, care sa permita curgerea libera a melasei prin conducte si sa favorizeze sedimentarea impuritatilor mecanice aflate in suspensie in cursul operatiei de limpezire;
diluarea finala pana la concentratia corespunzatoare fazei respective de multiplicare a drojdiei.
1.2. Acidularea melasei
Dupa diluarea melasei se face acidularea, de regula cu acid sulfuric pana la un pH final de 4,5÷5. Acidul sulfuric adaugat contribuie la limpezirea melasei si in acelasi timp pune in libertate acizii organici din sarurile lor. Prin aciditatea pe care o creeaza in plamezi acidul sulfuric protejeaza drojdia in cursul multiplicarii fata de contaminarile cu microorganisme straine, astfel incat nu este necesar sa se lucreze in conditii absolut pure.
Acidularea plamezilor se face diferentiat in functie de faza de multiplicare a drojdiei. Astfel, in primele trei faze de multiplicare a drojdiei, aciditatea este mult mai ridicata decat in ultimele doua faze, pentru a se evita aparitia contaminarilor.
Pentru corectarea pH-ului plamezilor de melasa din diferite faze de multiplicare se pot folosi si alti acizi, cum ar fi acidul fosforic, acidul lactic, etc.
1.3. Limpezirea si sterilizarea melasei
Operatia de limpezire a melasei este absolut necesara pentru indepartarea suspensiilor si substantelor coloidale care sunt daunatoare pentru dezvoltarea drojdiei si conduc la inchiderea culorii drojdiei - produs finit.
Pentru limpezirea melasei se folosesc in practica mai multe procedee: limpezirea prin sedimentare; limpezirea prin centrifugare; limpezirea prin filtrare.
Limpezirea melasei prin sedimentare se efectueaza prin incalzire pana la fierbere si pastrarea melasei astfel tratate in vederea decantarii naturale.
Procedeul de limpezire prin sedimentare reprezinta procedeul clasic de limpezire a melasei, care se realizeaza la cald sau la rece in vase de limpezire prin adaos de acid sulfuric si barbotare de aer comprimat. Este unul din procedeele cele mai eficiente de limpezire a melasei deoarece sub actiunea ionilor de H+ ai acidului sulfuric are loc coagularea substantelor coloidale incarcate cu sarcina negativa si astfel indepartarea lor din melasa. In afara de aceasta acidul sulfuric elibereaza din sarurile lor acizi volatili daunatori pentru multiplicarea drojdiei, care pot fi apoi indepartati prin fierbere si aerare. Sub actiunea acidului sulfuric are loc si descompunerea nitritilor toxici pentru drojdie la dioxid de azot care se indeparteaza apoi prin fierberea si aerarea melasei. De asemenea, are loc in mediu acid si invertirea unei parti din zaharoza, formandu-se glucoza si fructoza direct asimilabile, care accelereaza multiplicarea drojdiei.
In cazul in care melasa este de buna calitate se poate aplica si procedeul de limpezire la rece cu acid sulfuric, care dureaza 8÷10 ore.
Procedeul de limpezire prin sedimentare prezinta dezavantajul unei productivitati mai scazute si a unor spatii de dimensiuni mari pentru limpezire, deoarece pentru fiecare lin de multiplicare a drojdiei din fazele III, IV si V este necesar un vas de limpezire cu capacitatea de 10÷15m3.
In prezent, se prefera aplicarea de tehnici de curatire si sterilizare continua a melasei intr-un proces complet automatizat. Pentru acest scop se folosesc separatoare centrifugale si schimbatoare cu placi, realizandu-se o purificare a melasei de pana la 95%.
Cele mai cunoscute metode si instalatii de curatire si sterilizare continua poarta denumirea Westfalia si Alfa-Laval, dupa denumirea firmelor producatoare.
In instalatia Westfalia melasa este acidulata slab in prealabil cu acid sulfuric, diluata cu apa fierbinte si preincalzita la 550C. Intr-un schimbator de caldura cu placi melasa este incalzita si mentinuta la temperatura constanta de 1400C cu ajutorul unui circuit de reglare. Melasa trece prin zona de mentinere a temperaturii timp de 6 secunde ajungand apoi intr-un recipient de detentie unde este racita la 150C. Apoi este trecuta in separatorul centrifugal. Eliminarea namolului din separatorul centrifugal se efectueaza automat printr-un dispozitiv de comanda si ventile magnetice. In timpul eliminarii namolului se intrerupe alimentarea cu melasa in separator.
Instalatia "Alvotherm" a firmei Alfa-Laval urmareste asigurarea sterilizarii melasei la 1200C prin incalzire indirecta cu abur, mentinere la aceasta temperatura timp de 10 secunde si recuperarea in mare parte a energiei termice consumate.
In unele tari, pentru limpezirea melasei se utilizeaza filtrele Schenck, filtre cu kieselgur, obtinandu-se randamente ridicate in biomasa si un produs de culoare mai deschisa.
1.4. Adaosul de substante nutritive
Drojdiile au nevoie pentru crestere, multiplicare si mentinerea activitatilor biologice de prezenta in mediul de cultivare de substante nutritive care sa contina pe de o parte, elemente chimice necesare pentru sinteza constituentilor celulari, pentru activitatea enzimelor si sistemelor de transport si, pe de alta parte, sa le furnizeze substantele necesare pentru producerea de energie biologic utila.
Zarnea si colab.(1980) considera ca un mediu de cultura poate fi definit ca un suport nutritiv sterilizat, care permite dezvoltarea si studiul unui microorganism in afara nisei ecologice naturale. Fiechter (1981,1984) arata ca proiectarea sistematica a mediilor de cultura poate fi facuta in sase etape:
selectia componentelor si a formei in care acestea sunt prezentate in mediu;
prepararea mediului;
diagrama preliminara (concentratia biomasei si a substratului functie de dilutie);
determinarea constantelor de saturare si inhibitie pentru sursa carbon;
optimizarea mediului;
utilizarea tehnicii chemostatului in cazul folosirii mediului cu compozitia optima.
Pentru estimarea microelementelor necesare dezvoltarii drojdiilor pot fi utilizate datele prezentate in literatura referitoare la necesitatile nutritionale ale microorganismelor. Desi in lucrarile de specialitate sunt prezentate si retete de mediu, utilizarea acestor retete trebuie facuta cu multa prudenta, iar retetele de medii industriale reprezinta secrete de fabricatie.
Un aspect foarte important care trebuie avut in vedere pe parcursul procedurii de proiectare si optimizare a mediilor de cultura il constituie cerintele tehnico-economice, deoarece pretul materiilor prime reprezinta 10÷60% din costul de productie.
Intrucat pot folosi numai energie eliberata prin reactii chimice oxidative, drojdiile apartin tipului de nutritie chimiotrof, heterotrof, in sensul ca nu-si pot sintetiza substantele proprii decat pornind de la substante organice pe care sa le descompuna pana la produsi simpli utilizabili in metabolismul lor. Din acesti compusi ele isi realizeaza scheletele carbonice necesare sintezei constituentilor celulari si energia necesara pentru a permite initierea reactiilor de biosinteza (Dan, V., 1999).
Nutritia hidrocarbonata. Carbonul reprezinta circa 50% din totalul compusilor organici, de aceea necesarul in compusi de natura hidrocarbonata este foarte mare.
Principala sursa de energie si de carbon pentru drojdie este reprezentata de glucide. Natura si concentratia optima difera de tipul de drojdie si de caracteristicile procesului tehnologic. Drojdiile se dezvolta pe medii ce contin hexoze (D-glucoza, D-manoza si D-fructoza). D-galactoza nu este fermentata decat in cazul unor adaptari speciale a drojdiilor. Pentozele nu sunt metabolizate de catre drojdia de panificatie.
Lactoza nu este consumata de drojdiile de panificatie. Melibioza este asimilata de catre drojdiile din specia Saccharomyces uvarum, dar nu si de cele din specia Saccharomyces cerevisiae, fiind astfel un test pentru diferentierea celor doua specii.
Cu privire la concentratiile optime de hexoze, in conditiile din industrie, in functie de scopul de utilizare si de tehnologia aplicata s-au stabilit limite bine definite. Astfel, pentru drojdia de panificatie se folosesc medii cu concentratii de glucide fermentescibile de circa 2%, in cazul utilizarii tehnologiei clasice si de pana la 10% in aplicarea la procese cu aerare intensiva. La scaderea concentratiei apare pericolul marit de contaminare, respectiv de asimilare a glucidelor de catre alte microorganisme prezente in mediu. Concentratii ridicate de glucide in mediu impiedica inmultirea drojdiilor din genul Saccharomyces. Astfel, la concentratii de peste 20% apar fenomene de plasmoliza din cauza presiunii osmotice prea ridicate in mediu.
Asimilarea glucidelor depinde, in afara de concentratie, de temperatura, pH, cantitatea de celule prezente in mediu cat si de alti factori.
La inceputul fiecarei faze de crestere, in productia industriala a drojdiei de panificatie, trehaloza (drojdiile de panificatie contin pana la 14% trehaloza) este repede si aproape complet metabolizata, dar este resintetizata in ultima parte a fazei de crestere (Suomalainen si Pfaffi, 1961). Aceasta mobilizare rapida a rezervei de carbohidrati poate duce la acumularea unei rezerve de energie ce va fi folosita in faza de lag, cand celulele se pregatesc de diviziune. Trehaloza este hidrolizata specific de trehalaza care este o α-glucozidaza.
Drojdiile din specia Saccharomyces cerevisiae pot asimila etanolul si produsii de oxidare ai etanolului, acetaldehida si acidul acetic, precum si glicerina si acidul lactic.
Fiind facultativ anaerobe, drojdiile de panificatie sunt inzestrate cu mecanisme de schimb anaerob si aerob. Directia schimbului hidratilor de carbon in celulele de drojdie depinde nu numai de prezenta sau absenta aerarii, dar si de concentratia si tipul surselor de hidrati de carbon. In conditii aerobe, la cultivarea pe medii care contin glucoza (precum si fructoza si zaharoza) in cantitate mai mare de 5 g/l, se observa o glicoliza intensa, asa numita fermentare aeroba sau efectul Crabtree. Prin fermentare aeroba se oprima enzimele ciclului acizilor tricarboxilici (represie catabolica) si in mediul nutritiv se acumuleaza etanol. Dupa scaderea concentratiei glucidelor in mediul nutritiv se activeaza enzimele de respiratie si celulele trec la metabolismul oxidativ in timpul caruia drojdia asimileaza alcoolul format. Incepe a doua faza logaritmica a cresterii.
Directia metabolismului influenteaza compozitia celulelor de drojdie, mai ales continutul seriei de metaboliti care participa in reactiile importante ale celulei. In tabelul de mai jos sunt prezentate date despre continutul aminoacizilor liberi la Saccharomyces cerevisiae, cultivata pe medii cu diferite concentratii de glucoza si in medii cu galactoza. Prin marirea continutului de glucide in mediul nutritiv, cantitatea de aminoacizi liberi in celula scade, totusi mult mai brusc modificarea se observa in mediile cu glucoza. In acelasi timp se activeaza enzimele care participa la schimbul anaerob al hidratilor de carbon, piruvatdecarboxilaza si alcooldehidrogenaza.
Tabelul 20
Dependenta continutului in aminoacizi si activitatea unor enzime
la drojdia Saccharomyces cerevisiae de tipul si concentratia sursei de carbon
|
Sursa de carbon |
Glucoza |
Galactoza |
||||||
|
Concentratia, g/l | ||||||||
|
Aminoacizi, mg/g s.u. | ||||||||
|
Piruvatdecarboxilaza, nmoli/mg prot./min | ||||||||
|
Alcooldehidrogenaza, nmoli/mg prot./min. | ||||||||
La cresterea drojdiei pe galactoza, in acelasi timp au loc procese de respiratie si fermentatie, totusi prin marirea continutului galactozei in mediul nutritiv se intensifica respiratia si se oprima intr-o oarecare masura fermentarea. Tipul sursei de carbon exercita o influenta asupra concentratiei si continutului in diferiti metaboliti (tab. 20).
Asa cum se observa din tabelul 21, este evidenta deosebirea substantiala a drojdiilor cultivate pe mediul cu etanol: un continut mai ridicat in proteine, decat la drojdiile cultivate pe medii nutritive cu melasa, mai multi aminoacizi liberi.
Tabelul 21
Continutul in metaboliti a celulelor de drojdii cultivate pe medii cu diferite surse de carbon [mg/g s.u.]
|
METABOLIT |
SURSA DE CARBON |
||
|
Glucoza |
Galactoza |
Etanol |
|
|
Hexozo-6-fosfat | |||
|
Fructozo-1,6-difosfat | |||
|
Adenozinmonofosfat (AMF) | |||
|
Adenozinmonofosfat ciclic (c-AMF) | |||
|
Citrat | |||
|
Cetoglutarat | |||
S-au descoperit, de asemenea, cateva deosebiri in fractiile peptidice: la drojdiile cultivate pe medii cu etanol, fractia peptidica contine mai multa cistina si mai putini acizi diaminocarboxilici, decat la drojdiile cultivate pe medii cu glucoza. Drojdiile cultivate pe etanol sunt mai bogate in ergosterol, decat drojdiile cultivate in mediu glucidic, ceea ce explica metabolismul aerob la cresterea drojdiei pe etanol (Gancedo, J.M., 1973).
Proteinele drojdiilor se pot separa in cateva fractii. Continutul total in azot, proteine si proteine separate in fractii la cultivarea drojdiilor pe medii cu compozitie diferita sunt prezentate in tabelul 22. Primele doua fractii reprezinta proteine de tipul albuminelor, globulinelor si partial nucleoproteide, a 3-a si a 4-a fractie proteine greu solubile.
Tabelul 22
Continutul in azot total, proteine si fractii proteice la drojdia Saccharomyces cerevisiae cultivata pe diferite medii nutritive
|
Nr. var. |
Sursa de C |
Conc. |
(NH4)2SO4 [g/l] |
N2 total [%/s.u.] |
Proteine[%/s.u.] |
Fractii proteice |
||
|
Sol. tampon |
NaCl |
NaOH |
||||||
|
Melasa | ||||||||
|
Melasa | ||||||||
|
Melasa | ||||||||
|
Etanol | ||||||||
*in % zahar
La drojdia Saccharomyces cerevisiae predomina fractiile proteice 1 si 2. Tipul si concentratia sursei de carbon in mediul nutritiv exercita o influenta asupra spectrului proteic de fractii ale drojdiei.
Modificarea compozitiei mediului nutritiv sau intensificarea aerarii influenteaza directia si viteza proceselor biochimice principale ale celulei si determina nu numai activitatea generala a enzimelor, dar si prezenta si activitatea enzimelor separate. Tipul si concentratia sursei de carbon in mediul nutritiv determina activitatea enzimelor glicolizei, ciclului acizilor tricarboxilici si enzimelor care participa la glicogeneza. De exemplu, glucoza este represor catabolic al activitatii enzimelor ciclului glioxalic: malatsintetaza, izocitratliaza, malatdehidrogenaza. O sensibilitate deosebita, la prezenta glucozei in mediul nutritiv o prezinta fructozo-1,6-difosfataza, provenita din celulele de drojdie numai dupa indepartarea din mediul nutritiv a glucozei. La cresterea drojdiilor pe medii nefermentative (etanol, lactat), adaosul in mediul nutritiv de glucoza provoaca inactivarea rapida si totala a fructozo-1,6-difosfatazei.
Dintre enzimele care participa la procesele respiratorii ale celulei de drojdie, cea mai sensibila la represiile catabolice este citocrom-c-oxidaza (la concentratii ale glucozei de 0,1%). Pe masura consumului de glucoza activitatea enzimelor ciclului glioxalic se restabileste.
Nutritia azotata are un rol important in metabolismul drojdiilor, azotul fiind elementul major din compozitia proteinelor, enzimelor.
Drojdiile prezinta o anumita particularitate in nutritia azotata pentru ca ele produc enzime proteolitice intracelulare, enzime cu molecule mari, endogene, incat drojdiile nu pot folosi protidele in nutritia azotata. Aceste enzime proteolitice devin active cand celula de drojdie este lipsita de mediu nutritiv, actionand asupra compusilor celulari producand autoliza celulei de drojdie si in final moartea celulei (Dan, V., 1975).
Pentru Saccharomyces cerevisiae, sarurile anorganice de amoniu servesc ca sursa buna de azot, asigurand cresterea normala a celulei si biosinteza tuturor compusilor azotati. Majoritatea aminoacizilor naturali, cu exceptia acidului aspartic, asparaginei, acidului glutamic si glutaminei sunt asimilati de Saccharomyces cerevisiae mult mai lent decat ionii de amoniu, cu toate ca celulele de drojdie contin permeaze pentru aminoacizi. Transportul aminoacizilor are loc la fel ca si a glucidelor, prin absorbtie si difuzie pana la un nivel care nu depaseste concentratia lor in mediu, peste acest nivel transportul se realizeaza cu ajutorul unor transportori specializati. Sunt date in literatura care arata ca, azotul din acidul aspartic si asparagina este asimilat de doua ori mai repede decat azotul amoniacal, iar acidul glutamic este asimilat dupa ionul de amoniu (Anghel, I. et al., 1989).
Aminoacizii in celula de drojdie sunt supusi dezaminarii si transaminarii, totusi nu este exclusa posibilitatea, ca unii dintre ei sa fie utilizati direct in sinteza de proteine.
Prin cultivarea drojdiei de panificatie pe medii fara biotina (sursa de carbon-glucoza), prin adaosul in mediul nutritiv de acid aspartic se mareste considerabil cresterea drojdiilor. Efectul stimulator maxim s-a observat la adaosul concomitent de acid aspartic si acizi grasi nesaturati. Adaosul acidului aspartic poate compensa partial deficitul de biotina.
La cultivarea drojdiei Saccharomyces cerevisiae pe medii cu etanol ca sursa de carbon, biomasa rezultata se mareste daca in mediul nutritiv se adauga o cantitate de 0,075% acid glutamic. S-a dovedit ca acidul glutamic favorizeaza utilizarea etanolului in biosinteza. O influenta stimulatoare a cresterii, alaturi de acidul glutamic, o exercita si glutationul. Influenta favorabila a aminoacizilor indicati se manifesta numai in cazurile in care ei se adauga in mediul nutritiv la inceputul procesului de cultivare (Beker, M.E., 1977).
Saccharomyces cerevisiae nu poate asimila β-aminoacizii, ca de exemplu β-alanina si acidul β-aminobutiric. β-alanina, desi neutilizabila ca sursa de azot, este un factor de crestere, iar atunci cand este adaugata in cantitati mici poate mari cresterea drojdiei in prezenta unei surse asimilabile de azot.
Prezenta unor aminoacizi in cantitati excesive exercita actiuni toxice asupra drojdiilor. Astfel, Middelhoven (1977) citeaza cazul cisteinei la concentratii mai mari de 25 mg·dm-3. Lewis si Phaff au studiat fenomenele de eliminare a substantelor azotoase din drojdie in mediu, denumind fenomenul "excretie soc". Prin realizarea unei suspensii de drojdie intr-o solutie de glucoza se elimina rapid aminoacizii care se absorb apoi lent in doua sau trei ore. Cantitatea depinde de tulpina de drojdie, de continutul de aminoacizi intracelulari, de temperatura. S-a ajuns la concluzia ca acest soc este rezultatul schimbului de substante in celula la fluxul continuu de glucide fermentescibile prin membrana celulara.
Comparand influenta diferitelor saruri de amoniu asupra cresterii drojdiilor, Pirshle(1930) a gasit ca fosfatul de amoniu bibazic utilizat ca sursa de azot determina o crestere eficienta a drojdiei de panificatie. Surse la fel de bune includ fosfatul de amoniu mono-si tribazic, sulfatul de amoniu, bicarbonatul, acetatul, lactatul si tartratul, in timp ce clorura de amoniu s-a dovedit inferioara ca sursa de azot (Anghel, I. et al., 1989).
Azotul amoniacal liber in functie de concentratie a fost gasit ca are un efect limitant in rata de fermentare a drojdiilor si rata de crestere. Pe de alta parte, ionul de amoniu influenteaza rata de producere a alcoolului, influenta lui fiind conditionata de raportul lui cu ionul fosfat (Marchetti, R. et al., 1991).
Drojdiile nu pot asimila nitratii (care pot sa aiba un efect de inhibare al multiplicarii), iar nitritii au efect toxic si opresc dezvoltarea drojdiilor. La un continut in mediu de 0,0005% nitrit este inhibata inmugurirea normala a drojdiilor. Continutul in nitriti de 0,004% inhiba inmultirea drojdiilor de cultura cu 50%, iar la cantitatea de 0,02% se reduce inmugurirea. Nitritii modifica morfologia celulelor, intarzie respiratia, inhiba multiplicarea si activitatea fermentativa. Cea mai mare sensibilitate o prezinta in faza lag de crestere. S-au efectuat cercetari care au aratat ca, daca concentratia nitritilor in mediu se micsoreaza in cursul multiplicarii drojdiilor de la 0,004% la 0,002%, randamentul in drojdie se mareste cu 8÷10%, iar la concentratii de 0,001% se mareste cu 17÷21%. Kauzman arata ca o concentratie de 0,01% NO2 a micsorat randamentul si puterea de fermentare a drojdiilor.
Nutritia minerala. Este un proces fiziologic prin care microorganismele preiau din mediu substante minerale care intra in constitutia compusilor celulari. Diferitele elemente necesare nutritiei minerale intra in structura metalenzimelor, a pigmentilor, a unor vitamine si sunt necesare in cantitati foarte mici. La doze prea mari substantele minerale pot produce efecte toxice (Dan, V., 1999).
Un tablou general asupra efectului concentratiei optime de ioni asupra echipamentului enzimatic si rolul structural pentru drojdii este pe larg prezentat de Soumalainen si Oura (1970), James (1984, 1986). Performantele drojdiei sunt dramatic influentate de modificarile concentratiei ionilor limita, uneori in intervale foarte inguste. Concentratia optima pentru fiecare tip de ioni nu poate fi individual definita. De fapt, variatia unor ioni selectati pot sa produca efecte de stimulare sau inhibare in functie de concentratia altor ioni sau nutrienti.
Incercarea de optimizare a mediilor de fermentare, prin manipularea compozitiei lor ionice, necesita cunostinte mai profunde despre interactiunile fizico-chimice, in care sunt implicati atat ionii cat si solutiile lor si functionalitatea membranei celulare.
Fosforul este un element necesar atat pentru cresterea drojdiilor cat si pentru fermentatie, el reprezinta, sub forma de oxizi, aproape 50% din cenusa drojdiei.
Dintre toate substantele minerale, fosfatii au importanta cea mai mare (Markham, 1967). Acestia sunt consumati pentru dezvoltare, iar in caz de exces se stocheaza in celule, putand fi reutilizati in cazul lipsei de fosfor in mediu, drojdia dezvoltandu-se in continuare. Fosforul participa la transmiterea energiei in celulele de drojdie prin intermediul ATP si ADP, rolul acestuia fiind evidentiat la metabolismul glucidic.
Sulful care intra in compozitia aminoacizilor cu sulf este preluat de drojdii din sulfatul anorganic care insa poate fi inlocuit partial sau in intregime de alti compusi anorganici sau organici cu sulf.
Cele mai multe specii de Saccharomyces manifesta o crestere buna atunci cand sulfatul este inlocuit cu sulfit sau tiosulfat. Ele sunt insa incapabile de a utiliza aminoacizii cu sulf ca de exemplu, cisteina sau cistina, drept surse de sulf. Absorbtia sulfatului cere energie si atat glucoza cat si nutrientii cu azot trebuie sa fie prezenti in mediu. Intensitatea de aerare in cultivarea lui Saccharomyces cerevisiae influenteaza capacitatea sa de a folosi diferite surse de sulf. Marirea aeratiei si diminuarea nutritiei in timpul propagarii industriale a drojdiilor de panificatie duce la o diminuare a continutului total de sulf din celule.
Kotyk(1959) a dovedit ca absorbtia sulfului decurge cu aceeasi intensitate in ambele cazuri de aerobioza si anaerobioza.
Potasiul, element din grupa metalelor alcaline este necesar drojdiilor atat pentru crestere cat si pentru fermentatie. Absorbtia ionului K este inlesnita de absorbtia glucozei, cand aceasta este consumata, ionii de K devin netransportabili. Cand ionii de K sunt absenti din mediu, fosforul nu mai poate fi absorbit. Potasiul joaca un rol cheie in reglarea transportului cationilor bivalenti, in producerea de celule si rata de fermentare. S-a observat ca necesarul in potasiu al drojdiilor a crescut cu rata de crestere. Potasiul se regaseste in cantitati de 2,4÷2,8 % in drojdie (K2O in s.u.). In melasele normale, potasiu exprimat in K2O se gaseste in proportie de 2,5÷4,5%. Melasele cu un continut sub 2,5% K2O au influenta negativa asupra conservabilitatii drojdiei.
Magneziul este considerat un factor de crestere necesar pentru drojdii, el fiind un activator al transfosfatazei, enolazei si carboxilazei. Cand cresterea drojdiilor a fost limitata de catre magneziu, randamentul in celule a scazut, iar in compozitia proteinelor s-a observat o scadere a continutului in lizina si acid glutamic. De asemenea, magneziului i se atribuie un rol in reducerea activitatii de fermentare. In general, melasele sunt deficitare in magneziu, care trebuie sa fie asigurat la prepararea plamezilor prin adaugarea unor saruri de magneziu. In literatura de specialitate se mentioneaza ca optime urmatoarele cantitati de azot, fosfor si magneziu care se adauga sub forma de solutii la prelucrarea melasei (Banu, C. et al., 1978):
Azot (N2) 1,6÷1,8% fata de masa melasei;
Fosfor (P2O5) 0,6÷0,8% fata de masa melasei;
Magneziu (MgO) 0,1÷0,15% fata de masa melasei.
Calciul, desi aparent neesential pentru cresterea celulelor de drojdii, stimuleaza cresterea si fermentatia. Prezent sub forma de clorura in mediul de cultura, in concentratii de 25÷50 mg·dm-3 poate stimula inmultirea drojdiei Saccharomyces cerevisiae.
Litiul franeaza fermentatia alcoolica, dar stimuleaza multiplicarea drojdiei in doze de pana la 3mg·dm-3 mediu.
Borul stimuleaza activitatea complexului zimazic, in special a fosfatazei alcaline, dar blocheaza activitatea fosfatazei acide.
Fluorul poate fi prezent in medii cu saruri de fosfor si inevitabil este adus cu acestea in mediul de cultura. Prezenta permanenta a fluorului in mediu duce la adaptarea drojdiei la acesta. Din datele specialistilor polonezi, drojdiile sunt capabile sa asimileze fluor pana la concentratia de 10÷30 mg/kg fara o influenta vizibila asupra ritmului de crestere. NaF in cantitate de 20 mg·dm-3 in mediu franeaza activitatea vitala a drojdiei.
Aluminiul in concentratie de 0,01÷-0,1 mg·dm-3 contribuie la sinteza ARN la drojdii. Aluminiul este toxic, franand activitatea drojdiei la concentratii de 54 mg·dm-3 si are efect letal la 540 mg·dm-3 (in solutii de Al2SO3).
Fierul intra in compozitia multor metalenzime, succinat dehidrogenaza, catalaza si altele. Posedand valenta schimbatoare, participa activ la reactiile de oxidoreducere. In absenta ionilor de fier drojdia nu creste, totusi concentratiile ridicate in fier sunt toxice. Continutul admisibil al fierului in mediu este de 20 mg/dm3. Fierul intra in mediul de cultura cu melasa si alte componente in cantitate suficienta pentru biosinteza celulara.
Cuprul poseda valenta schimbatoare si in calitate de transportor de ioni, participa la reactii de oxidoreducere. Cuprul stimuleaza activitatea complexului zimazic si intr-o masura mai mica activitatea maltazei. Cuprul intra in compozitia multor oxidoreductaze, in concentratie de 0,01÷0,25 mg·dm-3 contribuie la multiplicarea drojdiei. Concentratii mai ridicate franeaza activitatea vitala a drojdiei, la concentratii de 50 mg·dm-3 produce inactivarea fiziologica a celulei.
Zincul intra in compozitia multor metalenzime. De exemplu, molecula de gliceroaldehidfosfatdehidrogenaza contine 2 atomi de zinc, moleculele de alcooldehidrogenaza si piruvatkinaza contin 4 atomi de zinc. Zincul accelereaza activitatea enzimelor complexului zimazic si a maltazei, imbunatateste puterea de crestere a drojdiei. El joaca un rol important in metabolismul celulelor si in concentratie de 0,2 mg·dm-3 accelereaza activitatea vitala a drojdiei. Continutul in zinc de 130 mg·dm-3 opreste multiplicarea (Novakovskaia, S.S., Sisatkii, I.I., 1980).
Melasa contine in general majoritatea factorilor (zaharoza, acizi organici, vitamine, saruri minerale, etc.) care asigura sinteza de catre celula de drojdie a substantelor mai sus mentionate si in consecinta desfasurarea normala a functiilor fiziologice a acesteia. Este insa deficitara in azot, fosfor, magneziu si uneori in alte elemente.
Pentru ca drojdia sa se dezvolte normal, in vederea asigurarii de randamente si calitate corespunzatoare, concomitent cu alimentarea cu melasa in timpul multiplicarii drojdiilor, se efectueaza si alimentarea cu solutie de substante nutritive, care contin azot si fosfor. Necesarul de azot si fosfor, pentru ca in final drojdia sa contina circa 1,8% azot si 0,8% fosfor, se calculeaza anticipat in raport cu rezultatele analizei melasei. Este necesar sa se calculeze de fiecare data necesarul de azot si fosfor, deoarece excesul in aceste elemente nu constituie in fond decat o risipa care mareste inutil cheltuielile de fabricatie. In cazul melaselor deficitare in magneziu, la prepararea solutiilor nutritive se adauga si saruri care contin acest element.
Substantele nutritive se adauga in plamezile de melasa sub forma de solutie limpede pasteurizata in prealabil pentru a se evita pericolul de contaminare. Instalatia de preparare a solutiilor de substante nutritive este formata din vase metalice pentru prepararea solutiilor, vase pentru depozitarea solutiilor si vase de dozare a solutiilor de substante nutritive, care sunt confectionate din material antiacid, intrucat solutiile de substante nutritive sunt corosive. Instalatiile clasice de preparare a solutiilor de substante nutritive sunt formate numai din vase de solubilizare. Ele prezinta dezavantajul antrenarii sedimentului depus in linurile de multiplicare si faptul ca, pentru fiecare lin de multiplicare este necesar un vas de solubilizare. Solutiile preparate de substante nutritive trebuie mentinute la temperaturi peste 65 C prin incalzirea lor cu ajutorul unei serpentine cu abur, pentru a se evita contaminarile cu microorganisme. La prepararea lor trebuie sa se urmareasca prin analiza de laborator sa nu contina anumite elemente toxice pentru drojdii peste limita admisa (in sulfat de amoniu, acid sulfuric si superfosfat de calciu, arsenul sa nu depaseasca 0,0001%, plumbul in sulfatul de amoniu 0,001%, iar in acidul sulfuric 0,001%.
2. Multiplicarea drojdiilor
2.1. Bazele procesului de multiplicare a drojdiei
Procesul de multiplicare si mecanismul biochimic de formare si dezvoltare a masei celulare nu sunt in intregime cunoscute in prezent, desi se cunosc contributiile celor mai multi factori ce participa in acest complex.
In conditiile experimentale, dinamica multiplicarii drojdiilor este bine cunoscuta. Procesul evolueaza intr-o serie de faze succesive.
Faza de latenta, de crestere zero sau de lag reprezinta etapa de timp cand dupa inoculare numarul celulelor ramane neschimbat, sau chiar scade, noile conditii de mediu implica latenta inductiei acelor enzime necesare pentru adaptarea la mediul nutritiv. Faza de latenta apare deci ca o perioada de adaptare la conditiile noi de cultura, in care drojdiile viabile din inocul isi acumuleaza in celula metaboliti si sistemele necesare cresterii, in cazul in care aceste componente biochimice le lipseau datorita conditiilor de mediu anterioare inocularii.
Faza de multiplicare exponentiala sau de crestere logaritmica este caracterizata prin aceea ca, dupa o scurta perioada (circa 2 ore) de accelerare a ritmului de crestere, in care multiplicarea se produce cu o viteza progresiva marita, acest ritm devine constant si caracteristic in anumite conditii de cultura, durata unei generatii fiind minima. Perioada de echilibru poate fi mentinuta numai atat cat nu intervin alterari importante, pe care cresterea le poate provoca in compozitia mediului. Numarul de celule de drojdie si cantitatea de materie vie formata creste temporar dupa o progresie geometrica cu ratia 2. Celulele aflate in faza exponentiala de multiplicare sunt cele mai potrivite pentru cercetari de genetica si fiziologie.
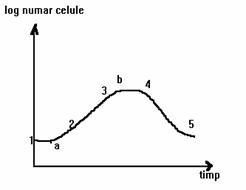
1÷2 - faza de repaos (faza lag);
1÷a - perioada de adaptare;
a÷2 - perioada de refacere;
2÷3 - faza exponentiala; 3÷4-faza stationara;
3÷b - perioada de spor de crestere negativ;
b÷4 - perioada de stationare propriu-zisa;
5 - faza finala de declin.
Fig. 20. Curba de multiplicare a drojdiilor
Faza stationara (de maturare) in care numarul celulelor viabile este maxim si ramane constant o perioada de timp. Celulele de drojdie nu mai inmuguresc, isi maresc volumul si tind spre forma sferica, se rotunjesc. Aceasta faza dureaza 1÷2 ore, in care nu se mai face alimentarea cu melasa si saruri lasandu-se drojdia sa-si consume substantele de rezerva din celula (exemplu, glicogen) si o parte din acizii organici din mediu. Intrucat functiile respiratorii ale celulei sunt acum slabite se micsoreaza debitul de aer la circa 50% din valoarea maxima folosita in faza logaritmica. Celulele de drojdii au in aceasta faza caracteristicile morfologice cele mai tipice genului si speciei.
Faza de declin se caracterizeaza printr-o scadere in progresie geometrica in raport cu timpul a numarului de celule vii. Pe masura ce mediul devine mai putin favorabil, celulele vii inceteaza a mai forma muguri, desi activitatea lor mai continua un timp dupa care mor si intra in autoliza. Acest declin poate interveni cu mai multa intarziere la tipurile "robuste" de drojdii. La sfarsitul acestei ultime faze se inregistreaza maximum absolut al numarului total de celule formate pe parcursul intregii evolutii a culturii.
Cunoasterea ratei de crestere a culturilor de drojdie este importanta in cercetare pentru studiul proprietatilor fiziologice a tulpinilor selectionate si in tehnologia de fabricatie pentru stabilirea conditiilor de reproducere cand acestea sunt folosite in calitate de cultura starter sau pentru obtinerea avantajoasa a unor compusi intracelulari.
Dupa stabilirea experimentala a curbei de crestere se pot calcula urmatorii parametri:
Cresterea exponentiala si timpul de generatie:
Numarul de diviziuni celulare:
n = ![]()
in care:
No - numar celule inocul;
N - numarul de celule rezultate prin inmugurire.
Constanta vitezei de reproducere:
V =![]() ,
,
in care:
t - timpul de cultivare
Timpul necesar pentru un ciclu de reproducere sau timpul de generatie :
g = ![]() ,
,
in care:
g - timpul de generatie, pentru dublarea numarului de celule;
n - numar de diviziuni;
v - constanta vitezei de reproducere.
Timpul de generatie pentru Saccharomyces cerevisiae la temperatura de 300C este de 2 ore (Prescott, L.M., 1990).
Dupa introducerea inoculului, cand in mediu exista o cantitate suficienta de oxigen, drojdiile pot produce oxidarea glucidelor fermentescibile pana la produsii finali ai respiratiei si are loc inmultirea celulelor. Apoi procesul de respiratie se reduce si se intensifica procesul fermentativ, iar celulele existente cresc in dimensiuni si in celule se acumuleaza glicogen.
Pentru stabilirea vitezei de inmultire se calculeaza coeficientul Km care exprima numarul de celule noi ce se formeaza pentru fiecare celula existenta, in timp de o ora cu relatia :
Km
= ![]()
in care:
N1 - numar initial dupa inoculare (la timpul t1);
N2 - numar de celule format in mediu in perioada t2-t1.
La cultivarea drojdiilor in plamezi de melasa s-a constatat in conditii practice o anomalie in desfasurarea logaritmica a curbei de dezvoltare, drojdia inmultindu-se prin impulsuri ritmice.
In cazul proceselor continue, rata de crestere a celulelor de drojdie, dupa Monod si Maxon (1955), se exprima prin relatia:
![]() = λ
= λ![]() ,
,
unde,
μ - rata de crestere specifica;
λ - rata de dilutie sau fractia volumica de lichid care paraseste vasul in unitatea de timp ;
db/dt - procentul de drojdie noua din cantitatea constanta B in vasul de multiplicare.
Cercetarile au permis stabilirea unor ecuatii matematice si a relatiilor dintre urmatorii factori care se iau in considerare la calculul multiplicarii drojdiei:
coeficientul de multiplicare;
modulul (factorul) de crestere orara;
viteza de asimilare a zaharului.
Coeficientul de multiplicare este un prim factor. Daca se noteaza cu A masa de drojdie care se afla la un moment dat in mediu, viteza de inmultire in acel moment este Ar, in care r este un factor constant al conditiilor specifice de mediu, denumit coeficient de multiplicare. Coeficientul r se exprima in grame de celule noi formate intr-o ora din gramele de celule de drojdie ce se aflau in mediu. Cantitatea de drojdie, care se afla in mediu, dupa o perioada t de multiplicare, poate fi calculata dupa urmatoarea ecuatie:
![]()
in care:
A - masa bruta a drojdiei produsa in timpul t, in grame;
A0 - masa drojdiei cuib, in grame;
e - baza logaritmica Neperiana egala cu 2,3718;
r - coeficientul de multiplicare.
Prin derivarea acestei ecuatii poate fi obtinuta viteza de inmultire care este:
![]()
Deseori, se aplica ecuatia Michaelis-Menten pentru descrierea comportarii microorganismelor in decursul dezvoltarii drojdiei. Ea se exprima prin relatia:
![]()
in care:
m - factorul de dezvoltare;
mm - factorul de dezvoltare maxima;
S - este concentratia mediului;
Ks - este constanta de saturatie, respectiv concentratia mediului la jumatatea factorului de dezvoltare.
Constantele au fost determinate experimental in medii de glucoza. Astfel Hartree (1948) a gasit pentru mm valori de 0,37 ore si pentru k, valori de 3,6*10-4 M la temperatura de 300 C si pH de circa 4.
Modulul orar de crestere (H) este un factor cu valoare practica pentru calculul cresterii orare in greutate a unei cantitati de drojdie supusa multiplicarii. Daca presupunem ca modulul orar H = 1,2 aceasta inseamna ca 1 g de drojdie creste la 1,2 g intr-o ora, la 1,2 kg la 2 ore, 1,2t in t ore. Prin urmare:
![]()
Egaland ecuatiile 1 si 2 rezulta ca:
![]()
sau:
![]()
Prin logaritmare obtinem:
![]()
sau:
![]()
Numeroase cercetari care s-au efectuat cu privire la randamentele maxime de drojdie ce se pot obtine din zaharul continut de melasa, au evidentiat ca in cele mai bune conditii drojdia foloseste pentru inmultirea ei 2/3 din carbonul moleculei de hexoza, iar restul de 1/3 pierzandu-se sub forma de dioxid de carbon. Deci, o drojdie de panificatie cu 27% substanta uscata contine 12,7% carbon. Un gram de zahar invertit contine 0,4 g carbon, din care, daca se folosesc numai 2/3 pentru formarea celulelor noi de drojdie, rezulta ca pentru 0,127 g carbon, care se gasesc intr-un gram de drojdie, sunt necesare 0,476 g hexoza.
Viteza de asimilare a zaharului in t ore de la inceperea procesului de multiplicare este data de derivata:
![]() , unde:
, unde:
![]()
![]()
Pentru obtinerea de randamente maxime in drojdie este necesar ca in plamada concentratia de zahar sa fie scazuta, aceasta realizandu-e prin adaosuri progresive de melasa cu un debit orar corelat cu necesitatile de multiplicare ale drojdiilor. Este indicat ca in orice moment din perioada alimentarii, zaharul din plamada sa fie consumat inainte de sosirea portiunii urmatoare de melasa, daca se ignora aceasta conditie, excesul de zahar se pierde sub forma de alcool.
![]() kg drojdie de
insamantare
kg drojdie de
insamantare
Daca valoarea factorului r este cunoscuta, cantitatea corecta de drojdie de insamantare se determina din ecuatia
![]() ,
,
unde G este masa zaharului asimilat in t ore. Spre exemplu, daca o plamada trebuie sa fie alimentata cu 1000 kg zahar din melasa in 12 ore si s-a determinat factorul r = 0,16, rezulta ca sunt necesare: cantitatea initiala de zahar la timpul zero trebuie sa fie de 0,476 x r x A0 = 27,5 kg/h (sau 55 kg melasa tip 50%). Cantitatea de zahar si implicit de melasa preparata creste progresiv, astfel ca in a 6-a ora este de 0,476 x r x A0 x e0,96 = 71,8 kg/h, iar in ora a 12-a, spre exemplu de 0,476 x r x A0 x e1,92 = 188 kg/h.
Un calcul mai simplu consta in stabilirea necesarului de zahar, care trebuie sa se adauge in plamada orar, in baza modulului orar de crestere H. In cazul exemplului mentionat mai sus:
![]() kg
kg
Cele 361 kg drojdie cuib vor spori la 361 x H in prima ora, deci in prima ora se vor forma 632 kg drojdie. Aceasta necesita pentru multiplicare 0,476 x 632 = 301 kg zahar.
2.2. Drojdia de cultura Saccharomyces cerevisiae
Se urmareste obtinerea in laborator a unor culturi de celule cat mai omogene, in ceea ce priveste metabolismul, randamentul, viteza de inmultire, capacitatea de reproducere si calitatea produsului finit. Inmultirea culturilor se efectueaza treptat, primele faze realizandu-se in laborator si in continuare, in statia de culturi pure a producatorului de drojdie de panificatie.
Mentinerea puritatii se realizeaza prin izolarea de celule individuale din culturi ce s-au comportat bine, din probe preluate din ultima faza de multiplicare. Pentru izolarea de celule se practica, in functie de mediul nutritiv, metode cu substrat lichid (Lindner si Hansen) si metode cu substrat solid (Koch si Hansen).
Dupa izolare se trece la verificarea puritatii culturilor izolate, vizual cu ajutorul microscopului si prin insamantari pe suprafata mediului nutritiv, solidificat in placi Petri. Prin controlul vizual al eprubetei, se poate observa uniformitatea cresterii si prezenta indicatorilor morfologici caracteristici pentru specia izolata. Prin control microscopic, in preparate umede se observa forma celulelor si absenta microorganismelor de contaminare.
Cultura pura de laborator se analizeaza si din punct de vedere al aspectului, a numarului de celule moarte, pentru a avea siguranta ca este corespunzatoare. Drojdia pura trebuie sa fie sedimentata intr-un strat compact pe fundul vasului; cand drojdia este raspandita in masa lichidului si aglomerata in flocoane vizibile, denota faptul ca mediul de cultura a fost contaminat. Celulele de drojdie moarte se identifica cu ajutorul metodei de colorare cu solutie de albastru de metilen.
In vederea asigurarii de parametri cat mai uniformi in decursul multiplicarii in productie, este necesara conservarea culturilor pure de drojdii. In momentul constatarii unor fenomene de degenerare, de contaminare sau aparitiei de mutante care modifica insusirile drojdiilor, precum si a unei proportii prea mari de celule moarte, se procedeaza la schimbarea culturii. Un laborator specializat in prepararea, conservarea si livrarea de culturi pure dispune de o micoteca cu o gama larga de tulpini de drojdii cu caracteristici bine cunoscute, care le livreaza la cererea producatorilor de drojdie de panificatie.
Dintre tehnicile aplicate pentru conservarea culturilor pure, bazate pe prelungirea fazei stationare de crestere si evitarea etapei de declin, se cunosc urmatoarele:
repicarea periodica prin transfer de celule din eprubeta cu cultura pura in care mediul nutritiv este epuizat, in eprubeta cu mediul nutritiv steril cu compozitie similara. Metoda necesita un mare volum de munca, prezinta risc de contaminare si este greu de realizat atunci cand numarul du culturi este mare;
prelungirea intervalului intre doua repicari prin scaderea vitezei de metabolism a celulelor se poate realiza prin:
mentinerea culturilor la temperaturi scazute, in conditii de refrigerare sau congelare;
privarea de oxigen: cultura dezvoltata in mediu solidificat se acopera cu un strat de ulei de parafina steril, prevenindu-se in acest fel si uscarea mediului;
reducerea umiditatii mediului conduce la trecerea celulelor in stare de anabioza care poate fi mentinuta timp indelungat fara a se produce modificari intracelulare, ireversibile;
pastrarea culturilor in stare liofilizata, este tehnica cea mai raspandita si avantajoasa deoarece prelungeste cel mai mult intervalul de conservare a culturilor, fara sa produca modificari ale proprietatilor lor fiziologice (Dan, V., 1999).
Scopul selectionarii unei tulpini de drojdie este obtinerea in laborator a unor culturi pure de celule cat mai omogene, in ceea ce priveste metabolismul, randamentul, viteza de inmultire, capacitatea de reproducere si calitatea produsului finit.
Principalele conditii pe care trebuie sa le indeplineasca o cultura pura de laborator, destinata fabricarii drojdiei de panificatie sunt urmatoarele (Dan, V., 1999):
sa posede o capacitate mare de fermentare, indiciu ca tulpina de drojdie contine tot complexul enzimatic. Aceasta este o proprietate complexa si reflecta activitatea combinata a produselor mai multor gene. Foarte important pentru industria panificatiei este utilizarea de drojdii cu potential inalt de fermentare a maltozei;
sa posede o rata de crestere si randament pe medii cu noi surse de carbon;
sa fie pura din punct de vedere microbiologic;
sa fie viguroasa pentru a rezista diferitelor microorganisme de contaminare;
sa suporte o concentratie cat mai mare de saruri, dat fiind compozitia melasei;
sa fie acomodata la anumite doze de conservanti, care sa inhibe microorganismele de contaminare;
sa posede rezistenta la congelare si uscare;
sa-si pastreze proprietatile biotehnologice timp indelungat. Aceasta proprietate este conditionata genetic.
Selectionarea consta in izolarea unei singure celule, careia i se creeaza apoi conditii de multiplicare pana la obtinerea unei cantitati ce este testata intr-o statie pilot, in vederea stabilirii insusirilor care caracterizeaza tulpina respectiva de drojdie. Daca aceasta prezinta caracteristicile dorite, devine una din tulpinile de baza, care se multiplica in fazele de laborator si apoi se introduce in fabrica. Producatorii de drojdie de panificatie acorda o atentie deosebita conservarii in conditiile impuse a tulpinii de drojdie, ca aceasta sa fie ferita de contaminare si pastrata la temperatura prescrisa pentru a evita aparitia unor mutatii care ar influenta negativ calitatea si randamentul drojdiei finite.
In prezent, datorita cunostintelor din domeniul geneticii s-au pus bazele ameliorarii drojdiilor industriale si modalitatilor practice de imbunatatire a calitatilor lor prin inducerea, identificarea, izolarea si caracterizarea unor mutante si linii cu proprietati biologice si economice superioare, obtinerea unor hibrizi si recombinati prin tehnici de inginerie genetica si fuziune de protoplasti.
Aprecierea concentratiei de celule de drojdie din medii lichide prin metode directe. Pentru aprecierea numarului de celule de drojdie apartinand speciei Saccharomyces cerevisae in medii de multiplicare sau de fermentare si evaluarea vitezei de crestere se pot folosi metode de numarare directe cu ajutorul camerelor de numarare. De exemplu, se poate utiliza camera Thoma. Aceasta camera prezinta o retea cu suprafata de 1 mm2 divizata in 400 microcelule elementare si o adancime de 0,1 mm incat volumul suspensiei de celule pentru numarat corespunzator unei microcelule este de 1/4000 mm3.
Relatia de calcul folosita in determinari va fi:
N= n x 4 x 106 x K
in care:
n = numar mediu de celule/microcelule;
4.106 = factor de transformare in cm3 a volumului microcelulei elementare;
k = factor de dilutie(facultativ);
N = numar celule/cm3 suspensie).
Determinarea viabilitatii celulelor de drojdie. Metoda se aplica frecvent pentru evaluarea calitatii drojdiei comprimate, a starii fiziologice si determinarea procentuala a celulelor autolizate, a influentei unor factori de mediu asupra activitatii fiziologice a celulei de drojdie. In principiu, metoda se bazeaza pe faptul ca celulele de drojdie neviabile fie in urma procesului de autoliza fie a unui factor exterior ce produce modificari ireversibile in structura proteinelor, isi pierd proprietatile fermentative. Prin suspendarea celulelor de drojdie in solutie diluata de albastru de metilen (1:10.000 in citrat de sodiu 2%), celulele inactive fiziologic, se vor colora in albastru deoarece in urma inactivarii reductazelor, nu se mai produce trecerea colorantului in forma sa de leucoderivat incolor, asa cum are loc in celula vie, activa. Cu ajutorul camerei Thoma se poate face numararea separata a celulelor neviabile (autolizate), din total celule si exprimarea lor procentuala / cm3 sau grame drojdie comprimata.
2.3. Multiplicarea drojdiei in laborator
Multiplicarea celulelor de drojdiei se efectueaza in doua etape, in laborator si apoi in fabrica.
Se pleaca de la o cultura pura de drojdie obtinuta la un institut specializat sau chiar in laboratorul fabricii prin metoda izolarii in picaturi sau in placi. Cultura de drojdie de baza se pastreaza pe must de malt cu agar la intuneric si la temperaturi scazute de 2÷50C luandu-se toate masurile de a o feri de contaminare cu microorganisme straine.
Multiplicarea culturii de drojdie in laborator are loc in patru faze, folosindu-se ca mediu de cultura must de malt. In prima faza se prepara cultura de drojdie intr-o eprubeta de 20 ml, in care se introduce mediul nutritiv care se solidifica in plan inclinat si se insamanteaza drojdia cu varful de platina al ansei preluata din tulpina selectionata. Din aceasta eprubeta se insamanteaza in vase Erlenmayer cu cresterea succesiva a volumului de must de malt, in trei faze, la intervale de 24 de ore.
La sfarsitul multiplicarii se obtine cultura pura de drojdie de laborator care serveste pentru insamantarea in primul vas de multiplicare a drojdiei in sectia de culturi pure.
2.4. Multiplicarea drojdiei in fabrica
Multiplicarea in fabrica are loc in cinci faze, primele doua faze in vase de multiplicare in statia de culturi pure, iar urmatoarele trei faze in linuri de multiplicare. Principalii parametri tehnologici in procesul de multiplicare a drojdiilor de panificatie, in cazul procedeului clasic sunt prezentati in tabelul 23.
Tabelul 23
Principalii parametri tehnologici in procesul de multiplicare a drojdiei de panificatie - procedeul clasic
|
Faza de multiplicare |
Concentratia plamezii |
Temperatura [0C] |
pH |
Durata de multiplicare [ore] |
|
|
Initiala [0Bllg] |
Finala [0Bllg] |
||||
|
Faza I | |||||
|
Faza a II-a | |||||
|
Faza a III-a | |||||
|
Faza a IV-a | |||||
|
Faza a V-a | |||||
Statia de culturi pure a fabricii asigura multiplicarea in doua trepte, in vase metalice, cu cresterea succesiva a volumului de 5÷10 ori. Ca mediu nutritiv se foloseste o solutie apoasa de melasa cu adaos de substante nutritive, denumita plamada. Pentru realizarea unei culturi viguroase, se urmareste multiplicarea celulelor de drojdie, concomitent cu o fermentatie alcoolica, intr-un mediu cu a aciditate ridicata.
Pentru faza I de multiplicare a drojdiei se utilizeaza vase de multiplicare, confectionate din cupru, prevazute cu racord de apa, abur, aer, gura de vizitare cu capac, robinet de prelevare probe, conducta de eliminare CO2, cu o capacitate de 300÷500 l/buc.
Vasul de multiplicare este mai intai curatat, spalat si sterilizat cu abur si formalina, dupa care se prepara mediul nutritiv, conform retetei de fabricatie, corectia de pH realizandu-se cu H2SO4 concentrat, pana la un pH de 4,0÷5,0. Plamada obtinuta se sterilizeaza cu abur direct timp de o ora, dupa care se raceste cu ajutorul sistemului exterior de racire la 28÷320C, apoi se insamanteaza plamada cu cultura pura de laborator.
Multiplicarea are loc prin fermentare aeroba cu formare de alcool, vasul fiind inchis cu capac. In timpul perioadei de fermentare din doua in doua ore se executa controlul temperaturii, gradului Balling, aciditatii si examenul microscopic al plamezii.
Continutul vasului este trecut integral prin conducta de legatura, sterilizata cu abur in prealabil, in vasul din faza a II-a culturi pure fabrica cu o capacitate de 1000÷2500 l.
Plamada pregatita conform retetei de fabricatie se sterilizeaza cu abur direct timp de o ora. Se raceste plamada la 28÷320C si se insamanteaza cu drojdie din faza I de multiplicare. Cultura pura de fabrica obtinuta este folosita integral pentru insamantarea in cea de-a treia faza de multiplicare a drojdiei.
Instalatia pentru obtinerea culturii pure este prezentata in figura 21.
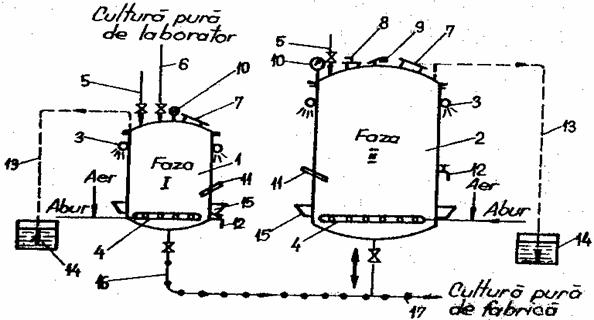
Vasele sunt prevazute cu tevi exterioare perforate 3, prin care se poate introduce apa rece sau calda pentru temperarea plamezii si cu tevi perforate in interior 4 prin care se poate introduce abur pentru sterilizarea mediului cat si aer comprimat in timpul multiplicarii drojdiei.
Vasele mai sunt prevazute cu racorduri pentru introducerea mediului nutritiv 5, racordul de insamantare cu cultura pura de laborator 6, guri de vizitare 7, supape de suprapresiune 8 si de vacuum 9, manometre 10, termometre 11, robinete de prelevare probe 12 si conducte de evacuare a dioxidului de carbon 13 care patrund in vasele de apa 14. Apa de racire ce se prelinge pe peretii exteriori este colectata si evacuata din jgheabul 15. Cultura de drojdie obtinuta in primul vas este trecuta prin conducta 16 in cel de-al doilea vas, iar cultura pura rezultata din acest vas trece prin conducta 17 in sectia de productie (Hopulele, T., 1980).
Drojdia obtinuta in statia de culturi pure este multiplicata in continuare in fabrica in 2÷4 faze, in functie de tehnologia si utilajele folosite. Se practica procese cu plamezi de melasa diluata (1/18÷1/25) sau concentrata (1/5÷1/10) si tehnici de multiplicare discontinua sau continua. Randamentele obtinute difera in functie de caracteristicile materiilor prime, a culturii de drojdie si a tehnologiei aplicate. In unele cazuri se urmareste producerea concomitenta de drojdie comprimata si de alcool etilic. Spre deosebire de primele doua faze in care mediul nutritiv este introdus la inceputul multiplicari integral in vasul de multiplicare, la multiplicarea drojdiei in fazele III÷IV, alimentarea cu melasa si substante nutritive se realizeaza in mod continuu, dupa diagrame de alimentare prestabilite, care trebuie respectate cu strictete.
In faza a III-a de multiplicare, capacitatea linurilor este de circa 10 ori mai mare decat a vaselor folosite in faza a II-a (7÷25 m3). Capacitatea utila reprezinta numai 75% din cea totala, restul de 25% fiind afectat pentru sistemul de aerare cat si pentru spuma formata.
Inainte de utilizare linurile se curata, se spala cu solutie de soda caustica 2÷4% si in final se face o sterilizare combinata cu abur si solutie de formalina 5÷10% timp de circa o ora. Se introduce apoi apa in lin pana la 50% din capacitatea utila a acestuia, se adauga 1/3 din melasa pregatita si o parte din substantele nutritive. Se omogenizeaza prin barbotare de aer si se insamanteaza cu drojdie rezultata din faza a II-a de multiplicare.
In timpul multiplicarii, spuma se combate cu substante antispumante care se introduc direct in plamada. Se respecta diagrama orara de alimentare cu melasa si substante nutritive a linului de multiplicare.
Indiferent de tehnologia aplicata, in instalatii de mare capacitate, plamada de drojdie rezultata in treapta a III-a de multiplicare este supusa concentrarii cu separatoare centrifugale inainte de insamantare pentru urmatoarea etapa de multiplicare. Totodata, se corecteaza pH-ul si se pastreaza laptele de drojdie obtinut in recipiente racite la temperatura de 4÷60C.
Multiplicarea drojdiei in faza a IV-a are loc in linuri asemanatoare din punct de vedere constructiv cu faza a III-a, avand insa capacitatea de 5÷6 ori mai mare (40÷100 m3). In aceasta faza se obtine drojdia cuib sau drojdia maia folosita pentru insamantarea mediului nutritiv din ultima faza de multiplicare (faza a V-a).
Conditiile de multiplicare a drojdiei in aceasta faza sunt mai favorabile decat in
fazele precedente:
concentratia si aciditatea mediului sunt mai reduse;
aerarea mediului este mai intensa;
procentul de alcool din plamada este foarte redus.
Pentru stabilirea cantitatii necesare de melasa pentru aceasta faza este necesar sa se tina seama de raportul de dilutie, care reprezinta raportul dintre cantitatea de melasa nediluata (tone) si volumul final al plamezii exprimat in m3. In faza a IV-a de multiplicare raportul de dilutie trebuie sa fie de circa 1/1 De exemplu, pentru o capacitate utila a linului de 75 m3 necesarul de melasa va fi 75:18 = 4,166 tone.
Dupa ce linul a fost spalat si sterilizat se introduce apa in proportie de circa 30% din volumul util, peste care se adauga 15% din cantitatea de melasa pregatita si 33% din cantitatea de substante nutritive pregatite sub forma de solutie, se omogenizeaza prin barbotare de aer si se face insamantarea prin conducta de legatura cu plamada de drojdie din faza a III-a. Restul de melasa si substante nutritive din reteta de fabricatie se adauga in timpul multiplicarii drojdiei. Astfel, in prima ora de multiplicare nu se adauga melasa si substante nutritive, drojdia aflandu-se in faza latenta a ciclului vital. Din acest motiv si debitul de aer este mai redus de 50 m3/m3 plamada si ora.
Incepand din ora a doua, cand drojdia intra in faza logaritmica de multiplicare, incepe adaugarea de melasa si substante nutritive in cantitati din ce in ce mai mari, dupa o diagrama prestabilita. In aceasta perioada de multiplicare intensa a drojdiei se foloseste un debit maxim de aer de 100 m3/m3 plamada si ora. In ultima ora nu se mai efectueaza alimentarea cu melasa si substante nutritive, debitul de aer scade la valoarea initiala, drojdia fiind lasata sa se maturizeze.
Plamada de drojdie rezultata din faza a IV-a nu se insamanteaza ca atare in faza a V-a, ci sub forma de lapte de drojdie obtinut prin separare centrifugala si pastrat pana la folosire, la temperatura de 0÷40C in colectoare de depozitare. Laptele de drojdie obtinut mai este denumit impropriu si maia, deoarece el serveste la insamantarea plamezilor din faza a V-a de multiplicare.
In aceasta ultima faza de multiplicare a drojdiei se obtine asa numita drojdie de vanzare. Multiplicarea are loc in linuri identice ca in faza a IV-a, folosindu-se circa 80% din capacitatea totala de fermentare pentru drojdia de vanzare, restul de 20% utilizandu-se pentru obtinerea drojdiei maia. Astfel la intervale de 2÷3 zile unul sau doua linuri sunt folosite pentru producerea drojdiei maia.
In ultima faza de multiplicare se asigura cele mai bune conditii de mediu pentru dezvoltarea drojdiei, concentratia si aciditatea plamezii fiind mai reduse decat in faza a IV-a de multiplicare. Se urmareste obtinerea unei puritati microbiologice crescute a plamezii si a unor randamente maxime.
In faza a V-a de multiplicare raportul de dilutie este de 1/25. Se introduce la inceput intreaga cantitate de apa in linul de multiplicare, adaugand apoi 8% din melasa necesara si 14% din cantitatea de substante nutritive, apoi se respecta diagramele orare de alimentare stabilite.
Din colectorul de depozitare maia se insamanteaza linul de multiplicare din faza a V-a cu o portie de maia egala cu ¼ sau 1/5 din volumul total rezultat de maia si se omogenizeaza plamada prin barbotare cu aer. Astfel, cu o maia se pot insamanta concomitent 4 sau 5 linuri de faza a V-a. In timpul multiplicarii se controleaza orar concentratia, aciditatea si temperatura, efectuandu-se corectiile necesare, iar la doua ore se efectueaza si un control microscopic al drojdiei.
2.5. Linurile de multiplicare a drojdiei
Linurile de multiplicare a drojdiei in diferite faze, denumite si fermentatoare, constituie utilajele principale folosite la fabricarea drojdiei de panificatie. Ele pot fi confectionate din tabla de otel antiacid, otel inoxidabil sau chiar din otel obisnuit protejat in interior cu un lac acidorezistent.
Fig.22. Lin de multiplicare a drojdiei prevazut cu sisteme static de aerare:
1 - sisteme de aerare;
2 - serpentina de racire;
3 - racord de alimentare cu drojdie;
4 - palnie de alimentare cu melasa;
5 - conducta de aer;
6 - conducta de evacuare a aerului uzat;
7 - conducta de abur;
8 - conducta de evacuare a plamezii de drojdie.
Linurile pot avea forma cilindrica sau paralelipipedica. Forma cilindrica permite o distributie mai uniforma a aerului in plamada si o curatire mai usoara, fiind astfel cea mai des intalnita. Linurile de forma paralelipipedica permit o utilizare mai buna a spatiului de la fermentare. Schematic un lin clasic de multiplicare a drojdiei prevazut cu un sistem static de aerare se prezinta in figura 22.
Sistemul de aerare este format dintr-o conducta centrala verticala pentru intrarea aerului care este in legatura cu o conducta orizontala amplasata la fundul linului, in care sunt infiletate o serie de tevi perforate, dispuse pe toata suprafata fundului linului astfel incat sa permita o distributie cat mai fina si mai uniforma a aerului in mediu, de care depinde in cea mai mare masura gradul de utilizare a oxigenului si deci consumul specific de aer. Orificiile de distributie a aerului au un diametru de 0,4÷0,5 mm, distanta dintre ele este de circa 4 mm si sunt amplasate pe partea laterala a tevilor perforate. Acestea sunt prevazute la capete cu capace infiletate, care se pot scoate pentru curatire si spalare.
O importanta deosebita are sistemul constructiv al instalatiei de aerare, solubilizarea oxigenului in masa variind de peste 10 ori la diversele instalatii, functie de particularitatile constructive (Anghel, I. et al., 1991). Progrese importante in tehnica aerarii au fost realizate dupa elaborarea de instalatii rotative. Dintre instalatiile care s-au impus in practica, se pot enumera: inferatorul, aeratorul Vogelbusch, sistemul de aerare cu jet adanc (VB-IZ), sistemul de aerare Frings.
Fig. 23. Lin de multiplicare a drojdiei tip Vogelbusch:
1 - rotor cu palete radiale (dispergator);
2 - motor electric;
3 - cutie de viteze;
4 - conducta admisie aer in lin;
5 - sisteme de racire prin stropire exterioara;
6 - jgheab colector;
7 - spargator de spuma;
8 - conducta pentru spuma;
9 - conducta de plamada;
10 - conducta evacuare aer uzat;
11 - conducta alimentare cu plamada;
12 - conducta evacuare plamada de drojdie.
Prin folosirea sistemelor dinamice de aerare s-au putut utiliza la multiplicarea drojdiilor plamezi mult mai concentrate decat in cazul procedeului clasic, obtinandu-se in final randamente superioare de biomasa de drojdie de 4÷5 ori mai mare.
Linurile moderne de fermentare sunt prevazute cu instalatii complexe de automatizare, care permit reglarea automata a alimentarii cu melasa si substante nutritive, a debitului de aer, apei tehnologice, antispumant, masurarea si reglarea automata a pH-ului plamezii si a temperaturii in lin, prin variatia debitului de apa de racire.
Intrucat multiplicarea drojdiei este un proces exoterm, eliberandu-se 2500÷3500 kcal/kg s.u. de drojdie, este necesara o racire corespunzatoare a plamezii care se poate realiza cu ajutorul serpentinelor de racire, a unor baterii de tevi demontabile verticale asezate in interiorul linului sau prin stropire exterioara. Atat sistemul de distribuire a aerului cat si cel de racire sunt construite din teava de cupru.
Datorita aerarii intense si a substantelor coloidale din melasa, in timpul multiplicarii drojdiei se formeaza cantitati mari de spuma, pentru combaterea careia se utilizeaza doua grupe de procedee:
procedee mecanice, care se bazeaza pe folosirea unor spargatoare de spuma;
procedee chimice, care utilizeaza pentru distrugerea spumei substante cu actiune antispumanta.
In fabricile de drojdie spuma se combate de obicei prin folosire de substante antispumante.
2.6. Factorii care influenteaza calitatea drojdiei in fazele de multiplicare
2.6.1. Efectul presiunii osmotice
Drojdiile se dezvolta in conditii bune, cand mediul in care se afla are o presiune osmotica cat mai apropiata de aceea din interiorul celulei (izotonie). Schimbarile bruste si importante ale presiunii osmotice a mediului pot provoca dereglarea functiilor compensatoare de adaptare ale membranei citoplasmatice si chiar lezari ale peretelui celular, ce pot duce la moartea fiziologica a celulei. In medii cu presiune osmotica ridicata, bogate in glucide sau saruri (medii hipertonice), celulele sunt silite sa realizeze in interiorul lor o contrapresiune osmotica echivalenta, lasand sa treaca in mediu o proportie corespunzatoare de apa.
Cand celulele se gasesc in medii cu presiune osmotica inferioara celei a continutului vacuolelor, in apa de exemplu, din aceeasi necesitate a realizarii unei contrapresiuni osmotice echivalente, accepta patrunderea de apa din mediul extern. Drept urmare, turgescenta celulelor creste pana cand presiunea intracelulara depaseste rezistenta peretelui, care plesneste. Fenomenul de turgescenta duce astfel la distrugerea celulelor. Prin deshidratarea drojdiilor, in celule se mareste concentratia in substante si creste presiunea osmotica, ce exercita influenta asupra proceselor fermentative. La marirea concentratiei in substante incepe franarea proceselor biochimice ale celulei si la un anumit nivel incepe socul osmotic (Dan,V., 1999).
2.6.2. PH-ul mediului
Drojdiile se dezvolta in limite largi de pH, pentru ca au capacitatea sa se adapteze la unele modificari ale mediului de cultivare. Astfel, daca pH-ul mediului este mai acid decat valoarea optima pentru crestere, in celula devin active enzimele decarboxilaze, cand pH-ul este mai bazic, decat valoarea optima, devin active dezaminazele. In aceste conditii, produsele rezultante din aminoacizi sub actiunea catalitica a acestor sisteme enzimatice tind sa realizeze neutralizarea si reprezinta sisteme tampon al efectului nociv al pH-ului. Dupa epuizarea stocului de aminoacizi, actiunea pH-ului duce la moartea celulelor, ca rezultat al unui dezechilibru, prin modificarea schimburilor osmotice intre celula si mediu.
Efectul pH-ului mediului nutritiv asupra multiplicarii drojdiilor este cunoscut de mult timp si valorificat in practica. Actiunea sa asupra celulelor de Saccharomyces cerevisiae a fost studiata de multi cercetatori. La un pH = 7,5 intensitatea de respiratie si randamentul de crestere este cu 60÷100 % mai mare decat la pH = 4 in diverse medii nutritive cu glucoza la 300C. Cu scaderea pH-ului in mediul nutritiv se stimuleaza patrunderea protonilor in celule. La un pH = 3,5 si cantitati suficiente de saruri de potasiu in mediu nutritiv, creste pH-ul intracelular, ajungand la valorile de 7,5. Variatia pH-ului intracelular are o importanta deosebita la reglarea glicolizei si a respiratiei celulelor de drojdie (Anghel, I. et al., 1991).
Valoarea optima a pH-ului la cultivarea drojdiei Saccharomyces cerevisiae oscileaza intre 4,5÷5,8, desi drojdiile sunt mult mai active intr-un mediu care are o valoare a pH-ului de 7÷7,5. Celulele de drojdii in acest domeniu se gasesc in stare fiziologica buna si se inmultesc rapid. De nivelul de pH in timpul cultivarii drojdiilor, depinde randamentul si calitatea produselor finite. In practica, dezvoltarea drojdiilor se realizeaza in mediu acid, concentratia mai mare in hidrogen fiind un mijloc de combatere a microorganismelor de contaminare.
Domeniul de pH in care drojdia se poate multiplica este influentat de compozitia mediului si de continutul in alcool al acestuia. Intr-un mediu de fermentare cu 4,5 % alcool, drojdiile pot sa actioneze pana la pH = 1, La un continut de 5,5÷6 % alcool, valoarea minima a pH-ului suportat de drojdii este de 2,3, iar la un continut de 8,5÷12,5 % alcool, limita inferioara a pH-ului la care drojdia poate actiona este de 3,5, ritmul de crestere la acest pH fiind incetinit. In afara de aceasta, in intervalul de pH 3÷3,5 se afla punctul izoelectric al unor substante colorante din melasa, care sunt absorbite de catre celulele de drojdie (Anghel, I. et al., 1991).
Schimbarea regimului de pH exercita actiune asupra activitatii enzimelor, asupra patrunderii substantelor nutritive in celula de drojdie si se intensifica respiratia. Brusc se franeaza schimbul de aminoacizi in celula de drojdie, scade cantitatea de biomasa rezultata, se inrautateste calitatea drojdiei. Valorile extreme de pH (medii puternic acide sau puternic alcaline) provoaca denaturarea ireversibila a enzimelor.
Sunt date care arata ca marirea pH-ului provoaca cresterea activitatii enzimelor, care participa la formarea poliglucidelor, solubile in acizi. Sinteza maxima a trehalozei s-a observat la cultivarea drojdiei Saccharomyces cerevisiae in mediul cu pH 4,5÷5,0.
La fabricarea drojdiei de panificatie se scade pH-ul la valori de circa 4, in primele faze de multiplicare (faza I si a II-a), urmarindu-se o acumulare de biomasa celulara activa. In continuare, pe masura progresarii numarului de faze de multiplicare, pH-ul creste pana la valoarea de 5,5.
Continutul in azot al componentelor nutritive contribuie la normalizarea pH-ului mediului. Din sulfatul de amoniu, drojdia asimileaza NH3 si elibereaza in mediu acid sulfuric. Adaosul de apa amoniacala neutralizeaza acidul sulfuric, compenseaza pH-ul, si, in acelasi timp, furnizeaza drojdiei necesarul de azot.
Corectarea pH-ului in industria drojdiei de panificatie cu ajutorul acidului lactic este mai favorabila celulelor de drojdie. Mai putin favorabil actioneaza acidul fosforic si acidul clorhidric, iar acidul sulfuric este pe ultimul loc. La acidifierea cu acid lactic s-a obtinut o drojdie cu putere de crestere de 10 minute, biomasa 32 g·dm-3 si o activitate maltazica de 304 U.A.
2.6.3. RH-ul mediului
Drojdiile prezinta diferite grade de sensibilitate la potentialul de oxidoreducere. Dintre substantele care ajuta la mentinerea unui potential de oxidoreducere redus, sunt acidul ascorbic, glucidele reducatoare si substante ce contin grupele - SH. Fiecare sistem biologic are in compozitia sa, atat substante oxidante cat si reducatoare, incat valoarea potentialului de oxidoreducere, este in functie de raportul intre ele si mai depinde de tensiunea de oxigen si de valoarea pH. La modificari ale rH-ului se pot produce modificari in metabolismul celular, sau in cazul unor valori limita, este oprita cresterea.
2.6.4. Temperatura
Temperatura este, din punct de vedere al procesului de biosinteza desfasurat la scara industriala, unul dintre parametrii fizici cei mai importanti, implicat profund, prin efectele sale, in optimizarea procesului. Variatiile temperaturii au efect asupra randamentului de transformare a substratului in produsul dorit, asupra cerintelor nutritive ale drojdiei, compozitiei biomasei obtinute si vitezei de crestere (Anghel, I. et al., 1991).
In cursul evolutiei, celulele de drojdie au suferit numeroase adaptari, incat in lumea microbiana, drojdiile se multiplica la temperaturi variate in limite foarte largi.
Drojdia Saccharomyces cerevisiae apartine grupului mezofil, temperatura optima oscileaza intre 260C si 360C. Datele din literatura de specialitate arata ca, drojdia de panificatie cu puterea de crestere cea mai buna se obtine la temperatura de 300C.
Deplasarea cu cateva grade in jurul temperaturii optime de crestere influenteaza nu numai randamentul in biomasa obtinuta si viteza de crestere, dar si compozitia biochimica a celulei de drojdie. Datele din literatura publicate arata ca variatiile de temperatura afecteaza multe procese metabolice din celula, precum si compozitia biomasei in proteine si lipide, continutul in ARN al celulei (Hunter si Rose, 1972). Raportul dintre continutul in ARN al drojdiilor si viteza lor de crestere se mareste la scaderea temperaturii.
Temperatura procesului de cultivare conditioneaza continutul in lipide din compozitia membranelor celulare. Astfel, membrana drojdiilor psihrofile contine in cantitati mai mari acizi grasi polinesaturati, cele termofile, acizi grasi mononesaturati, iar cele mezofile acizi mono- si polinesaturati.
Celulele de drojdie pot suporta temperaturi foarte scazute, pana aproape de zero absolut. Ele supravietuiesc mai usor la rece intr-un mediu uscat, decat intr-unul umed. S-a observat ca prin scaderea temperaturii sub limita inferioara de 00C se constata o reducere a vitezei de metabolism. Astfel, prin scaderea cu 100C sub temperatura minima are loc o scadere cu 50% a vitezei de metabolizare a substantelor nutritive. Aceasta scadere de activitate se explica prin faptul ca, prin reducerea temperaturii are loc o pliere a lanturilor proteice si mascarea centrilor activi ai enzimelor incat acestea nu mai fac legatura cu substratul si nu mai indeplinesc functia de biocatalizatori. La temperaturi scazute, se produc pierderi de apa intracelulara, drojdiile trec in stare latenta de viata, cand metabolismul se desfasoara foarte lent si pot ramane viabile timp indelungat. Prin congelare, drojdiile se pot pastra timp nelimitat, deoarece cantitatea de apa libera in exteriorul si interiorul celulei se reduce, trecand in stare solida si numai o parte ramanand disponibila pentru a fi folosita de celule, incat activitatea celulei este oprita. Campbell a descoperit ca o congelare a drojdiei Saccharomyces cerevisiae suspendata in apa la -300C sau -500C a distrus 48÷94 % din celule. Dezghetarile si inghetarile repetate au provocat moartea celulelor de drojdie. Aceste rezultate au fost confirmate de Baum cu o cultura pura de Saccharomyces cerevisiae. Efectele multiple ale inghetarii asupra celulelor de drojdie au fost descrise de Mazur(1966).
Scaderea temperaturii de cultivare de la 300C la 150C contribuie la marirea continutului de lipide, la 300C el este de 12 %, iar la 150C este de 14,5 %. La Saccharomyces cerevisiae, prin scaderea temperaturii de crestere mai jos de optim, se mareste cantitatea de proteine si acizi ribonucleici, iar cantitatea totala de glucide ale celulei scade, in principal, prin scaderea continutului de trehaloza (Cernis, V.G., 1975).
S-a constatat ca drojdia obtinuta la temperaturi scazute (29÷300C) este mai bogata in azot si fosfor si mai rezistenta la pastrare. La temperaturi ridicate se inrautateste brusc calitatea productiei, ca o consecinta a faptului ca poate avea loc, ocazional, multiplicarea drojdiilor atipice si a microbiotei bacteriene. Microorganismele contaminante consuma substratul, destinat pentru drojdie, ceea ce conduce la scaderea randamentului.
Obtinerea drojdiei de panificatie la temperaturi mai ridicate de 300C se face cu acumulare in celula de drojdie a trehalozei. Acumularea maxima de trehaloza in celule s-a obtinut prin cultivarea drojdiei la temperaturi de 400C. Cresterea in continuare a temperaturii nu numai ca a oprit considerabil multiplicarea drojdiei, dar a provocat si scaderea cantitatii de trehaloza in celule. Marirea cantitatii de trehaloza in celule la temperaturi ridicate de crestere este insotita de scaderea glicogenului.
Viteza de acumulare a alcoolului, de asemenea, creste cu marirea temperaturii, atinge un maxim la 400C, dupa aceea scade brusc la cresterea mai departe a temperaturii.
Cresterea temperaturii de cultivare duce la reducerea randamentului in drojdie ca rezultat al scaderii in umiditate si ca o consecinta, marirea consumului de substante nutritive la fermentare. Prin ridicarea temperaturii de crestere se inrautateste aprovizionarea celulei cu acizi, se mareste excretia aminoacizilor liberi in lichidul cultural si de asemenea creste necesitatea drojdiilor in vitamine. Acest fapt poate provoca modificarea coeficientului economic.
In domeniul de temperatura supraoptimala, cresterea este concurata de moartea microorganismelor. Moartea celulelor de drojdie la temperaturi supramaximale se datoreaza denaturarii termice a proteinelor si enzimelor celulare, incat activitatea de metabolism este oprita si se produce moartea fiziologica a celulei fara sa aiba loc distrugerea fizica.
Liza drojdiilor este o functie puternic dependenta de temperatura, fiind caracterizata prin valori ale energiei de activare de 70÷90 kcal/mol.
2.6.5. Umiditatea
Apa este importanta pentru celula de drojdie nu numai pentru ca este principalul constituent din punct de vedere cantitativ, ea reprezentand circa 80% din greutatea celulei vii, ci si pentru ca indeplineste urmatoarele functii (Pirt,1975):
ca reactant chimic prezent in celula, apa participa la reactiile de hidroliza;
actioneaza ca solvent pentru metabolitii intracelulari;
rol mecanic important in mentinerea formei si dimensiunilor celulei impuse de presiunea hidrostatica care ia nastere in interiorul celulei;
indeplineste o functie structurala in hidratarea proteinelor si a altor componente celulare.
De exemplu, de gradul de hidratare a mitocondriilor depinde intensitatea proceselor de fosforilare oxidativa care au loc in aceste organite. Apa participa direct la formarea citoplasmei celulare, de a carei stare depinde functia sa fiziologica. Formeaza legaturi de H si participa la structura unor compusi macromoleculari (Anghel, I. et al., 1993).
Unele reactii chimice, biochimice, enzimatice si in special microbiologice sunt strans corelate cu umiditatea substratului. In conditiile in care umiditatea scade, se reduce viteza de metabolism, enzimele trec in stare inactiva si celulele de drojdie se mentin viabile in stare de anabioza.
Drojdiile pot folosi numai apa libera, nelegata de componentii chimici ai mediului si nu pot folosi apa legata chimic sau fizic. Drojdiile necesita, pentru o dezvoltare normala, o cantitate de apa libera care sa asigure un transfer corespunzator al substantelor nutritive in celula. Marea majoritate a drojdiilor nu pot sa se dezvolte in medii cu indice de activitate al apei (aw) inferior valorii 0,90, dar exista si drojdii osmotolerante care rezista la presiuni osmotice mai ridicate corespunzatoare unui aw = 0,60 (Dan, V., 1999).
Concentratia in oxigen si influenta asupra bioenergeticii levuriene
Drojdiile de panificatie sunt facultativ anaerobe, ele sunt capabile de activitate vitala in conditii anaerobe si aerobe.
Viteza de respiratie se
masoara in cantitatea de oxigen absorbita de substanta uscata a drojdiei, si notata ![]() ,iar viteza de fermentare reprezinta cantitatea de
dioxid de carbon degajata la 1 g substanta uscata drojdie intr-o ora. In
conditii aerobe, aceasta valoare s-a notat
,iar viteza de fermentare reprezinta cantitatea de
dioxid de carbon degajata la 1 g substanta uscata drojdie intr-o ora. In
conditii aerobe, aceasta valoare s-a notat![]() , iar in anaerobioza (in atmosfera de azot) -
, iar in anaerobioza (in atmosfera de azot) - ![]() . In conditii aerobe se produce efectul Pasteur care s-a
notat PE, realizandu-se blocarea fermentarii in favoarea respiratiei.
Gradul de blocare depinde, intr-o masura considerabila, de
caracteristicile culturii si se exprima prin egalitatea:
. In conditii aerobe se produce efectul Pasteur care s-a
notat PE, realizandu-se blocarea fermentarii in favoarea respiratiei.
Gradul de blocare depinde, intr-o masura considerabila, de
caracteristicile culturii si se exprima prin egalitatea:
![]()
Drojdiile, in functie de conditiile de cultivare, aerobe sau anaerobe, pot suferi modificari morfologice ale celulei. In conditii de anaerobioza celulele nu sunt capabile sa sintetizeze tipul de citocromi -a1, a3, b, c1, se dezechilibreaza transportul electronilor in lantul respirator si scade activitatea enzimelor respiratorii. In conditii de crestere in anaerobioza, in celule nu se produce sinteza acizilor grasi nesaturati si a ergosterolului (Novakovskaia, S.S., Sisatkii, I.I., 1980).
Pentru functionarea normala a lantului respirator si desfasurarea activitatii enzimelor respiratorii mitocondriale e necesara prezenta in mitocondrii a unei cantitati definite de lipide, care sa contina acizi grasi nesaturati.
Alternanta conditiilor anaerobe de cultivare cu cele aerobe la Saccharomyces cerevisiae duce la formarea tuturor componentelor celulare, activarea enzimelor respiratorii si biosinteza mitocondriilor.
In ultimii ani s-au obtinut date despre faptul ca, pe langa oxigen, asupra cresterii drojdiilor influenteaza si ceilalti componenti ai amestecului gazos. La inlocuirea treptata a azotului din aer cu CO2 (concentratia in oxigen ramanand constanta), se reduce cantitativ biomasa rezultata si continutul in azot al biomasei se mareste. Daca in experienta de control (pentru aerare s-a intrebuintat aer), biomasa de drojdie rezultata a fost de 0,502 g/1g zahar consumat si continutul in azot total in biomasa a fost de 8,8 % la s.u., atunci prin inlocuirea a 20 % azot din aer cu CO2, datele corespondente obtinute au fost 0,472 g/1g zahar consumat si 9,55 % azot total (Tuliakov, V.S. et al., 1980).
2.6.7. Aerarea si agitarea mediului
In procesul de biosinteza, aerarea urmareste asigurarea continua a celulei cu oxigen, eliminarea dioxidului de carbon format care are efect inhibitor asupra procesului de multiplicare, transportul rapid la celule a substantelor nutritive adaugate si mentinerea celulelor in stare de suspensie (Stoicescu, A., 1985).
Asigurarea necesitatii drojdiilor cu oxigenul din aer reprezinta o etapa de consum energetic mare in cadrul procesului tehnologic care se reflecta in final asupra avantajelor economice in productia de drojdie. S-a stabilit ca, o cauza importanta a scaderii de randament este insuficienta aerare a mediului de cultura (Dmitriev,D., 1983). La fabricarea drojdiei, consumul de aer este prevazut ca o norma de productie de consum, la volumul mediului cultural si este de 50÷100 m3/1m3 plamada.
Problema principala este asigurarea continua a celulei de drojdie cu oxigen dizolvat in lichid. S-a luat in considerare, ca la 1 kg de melasa cu un continut in zahar de 50 % sunt necesari aproximativ 19 m3 aer. Randamentul maxim in drojdie s-a obtinut, cand pentru fiecare gram de zahar folosit a corespuns 1,6 g (si mai mult) oxigen. La reducerea cantitatii de oxigen drojdia obtinuta scade proportional.
Alimentarea cu aer in vasul de cultivare trebuie sa se realizeze in concordanta cu alimentarea cu zahar si se urmareste viteza de multiplicare a drojdiei. Perturbarea regimului de aerare brusc, schimba mersul procesului de dezvoltare al drojdiei, in directia metabolismului anaerob, cu formare de alcool si alti produsi secundari. Producerea de biomasa scade brusc. In prezenta oxigenului in exces ritmul multiplicarii celulelor incepe sa scada, iar randamentul se reduce prin marirea consumului de zahar si formare de CO2.
Rolul oxigenului este diferentiat in functie de procedeele de multiplicare a drojdiilor. Daca prin metoda statica, la cultivarea drojdiilor fara aerare artificiala sau cu o slaba aerare, drojdia obtinuta a constituit 10÷12 % din greutatea materiilor prime consumate si durata multiplicarii drojdiilor a fost de aproximativ 20 h sau chiar mai mult, atunci prin metoda cu aerare-agitare, in mediu diluat, drojdia obtinuta a ajuns la 165 % calculata la greutatea zaharului sau pana la 100 % din greutatea melasei si ciclul complet de multiplicare a drojdiei a durat 8÷11 h. Consumul de aer a fost de 100 m3/h la 1 m3 plamada, iar cantitatea de oxigen utilizata de 6÷9 % din cantitatea totala. Consumul de oxigen la 1 g drojdie pentru celulele tinere a fost de 80÷100 mg/h, pentru cele mai in varsta 40÷60 mg/h. Necesitatea marita a celulelor tinere in oxigen este determinata de formarea substantelor fara azot din celulele de drojdie.
S-a stabilit ca pentru fiecare concentratie de plamada exista un optim al aerului adaugat. Surplusul de oxigen din mediu nu mareste cantitatea de biomasa obtinuta, insa intensifica procesele de oxidare, mareste potentialul de oxidoreducere.
La adaugarea in mediul sintetic de reducatori (0,02% acid tioglicolic sau 0,1% Na2S2O3) se mareste viteza de multiplicare. In consecinta, pentru o aerare oarecare, este necesara atat prezenta oxigenului cat si prezenta substantelor reducatoare, care scad potentialul oxidoreducator al mediului. Este evident ca, acest lucru este in legatura cu activitatea enzimelor. Se stie ca, de exemplu, reducatorii (Na2S si altii) maresc activitatea coenzimei A. Celulele de drojdie, crescute in conditii aerobe, spre deosebire de cele crescute anaerob, nu numai ca sunt mai putin bogate in glicogen, metacromatine si compusi azotati, dar au, de asemenea, masa mai redusa. Daca 1 miliard de celule crescute in conditii anaerobe, cantaresc 70÷90 mg, in conditii de aerobioza cantaresc 20 sau 25 mg, cel mult 50 mg. In consecinta, celulele mentinute in conditii de fermentare au masa de 2÷3 ori mai mare decat celulele in conditii de respiratie.
Cantitatea insuficienta de aer pentru inmultire, conduce la marirea numarului de celule mici in partea a doua a procesului de cultivare.
La cultivare pe agitator rotativ cresterea vitezei de aerare poate fi marita prin agitare. Maxon si Johnson au determinat eficienta aerarii, exprimata in milimoli de oxigen, raportat la litru si ora, in functie de debitul de aer si de agitarea mediului, la variatii ale turatiei agitatorului intre 550 si 1660 rot/min.
Unele cercetari recente recomanda recircularea aerului la fabricarea drojdiei de panificatie, deoarece creste randamentul printr-o mai buna folosire a substantelor nutritive ale mediului si da o intensificare a proceselor de multiplicare a celulelor de drojdie. In afara de aceasta, recircularea aerului imbunatateste un indice foarte important pentru activitatea drojdiei, activitatea maltazica. Eficienta recircularii aerului s-a exprimat prin cresterea procentului de celule inmugurite, numarate la un interval de doua ore pentru fiecare generatie (Dmitriev, A.D., 1982).
O importanta deosebita o reprezinta calitatea aerului, care poate fi o sursa de contaminare in fazele de multiplicare a drojdiilor. De aceea este necesara operatia de filtrare/sterilizare a aerului inainte de utilizarea lui in procesul tehnologic.
3. SEPARAREA SI SPALAREA BIOMASEI DE DROJDIE
Se realizeaza cu ajutorul separatoarelor centrifugale, de regula in doua sau trei trepte de separare, obtinandu-se in final un lapte de drojdie concentrat, care este apoi racit in schimbatoare de caldura cu placi, pana la temperatura de 2÷40C si pastrat in colectoare de depozitare.
La sfarsitul ultimei faze de multiplicare se obtine o plamada fermentata, in care celulele de drojdie se afla in suspensie, concentratia in drojdie a plamezii variaza in functie de calitatea melasei si de procedeul tehnologic folosit.
In cadrul procesului tehnologic clasic de fabricare a drojdiei se ajunge la o concentratie de 42÷50 g drojdie cu 27% s.u. la litru de plamada. Prin folosirea sistemelor dinamice de aerare concentratia plamezii in drojdie atinge valori de 4÷5 ori mai mari.
Prin separarea si spalarea laptelui de drojdie se urmareste concentrarea drojdiei din plamada intr-un volum mai mic si indepartarea resturilor de plamada in scopul imbunatatirii aspectului comercial si a conservabilitatii produsului.
Separarea drojdiei se efectueaza cu ajutorul separatoarelor centrifugale cu talere tip Alfa Laval sau Westfalia cu turatii de 4000÷5000 rot./minut si capacitati cuprinse intre 10 si 100 m3 plamada/ora.
In practica, operatia se realizeaza in doua sau trei trepte de separare si spalare, cea mai utilizata fiind separarea in trei trepte. In prima treapta de separare, in functie de concentratia initiala a plamezii, laptele de drojdie se concentreaza pana la 150÷200 g/l. Inainte de trecerea la treapta urmatoare de concentrare este necesara o racire si diluare cu apa, folosind in acest scop ejectoare. Cantitatea de apa necesara este de 4÷8 ori mai mare decat cea de lapte de drojdie.
In treapta a doua de separare se poate obtine o concentratie de 300÷400 g/l. Acest proces se repeta in treapta a treia, obtinandu-se in final un lapte de drojdie cu o concentratie de 600÷800 g/l.
Laptele de drojdie concentrat este racit in schimbatoare de caldura cu placi pana la temperatura de 2÷40C si pastrat in colectoare de depozitare. Prin racire procesele vitale din celula de drojdie sunt incetinite si este franata dezvoltarea si activitatea microorganismelor de contaminare.
Pregatirea separatoarelor si a instalatiei pentru un ciclu de separare consta in: demontarea acestora, spalarea cu peria a fiecarui taler cu o solutie de fosfat trisodic sau soda caustica, urmeaza clatirea cu apa curata pentru desfundarea duzelor. De asemenea, se spala si se clatesc cuvele pentru colectarea laptelui separat, precum si pompele si conductele aferente instalatiei.
Colectoarele pentru lapte de drojdie sunt confectionate din otel inoxidabil, prevazute cu manta dubla de racire, agentul frigorific fiind apa racita si cu agitatoare actionate electric in vederea omogenizarii.
In timpul depozitarii se controleaza la intervale de timp de 4 ore temperatura laptelui de drojdie care trebuie sa se mentina la 2÷40C.
4. Filtrarea laptelui de drojdie
Laptele de drojdie nu poate fi comercializat ca atare atat datorita faptului ca este usor expus la contaminarea cu microorganisme straine care ii micsoreaza conservabilitatea cat si datorita greutatii in manipulare. Din aceste motive laptele de drojdie este supus in continuare operatiei de filtrare si presare, prin care drojdia se concentreaza in substanta uscata ocupand un volum de circa doua ori mai redus. Aceasta operatie tehnologica se realizeaza in practica cu ajutorul filtrelor-presa (cu rame si placi) si a filtrelor rotative sub vid.
Procesul de filtrare cu ajutorul filtrelor presa se desfasoara astfel:
inainte de utilizare, filtrul-presa este spalat cu apa, montat si sterilizat cu abur timp de 15÷30 minute fara panza;
se strang in pachet compact ramele acoperite cu panza si placile fiecarei prese folosind compresorul de ulei;
laptele de drojdie racit, este introdus cu ajutorul unei pompe printr-un canal central in spatiul pe care il formeaza ramele marginite de panze filtrante, drojdia ramane in spatiul pe care il formeaza rama, iar apa trece prin panza si se scurge la canal;
filtrarea dureaza 15÷30 minute, pana cand nu se mai observa evacuarea apei;
la sfarsitul operatiei se desfac treptat ramele si placile, se detaseaza cu ajutorul unor cutite drojdia comprimata, care se colecteaza intr-un carucior;
la intervale de timp de circa o saptamana panzele colmatate se spala mai intai cu jet de apa si cu peria si apoi cu ajutorul unei masini de spalat panze si se trec in uscatorul de panze.
Drojdia comprimata rezultata la sfarsitul presarii are un continut ridicat in substanta uscata de 30÷35%, insa folosirea filtrelor-presa prezinta dezavantajul unei productivitati scazute si a unui volum mare de munca.
Fabricile moderne de drojdie utilizeaza filtre rotative sub vid, care folosesc drept strat filtrant amidonul. Cu acest filtru se imbunatateste considerabil substanta uscata a produsului finit (de la 27% s.u. la obtinerea drojdiei cu filtre-presa s-a ajuns la 33÷37% s.u.). Prin inglobarea de clorura de sodiu in laptele de drojdie si spalarea acestuia, s-a putut mari continutul de substanta uscata al drojdiei la peste 30%, prin eliminarea apei extracelulare, cu marirea concomitenta a plasticitatii.
5. Modelarea si ambalarea drojdiei presate
Modelarea si ambalarea drojdiei presate se realizeaza in prezent cu masini automate de constructie speciala, utilizand hartie parafinata sau sulfurizata cu film de celofan. Pentru a se obtine consistenta necesara modelarii este necesar sa se adauge o anumita cantitate de apa, ulei comestibil sau alti plastifianti. Pentru pastrarea culorii drojdiei se mai pot adauga cantitati mici de polialcooli (de exemplu, glicerina, inozitol) sau substante emulsionante (lecitina, stearati si oleanati ai glicerinei si glicolului), iar pentru protectia impotriva dezvoltarii mucegaiurilor se pot adauga cantitati mici de alcool etilic, propilic, butilic sau amilic.
6. DEPOZITAREA SI LIVRAREA DROJDIEI DE PANIFICATIE
Atunci cand livrarea drojdiei nu se realizeaza imediat, lazile sau cutiile de carton cu drojdie trebuie depozitate intr-o incapere racita la temperatura de 0÷40C si umezeala relativa a aerului de 65÷70%. Lazile sau cutiile de carton sunt asezate pe stelaje sau paleti in forma de fagure, cu locuri pentru circulatia aerului.
Transportul drojdiei la beneficiari se poate face cu mijloace de transport obisnuite pe distante mici, iar pe distante mai mari in vagoane sau mijloace auto izoterme. Livrarea se efectueaza pe sarje, in ordinea fabricarii, prin reluarea lazilor sau cutiilor de carton de pe palet, pe banda si evacuate la rampa pentru incarcarea mijloacelor de transport.
7. Drojdia de panificatie - produs finit
Cunoasterea compozitiei chimice a drojdiei de panificatie este importanta pentru stabilirea cantitatilor de substante nutritive necesare pentru multiplicarea drojdiei in diferite faze cat si modul lor de adaugare, in vederea obtinerii de randamente maxime in drojdie si pentru intelegerea proceselor care au loc in timpul pastrarii drojdiei in calup.
Compozitia drojdiei de panificatie comprimata este prezentata in tabelele 24; 25; 26; 27.
Tabelul 24
|
Azot, % s.u. | |
|
Protide, % s.u. | |
|
Glucide, % s.u. | |
|
Lipide, % s.u. | |
|
Cenusa, % s.u. | |
|
P2O5, % s.u. | |
|
Apa, % |
Se apreciaza ca, aproximativ 94% din substanta uscata a drojdiei este alcatuita din principalele elemente: carbon, hidrogen, oxigen si azot, care sunt reprezentate de glucide (glicogen, gume, hemiceluloze), proteine, acizi nucleici, baze organice, lipide, substante minerale, vitamine si enzime. Continutul in carbon al unei drojdii cu 27% s.u. este de aproximativ 12,7% si serveste ca baza pentru calculul necesarului de glucide pentru acumularea biomasei de drojdie.
Tabelul 25
Continutul in aminoacizi al proteinelor drojdiilor (% din greutatea proteinelor)
|
Cistina, triptofan, metionina | |
|
Histidina, alanina | |
|
Glicocol, prolina, tirozina, arginina, izoleucina | |
|
Serina, treonina, valina | |
|
Alanina, leucina | |
|
Lizina, asparagina | |
|
glutamina |
Aproximativ 70% din azotul total al drojdiei este inclus in proteine, 8÷10% in baze purinice, 4% in pirimidine, restul fiind format din produse solubile ca aminoacizi si nucleotide. Plecand de la continutul in azot al drojdiei se stabileste necesarul de substante cu azot pentru corectarea melasei care este deficitara in azot.
Drojdia contine si cantitati importante de vitamine, in special din grupul B.
Tabelul 26
Continutul de vitamine al drojdiei de panificatie (μg/%s.u.)
|
Tiamina (B1) | |
|
Riboflavina (B2) | |
|
Piridoxina (B6) | |
|
Acid nicotinic (PP) | |
|
Biotina | |
|
Acid pantotenic | |
|
Acid p-aminobenzoic | |
|
Mezoinozitol | |
|
Acid pteroilglutamic |
Substantele minerale se gasesc fie in combinatii anorganice sau intra in compozitia unor substante organice, aflandu-se deci ca electroliti in solutie sau sub forma de complexe coloidale.
Tabelul 27
Compozitia minerala a drojdiei de panificatie (mg/100 g)
|
Potasiu | |
|
Sodiu | |
|
Calciu | |
|
Magneziu | |
|
Fosfor |
Valoarea energetica : 350÷430 KJ/100 g.
Biomasa unui gram de drojdie comprimata contine aproximativ 10 miliarde de celule.
In cursul procesului de fabricare a drojdiei de panificatie, concomitent cu multiplicarea celulelor apartinand culturii pure, in diferite faze ale fluxului tehnologic se pot dezvolta si alte microorganisme, care maresc gradul de contaminare a produsului finit si determina reducerea calitatilor tehnologice si conservabilitatea drojdiei comprimate.
Pentru a preveni multiplicarea microorganismelor contaminante, se impune un control microbiologic riguros pe faze de productie, prin studiul gradului de igiena si detectarea contaminantilor ce pot proveni din sursele prezentate in figura 24.
Din punct de vedere microbiologic, drojdia comprimata de buna calitate are caracteristicile din tabelul 2
Tabelul 28
|
Staphylococcus aureus |
< 10/g |
|
Salmonella |
Lipsa in 25 g |
|
Bacterii lactice |
<103 |
|
Bacterii coliforme |
<102 |
(cuibul) (melasa) Materiale auxiliare Microbiota aerului si apei Igiena materialelor pentru ambalat Igiena individuala a personalului Spalarea si dezinfectarea
necorespunzatoare a utilajelor si conductelor produs finit Igiena
salilor de productie
Cultura
de drojdie
Materia
prima
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
DROJDIE
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Fig.24. Surse de contaminare la producerea industriala a drojdiei de panificatie
Drojdia de panificatie - produs finit trebuie sa prezinte urmatoarele caracteristici biotehnologice:
putere de fermentare - max. 70 minute;
umiditate - max. 76%;
durabilitate la 350C - min. 5 zile;
durabilitate la 0÷40C - min. 10 zile.
Copyright © 2024 - Toate drepturile rezervate