|







 |  |
|
| |
 | Alpinism | Arta cultura | Diverse | Divertisment | Film | Fotografie |
| Muzica | Pescuit | Sport |
FACTORII DE MEDIU CARACTERISTICI MARII NEGRE CU INFLUENTA ASUPRA RESURSELOR PESCARESTI, UNELTELOR SI CONDITIILOR DE PESCUIT
1. Istoric
Numele de Neagra al marii care scalda litoralul Romaniei are o provenienta care se pierde in negura istoriei popoarelor care au stapanit-o si au locuit in jurul tarmurilor ei (Leonov A., K., 1960). Istoricul latin Strabon aminteste ca grecii antici numeau bazinul Marii Negre Axenos (inospitaliera), din cauza frecventelor furtuni care bantuiau aceasta mare. Mai tarziu, cand grecii au cunoscut mai bine Marea Neagra si au fondat coloniile de-a lungul coastelor sale, au denumit-o Euxinus (ospitaliera), iar romanii (secolul I i.e.n.), au compus cuvintele grecesti Pontos (mare completa) cu Euxinus, dand numele de Pontus Euxinus (mare ospitaliera).
In antichitate Marea Neagra s-a mai numit si Marea Scytica, Cimerica, Amasonica, Sarmatica, Pontica (Seiseanu R., 1956). Grecii bizantini au fost primii care au denumit-o Mare ajore iar venetienii au numit-o Marea Maura.
Istoricul roman Hajdeu pretinde ca numele de Negreata Pontului s-a ivit abia in evul mediu, gasindu-se pentru prima data la tatari, slavi, scandinavi, unguri si la turci, acele popoare care cutreierau pe atunci partea de nord-vest a marii (Seiseanu R., 1956). Hajdeu mai sustine ca originea numelui de Neagra se poate duce cel mult pana in secolul al X-lea si ca ea se datoreste romanilor. Domnitorul Dimitrie Cantemir o denumeste in lucrarea sa Tabula Geographica Moldavie (1710), Mare Nigrum.
Marea Neagra a inceput sa fie studiata inaintea altor mari. In secolul al V-lea i.e.n. istoricul grec Herodot a vizitat tarmurile nordice si a descris natura si clima, denumind cateva rauri care se varsau in mare in aceste zone. Pe masura dezvoltarii schimburilor si comertului pe apa, au crescut tot mai mult nevoile navigatiei. Primele indreptare de navigatie s-au intocmit in secolul IV-lea i.e.n., purtand numele de Peripl, ceea ce corespunde intru totul cu modernele carti pilot. In secolul al III-lea i.e.n. a fost intocmita prima harta a Marii Negre.
2. Caracterizarea bazinului Marii Negre
2.1. Delimitarea geografica si pescareasca a Marii Negre
Prin pozitia sa in interiorul uscatului, Marea Neagra este o mare de tip intercontinental, oarecum izolata, indepartata de ocean si care separa Europa sud-estica de Asia Mica si tarmurile Caucazului, fiind legata cu apele Marii Marmare la sud-vest prin stramtoarea Bosfor, iar la nord-est comunica cu Marea Azov prin stramtoarea Kerci, a carui adancime de circa 12 m este intretinuta artificial (Fig. 1).
|
Fig. 1. Delimitarea geografica |
Limitele bazinului actual se inscriu intre 40 latitudine nordica si 27 longitudine estica, avand o forma alungita usor ingustata la mijloc. Suprafata bazinului reprezinta o arie 42488 km2, cuprinzand un volum de apa de 537.000 km3 de apa. Bazinul este orientat est-vest, fiind o depresiune intermontana, marginita de doua centuri de cute alpine (Degens & Ross si al., 1974). Adancimea maxima a bazinului Marii Negre este de 2.212 m, cea medie de 1.271 m.
Linia totala a tarmului Marii Negre este de circa 4.340 km lungime, din care: tarmul bulgaresc este de 300 km lungime; cel georgian de 310 km; cel romanesc de 244 km; cel rusesc de 475 km; cel turcesc de 1.400 km iar cel ucrainian de 1.628 km. Catre sudul Marii Negre sunt muntii Pontici cu inaltimea maxima de 937 m (varful Kachkar) si in est, muntii Caucaz cu inaltimea maxima de 5.642 m (varful Elbrus). Tinuturile marginale nordice si vestice reprezinta terenuri relativ joase, cea ce mai accentuata forma de relief fiind a peninsulei Crimeii cu o inaltime maxima de 1.545 m, cu peninsula Kerci legata de ea.
Principala particularitate a Marii Negre este stratificarea verticala. Marea diferenta intre densitatea apei din stratul superior de 100 -150 m si a celui de fund, si implicit marea diferenta intre regimul termic, salin, cel al gazelor dizolvate, al nutrientilor si al structurii biotice a celor doua straturi au definit Marea Neagra ca un bazin bioanizotrop, sau bazin tipic cu anomalie caracteristica.
A doua particularitate, intalnita numai in cateva sectoare ale oceanului mondial, este cantitatea enorma de hidrogen sulfurat, 85-90% din volumul apelor acesteia si aproximativ 65% din suprafata sa fiind lipsite de viata datorita prezentei hidrogenului sulfurat. Astfel spus, Marea Neagra este compusa din doua paturi de apa suprapuse, cu caracteristici net diferite, intre care schimbul de ape prin curenti verticali este practic inexistent: una la suprafata (0-150 m adancime), patura de apa cu salinitate mai redusa, cu schimbari sezoniere ale regimului termic si salin, cu oxigen in zona oxidarii, cu un dinamism remarcabil, zona cu conditii favorabile existentei vietii si cea de a doua patura de apa, de fund (sub 150-200 m), cu o salinitate mai ridicata, cu o stabilitate termica si salinitate pronuntata, cu un dinamism redus practic la zero, lipsita de oxigen dar bogata in hidrogen sulfurat, zona lipsita de viata (cu exceptia bacteriilor).
Gradul inalt de izolare fata de Oceanul mondial, bazinul de receptie extins (peste 2 milioane km2) si numarul mare de rauri care se varsa in mare, contribuie la balanta unica a Marii Negre. Balanta hidrografica este un factor important in ecosistemul marii. Schimbarile in regimul hidrografic au un impact semnificativ in balanta salinitatii si apei, care sunt cele mai vulnerabile la orice schimbari antropogene, in special in zona de nord vest cu o productivitate biologica ridicata.
O trasatura specifica Marii Negre ce o deosebeste de majoritatea marilor continentale, este faptul ca minimum termic nu se gaseste pe fundul bazinului ci in stratul 50-100 m. Aceasta situatie se explica prin existenta diferentelor mari de salinitate dintre paturile superficiale si masele de apa profunde ce un permit decat o circulatie verticala foarte redusa. Stratificatia salina transversala pe linia de tarm este puternic influentata de circulatia longitudinala a masei de apa si indeosebi de curentii formati sub actiunea vanturilor.
Configuratia reliefului marin se compune din patru elemente fiziografice: platforma continentala 29,9%, panta continentala 27,3%, soclu continental 30,6% si platforma abisala 12,2%.
Dezvoltarea maxima a platformei continentale este la vest de peninsula Crimeea, unde latimea ei poate depasi 90 km. De-a lungul coastelor Anatoliei, Caucazului si la sud de Crimeea, latimea platformei continentale nu depaseste decat rareori 20 km, iar in dreptul coastelor bulgaresti si turcesti latimea medie a platformei este de 40 km.
Tarmul romanesc al Marii Negre poate fi impartit in doua sectoare geografice si geomorfologice diferite:
sectorul nordic (circa 166 km) care se intinde intre Baia Musura si Cap Midia, aflat in imediata apropiere a Deltei Dunarii si a complexului Lagunar Razelm-Sinoe, avand sedimente de origine aluvionara;
sectorul nordic (circa 78 km) care se intinde intre Cap Midia si Vama Veche, unde predomina falezele inalte, intrerupte doar in locurile cu mici plaje inguste.
Datorita faptului ca platforma continentala, cu o adancime de pana la 200 m, are o intindere mare (22.998 km2) in dreptul litoralului romanesc, biodiversitatea este foarte ridicata, intalnindu-se aici si cel mai mare numar de specii endemice relicte.
Gestionarea si supravegherea resurselor vii din zona FAO 37, respectiv 37.4.2 (Marea Neagra)(Fig. 2) este sub directa indrumare a General Fisheries for the Mediterranean (GFCM), organism international infiintat la data de 24 septembrie 1949, in baza articolului XIV al Regulamentului FAO si a intrat in vigoare la data de 20 februarie 1950, suferind pana in prezent doua amendamente (1963 si 1976). La nivelul anului 1995, la acest organism erau afiliate un numar de 20 de state, majoritatea tari riverane, printre care si Romania.
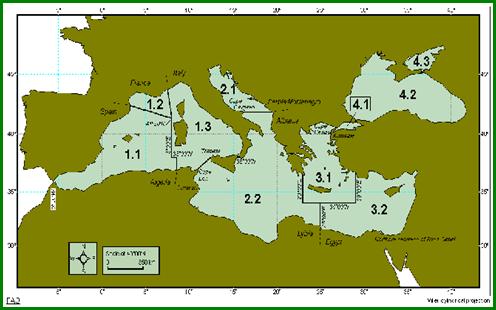
Fig. 2. Delimitarea pescareasca
2.2. Descrierea habitatului
Fundul Marii Negre este impartit in self, panta continentala si depresiunea marina adanca. Partea nord-vestica a Marii Negre, este o platforma larga cu portiuni inguste catre sud, care ajung pana la Bosfor. Izobata de 200 m, limita platoului continental, este mai aproape de tarmurile nord-estice, estice si sudice ale Marii Negre, si la 20 Mm de cele vestice si aproape la 120 Mm la cele nord-vestice. Panta continentala este foarte rapida in sud, unde de la 200 la 1000 m se trece in mai putin de 10 Mm. Centrul Marii Negre este ocupat de un bazin ale carui adancimi ating 2.244 m.
Selful sau platforma continentala, continuarea directa a uscatului, are o latime maxima in partea nord-vestica, unde izobata de 200 m se indeparteaza de tarm pana la 180-200 Km. In partea de vest platforma se ingusteaza intr-o fasie lata de 50 km, pentru ca in partea estica si sudica aceasta sa atinga numai cativa kilometri. Dupa platforma continentala adancimile cresc brusc, atingand pante de 0,11 in dreptul costei sudice a Crimeei, 0,25 pe coasta Caucazului si chiar 1,19 in dreptul regiunii Novorosiisk. Sub izobata de 800 m, taluzul fundului submarin este cu mult mai mic, avand
inclinari de ordinul minutelor.
Din suprafata totala a marii de 41490 Km2, platforma continentala (adancimi sub 200 m) ocupa circa 35% (13000 km2).
Tarmul de vest al peninsulei Crimeea este abrupt si aproape pe intreaga intindere este alcatuit din argile rosii. Pe masura apropierii de Sevastopol argilele rosii trec in argile galbui. Spre capul Alia, tarmul devine foarte ridicat si chiar stancos, iar dupa capul Saraci, devine plat.
Partea sud-estica a marii, tarmul este alcatuit in majoritate din sisturi argilo-calcaroase cenusii deschise, din calcare cenusii si conglomerate ale jurasicului superior. Tarmul sudic al marii este in intregime muntos pana la Bosfor, cu faleze inalte, constituite din argile, marme, gresii, calcar iar partea nord -vestica este format din cordoane nisipoase joase, separate de numeroase balti si limane. Sedimentele care acopera fundul marin se impart in doua mari grupe:
A sedimente de adancimi mici (pana la 180-200 m) cuprinse in patura superficiala de mase de apa situate deasupra platformei continentale;
A sedimentele de adancimi mari (peste 200 m) cuprinse in paturile inferioare de apa neoxigenata si bogate in hidrogen sulfurat.
In sedimentele de adancimi mici se incadreaza sedimentele litorale ce contin blocuri, bolovani, pietrisuri si nisipuri ce se intind pana la adancimi de 40-50 m si sedimente maloase, care pot fi maluri cu midii (intre 30-65 m) si maluri cu Modiola phaseolina (adancimi de 65-200 m). Sedimentele nisipoase sunt constituite din cuart, muscovit si mai rar din silmenit si stavrolit. Substanta organica continuta de nisipurile litorale este neinsemnata. Malurile litorale contin in general cantitati insemnate de argile de culoare cenusie intunecata, avand incluse in ele scoici de diferite dimensiuni.
Sedimentele de adancimi mari sunt alcatuite din argile cenusii de mare adancime, maluri argilo-calcaroase, calcaroase si nisipoase.
In general, tarmurile sunt acoperite de bolovanisuri, pietrisuri si nisipuri, cele inspre larg trec in nisipuri curate si maluri nisipoase. Intre izobatele de 200-1.500 m, se intalneste malul negru, iar peste 1.500 m mal calcaros cenusiu deschis compus din carbonat de calciu. Cele mai mari adancimi ale marii sunt acoperite cu argila omogena si nestratificata in care se afla foarte multa substanta organica.
Fig. Natura sedimentelor
din Marea Neagra 1 - nisip; 2 - mal; 3 -
argila cenusie de adancime; 4 - mal argilos; 5 - mal intermediar;
6 - mal intermediar cu stratif de argila cenusie si nisip; 7
- mal calcaros; 8 - zone cu abundenta de sedimente recente; 9 -
scoicarisuri; 10 - mal faseolinic; 11 - argila cenusie;
12 - argila cenusie cu statificari, 13 mal cu argila
cenusie; 14 - mal calcaros; 15 - calcare din scoici pe mal nisipos
si nisip malos
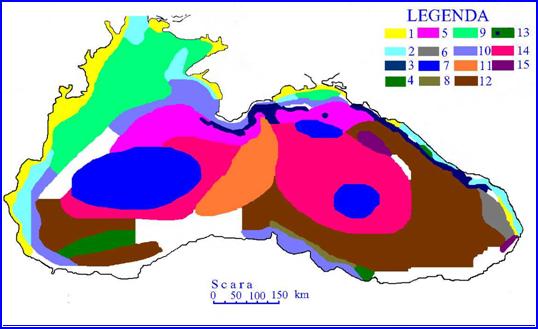
Cea mai mare parte a fundului Marii Negre este acoperita de maluri. Astfel, la nord de golful Burgas, fundul platformei continentale este format din mal, nisip si pietris. Aceeasi configuratie o intalnim si intre Burgas si Baia Zembrisnskaia, unde banda costiera este foarte ingusta. In sectorul baia Zembrinskaia pana la Odesa, fundurile din apropierea tarmului sunt nisipoase sau pietroase iar in zona de larg un amestec de mal, nisip si pietris. Intre Odesa si Ins. Tendra zonele nisipoase si pietrisurile alterneaza cu zonele de mal nisip si pietris. La sud de peninsula Crimea si pana la baia Feodosia fundul platoului continental este acoperit de mal si nisip. In continuare pana la granita turca, fundul costier este format fie din placi largi de mal depus direct pe roca, fie din mal, nisip sau pietris. In aria platoul continental turcesc se intalnesc adesea mal in amestec cu cateva placi nisipoase (Fig. 3).
2. Caracterizarea conditiilor climatice ale zonei
Marea Neagra, prin pozitia ei, se gaseste intr-o regiune in care presiunea medie anuala de la nivelul marii este aproximativ 1016-1018mb. Valorile extreme ale presiunii medii lunare sunt atinse in lunile iulie-august (1010-1014mb) si in luna ianuarie (1018-1025mb). Mediile multianuale si valorile extreme ale presiuni atmosferice de la nivelul marii, inregistrate la statiile meteorologice Sulina, Constanta si Mangalia, scot in evidenta unele particularitati importante (Tabel 1). Astfel, faptul ca media multianuala a minimelor absolute a valorilor presiunii in statiile Constanta (987,5 mb) si Sulina (989,8 mb) sunt mai mici decat cele de la Mangalia (992,9 mb) presupune ca traiectoria campului baric minim ar fi plasata intre Sulina si Constanta (mai aproape de Constanta). Mediile multianuale sunt foarte apropiate intre ele, fapt ce confirma ca litoralul romanesc se gaseste in general sub influenta acelorasi procese macrosinoptice. Mediile maximale scad de la Sulina (1038,2) la Mangalia (1034,7 mb, scadere determinata direct de influenta pe care o exercita dorsala anticiclonului siberian asupra proceselor din aceasta zona.
Tabel 1
Valori medii multianuale ale presiunii atmosferice (mb),
inregistrate in sectorul litoralului romanesc al Marii Negre
Statiameteorologica |
Valori caracteristice (mb) |
|||
|
Medie minimelor |
Media multianuala |
Media maximelor |
Amplitudinea absoluta |
|
|
Sulina | ||||
|
| ||||
|
Mangalia | ||||
Datorita legaturii stanse intre distributia spatiala a presiunii si deplasarea maselor de aer, regimul vanturilor din regiunea Marii Negre prezinta urmatoarele caracteristici:
in perioada rece, datorita zonei depresionare de pe mare si influenta dorsalei anticiclonului siberian, Marea Neagra se afla sub actiunea vanturilor din sectorul nordic (NE, N si NV), care sunt foarte puternice de-a lungul coastei si mai slabe spre larg. Incepand cu luna aprilie, odata cu restructurarea campului baric determinata de schimbarile survenite in circulatia generala a atmosferei, se schimba si directia vantului;
in perioada calda, din cauza predominarii maximului azoric, vanturilor in partea de vest a Marii Negre sufla din directia NV, V si SV iar in rest vanturile sufla dinspre larg spre coasta.
Analiza datelor prelevate de catre INCDM Grigore Antipa Constanta, in perioada 1990-2000, arata ca frecventa vantului din sectorul nordic (NV, N si NE) reprezinta circa 44% din totalul cazurilor. Vanturile din sectorul nordic au si vitezele medii cele mai mari fata de celelalte sectoare. Astfel, vitezele medii ale vanturilor din sectorul nordic sunt cuprinse intre 3,4 m/s (Mangalia) si 4,9 m/s (Sulina), in timp ce vitezele medii pe celelalte sectoare sunt cuprinse intre 2,7 m/s (Mangalia) si 3,4 m/s (Constanta).
Pentru Marea Neagra si zonele inconjuratoare, factorul care genereaza o distributie spatiala diferentiata a temperaturii aerului este suprafata adiacenta. Temperatura aerului din zona litoralului Marii Negre corespunde unui climat de tip mediteranian, caracterizat prin ierni calde si umede si veri foarte calde si sarace in precipitatii, exceptie facand regiunea de sud-est a marii, unde precipitatiile sunt abundente, iar clima este mai mult de tip tropical. In zona de larg izotermele medii anuale sunt aproape paralele cu coasta.
Temperatura
medie anuala creste de la nord-vest (unde este de 10°C) spre sud-est
(unde atinge 15,5°C). Temperatura medie lunara este cuprinsa intre
-0,6°C (Sulina) si +0,2°C (Mangalia), in luna ianuarie si intre
22,4°C (Sulina) si 24,8°C (Mangalia), in luna iulie. Media anuala
este de 11,1°C (Sulina) si 11,2°C (
Valorile medii multianualeale temperaturii aerului (°C),
la litoralul romanesc al Marii Negre(1896 - 2000)
|
Statia |
V a l o r i l e l u n a r e |
Val. anuala |
Media 12 luni |
Ampli. lunara |
||||||||||||
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
XI |
XII |
|||||
|
Media |
Sulina | |||||||||||||||
|
C-ta | ||||||||||||||||
|
Mang. | ||||||||||||||||
|
Media Max. |
Sulina | |||||||||||||||
|
C-ta | ||||||||||||||||
|
Media Min. |
Sulina |
| ||||||||||||||
|
C-ta | ||||||||||||||||
|
Max. absol |
Sulina | |||||||||||||||
|
C-ta | ||||||||||||||||
|
Mim. absol |
Sulina | |||||||||||||||
|
C-ta | ||||||||||||||||
|
Media max. zilnica |
Sulina | |||||||||||||||
|
C-ta |
2.5 | |||||||||||||||
|
Mang. | ||||||||||||||||
|
Media mim. zilnica |
Sulina | |||||||||||||||
|
C-ta | ||||||||||||||||
|
Mang. | ||||||||||||||||
2.4. Caracterizarea hidrologica a regiunii
Dat fiind schimbul de apa din Marea Neagra are loc prin stramtorile Bosfor si Kerci, foarte slab in comparatie cu volumul de apa al cuvetei, regimul general al curentilor prezinta caracteristicile curentilor din bazinele oceanice izolate.
In sezonul rece al anului, pe intreg bazin al Marii Negre se formeaza un sistem ciclonic de curenti cu doua centre: unul in jumatatea vestica si altul in jumatatea estica. In extremitatea nord-vestica si sud-estica se formeaza doua anticicloane secundare. In perioada de tranzitie dintre sezonul rece si cel cald situatia este inversa fata de sezonul rece, insa ceva mai slab dezvoltata.
In perioada calda a anului, in jumatatea vestica se formeaza un centru ciclonic iar in jumatatea estica un anticiclon. In extremitatea nord-vestica se formeaza un centru secundar anticiclonic, iar in cea sud-estica un centru secundar anticiclonic (Fig. 4)
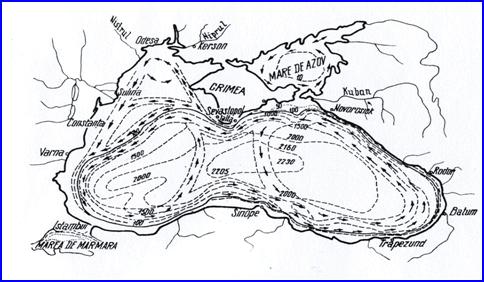 Fig. 4. Curentii ciclonici din Marea Neagra
Fig. 4. Curentii ciclonici din Marea Neagra
In partea nord-vestica a Marii Negre, de-a lungul litoralului romanesc in circulatia apelor se face simtita ramura curentului ciclonal al bazinului, cunoscuta sub numele de curentul Rameliei. Directia sa generala de curgere este de la nord spre sud, reprezentand rezultatul actiunii unui cumul de cauze: concentrarea surselor de apa dulce din sectorul nord-vestic al marii; evacuarea excedentului de apa prin Bosfor; dominanta vanturilor din sectorul nordic.
Datorita instabilitatii vanturilor, atat ca directie cat si ca intensitate, rareori curentii ating regim stationar, ei prezentand un mare grad de variabilitate. Faptul ca frecventa vanturilor din sectorul nordic este mai mare decat a celor din sud, face ca rezultanta generala a circulatiei superficiale sa fie dirijata spre sud.
Avand in vedere frecventele insemnate ale componentelor nordice si estice ale curentilor de suprafata, se poate afirma ca cel putin sase luni pe an la litoralul romanesc al Marii Negre au existat si exista conditii de formare a curentilor de upwelling. Durata si intensitatea acestora depende de marimea perioadelor de a curentilor favorabili, intalnite in special in lunile februarie, aprilie, iunie si iulie (Rapoarte INCDM, 1998-2006).
Dimensiunile relativ mari ale Marii Negre, adancimile mari din apropierea coastelor, micile cutari ale liniei tarmului, slaba si scurta darata de dezvoltare a gheturilor, aparitia frecventa a cicloanelor cu intensitate maxima in perioada de iarna, precum si predominarea vanturilor din sectorul nordic si nord-estic, creaza conditii prielnice pentru dezvoltarea valurilor de vant.
Intensitatea medie lunara a agitatiei valurilor oscileaza sincron cu mersul anual al intensitatii vantului de la 2,8 grade in luna august la 4-5 grade in lunile ianuarie-februarie (Bondar si al. - 1972)
De-a lungul litoralului romanesc, in cea mai mare parte a anului, marea este supusa actiunilor vanturilor producatoare de valuri. Din totalitatea valurilor marii care apar in zona de coasta a litoralului romanesc, circa 76% sunt valuri de vant si numai 24% valuri de amortizare (hula).
Regimul valurilor la litoralul romanesc este determinat nu numai de regimul vanturilor ci si de structura batimetrica a zonei si de configuratia si orientarea liniei de coasta. Datorita variabilitatii considerabile a regimului vanturilor, caracteristicile campurilor de valuri existente in zona litoralului romanesc se modifica in mod semnificativ in decursul unui an si prezinta mari diferente interanuale.
Pe termen lung, durata perioadelor de calm (inaltimea valurilor mai mica de 0,2 m) este maxima in luna mai (66,5%, din perioada) si minima in luna februarie (52,1%), iar ponderea situatiilor cu valuri de vant intre 20,3% (mai) si 39,4% (octombrie), iar a celor de hula intre 13,1% (iunie) si 20,6% (noiembrie) (Rapoarte INCDM, 1998-2006). Pe perioada unui an, cea mai mare pondere o au tot valurile de vant din directia nord-est (25,2%) si a hulelor de est (12%).
Analiza curbelor de frecventa pentru parametri caracteristici campului valurilor, releva faptul ca 88,8%, din totalul valurilor masurate intr-un interval de 23 de ani, au inaltimi de 0,2 m si 1,6 m, iar 82,5% au lungimi de 1,0-4,1 m (Rapoarte INCDM, 1998-2006). Inaltimea maxima observata a valurilor de la litoralul romanesc a fost de 5,0 m, inregistrat in luna decembrie 1997 si de 9,5 m in larg de platforma petroliera Gloria (Raport IRCM, 1997).
Din punct de vedere termic Marea Neagra prezinta particularitatile marilor semiinchise situate in zona temperata si a caror caracteristica esentiala o constituie diferentele foarte mari de valori ce se inregistreaza in stratul superficial al apei, de la sezonul rece la cel cald. O trasatura specifica Marii Negre, ce o deosebeste de majoritatea marilor si oceanelor globului este faptul ca minimum termic nu se gaseste pe fundul bazinului ci in stratul de 50-100 m. Aceasta situatie se explica prin existenta diferentelor mari de salinitate dintre paturile superficiale si masele de apa profunde ce nu permit decat o circulatie verticala foarte redusa.
In general, distributia pe verticala a temperaturii apei este strans legata de regimul termic al atmosferei si de factorii dinamici ai marii (curenti si valuri), care produc amestecul maselor de apa. Cum regimul termic al atmosferei este relativ constant, singurii factori care exercita o influenta determinanta asupra proceselor de incalzire pe verticala a maselor de apa sunt curenti si valurile. Din acest motiv, patura superficiala a maselor de apa este supusa unor oscilatii termice importante pana la adancimea la care ajunge influenta de amestec a apei provocata de valuri si curenti. Amestecul intens de apa atinge in general adancimea de 100-150 m si foarte rar 200 m. Masele de apa intermediare si de mare adancime (care cuprind circa 88% din volumul cuvetei marii), desi se gaseste intr-o stare de schimb continuu, dar lent, cu paturile superioare, sunt supuse unor variatii termice foarte mici, practic neglijabile (Bondar si al. - 1972).
Caracterist pentru masa de apa din bazinul Marii Negre este faptul ca sub adancimea de 200 m temperatura apei ramane practic constanta (8-9°C), cu o usoara tendinta de crestere spre straturile mai profunde.
Apele Marii Negre din zona litoralului romanesc sunt caracterizate la suprafata printr-o temperatura medie anuala de 12.6°C, langa coasta si de 13,0°C in zona de larg. Temperatura maxima a apei marii atinge uneori valoarea de 27,0°C (luna iulie). Temparatura minima nu scade sub valoarea punctului de inghet (circa -1,3°C, pentru o salinitate de 18 )(Rapoartele INCDM, 1998-2006).
Pe sezoane exista urmatoarele caracteristici ale distributiei verticale a temperaturii apei: in sezonul rece al anului (decembrie-februarie) temperatura apei creste odata cu adancimea, ele fiind mai pronuntate in perioada cea mai friguroasa, cand temperatura apei de suprafata atinge valorile cele mai mici (0,5-2,0°C), interval in care temperatura la adancimea de 50 m este de 6-7,5°C; incepand cu luna aprilie, temperatura apei marine creste continuu, pana in luna augut, cand atinge maximul, dupa care, ca urmare a unui bilant caloric negativ, incepe sa scada.
In perioada de incalzire si racire a atmosferei, stratul activ de apa al marii (pana la 70 m) se incalzeste si se raceste corespunzator, de la suprafata spre fund si de la mal spre larg, ceea ce inseamna ca, in perioada de incalzire, masa de apa din zona de larg este mai rece decat apa costiera, si invers in perioada rece, mai calda in zona de larg si rece la mal.
Oscilatiile bruste ale temperaturii se produc mai ales la schimbarea sensului curentilor in lungul coastei. Circulatia nordica provoaca adventia la mal a maselor de apa de adancime, cea ce determina in sezonul cald scaderi bruste ale temperaturii apei, iar in sezonul rece al anului, cresteri. De exemplu, in luna iulie, temperatura apei marii la coasta poate sa scada, in conditiile vanturilor din sectorul sudic, de la 24°C la 7-8°C. Iarna, in conditii asemanatoare, temperatura apei marii creste in sectorul malurilor de la 1°C la 6-7°C.
Cuveta Marii Negre se caracterizeaza printr-un echilibru salin dictat de schimbul de apa prin stramtoarea Bosfor, existand o egalitate intre debitele de sare afluente si cele efluente.
Atat in jumatatea de vest, cat si in jumatatea de est a Marii Negre, in perioada de iarna, cand stratul superior al marii se gaseste intr-o stare de amestec convectiv, salinitatea la suprafata marii in zona de larg oscileaza in jurul valorii de 18,15%o. Spre profunzime, pana la circa 50 m, salinitatea oscileaza foarte putin iar sub orizontul de 50 m, creste foarte rapid. Salinitate de peste 20,0 se intalneste in zona centrala a marii, la adancimi de 70-80 m, pe coastele sudice la adancimi de 175 m si pe cele nordice la adancimi de peste 200 m (Fig. 5).
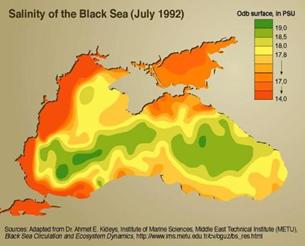
(https://maps.grida.no/go/graphic/salinity_of_the_black_sea)
Partea nord-vestica a Marea Neagra se caracterizeaza prin indulcirea deosebita a apelor, datorat in special aportului masiv de apa dulce din fluvii (Dunarea, Nispru, Nistru). In aceasta parte a marii, in cursul verii salinitatea coboara la suprafata pana la valori de 14-16 in zona de larg si pana la 5-10 la coasta. In restul spatiului marii, in afara sectoarelor din apropierea gurilor raurilor,
salinitatea oscileaza in jurul valorilor de 17-18 Bondar si al Rapoarte INCDM, 1998-2006
Caracteristic pentru litoralul romanesc este faptul ca distributia pe verticala a salinitatii sufera oscilatii functie de sector si sezon. Astfel, salinitatea marii creste de la suprafata spre zonele profunde, valorile salinitatii din fiecare orizont variind functie de timp.
Stratificatia salina transversala pe linia de tarm este puternic influentata de circulatia longitudinala a masei de apa si indeosebi de curentii formati sub actiunea vanturilor. In conditiile circulatiei sudice, paturile superficiale ale apei mai putin saline sunt impinse spre mal, marind coloana stratului de apa dulce langa tarm si producand o impingere in jos a izohalinelor, iar in conditiile circulatiei nordice, paturile superficiale de langa mal sunt antrenate spre larg, impingand izohalinele in jos in zona de larg. Tot in cazul circulatiei nordice, in paturile de fund apare tendinta unui aport de ape saline din larg, care iau locul maselor de apa de la mal, impinse spre larg, ridicand in sus izohalinele de fund (Bondar si al., 1972).
Procesul de stratificare al maselor de apa langa tarm este de asemenea influentat de starea vanturilor. Vanturile din sectorul nordic, intensificand circulatia sudica, conduc la accentuarea neomogenitatii saline, din cauza impingerii spre sud a maselor de apa indulcita din zona gurilor Dunarii. Vanturile din sectorul sudic, dand nastere circulatiei nordice, in conditiile careia spre tarmurile romanesti vin mase de apa cu salinitati omogene, din partea sudica a marii, conduc la o omogenizare a izohalinelor.
Caracteristica distributiei plane a salinitatii apelor Marii Negre in zona litoralului romanesc o constituie tendinta de scadere a salinitatii de la sud spre nord si din zonele de larg spre coasta, datorita procesului de indulcire al apelor marii, sub influenta debitelor fluviului Dunarea.
Stratificarea salina prezinta urmatoarele caracteristici:
- in stratul de apa cuprins intre 0-10 m, salinitatea are valori valori sub 16
- in stratul de apa cuprins intre 10-25 m, salinitatea este in jurul valorii de 17
Marea Neagra are doua paturi de apa suprapuse: una la suprafata (10-200 m) patura de apa cu oxigen - zona oxidarii, cu un dinamism remarcabil, cu conditii favorabile existentei vietii si cea de-a doua patura, de fund (sub 150-200 m), cu o salinitate ridicat si o stabilitate termica pronuntata, un dinamism redus practic la zero, lipsita de oxigen, dar bogata in hidrogen sulfurat - zona de reducere, lipsita de viata, cu exceptia bacteriilor.
Oxigenul dizolvat in apa Marii Negre exista numai in stratul de suprafata, in zonele centrale ale marii pana la adancimi de 125 m si in arialele marginale ale marii pana la adancimi de 225 m (Black Sea Pilot).
In apa marii, continutul de oxigen pana la adancimi de 100 m oscileaza in jurul valorii de 55-95%, la adancimi de 100-150 m descreste brusc pana la 17-28% iar la adancimi peste 200 m atinge 0,21% mg/l. La adancimi de 300-350 m, urmele de oxigen dispar si din zonele marginele ale marii.
In zona cu adancimi mari, primul strat de salt al oxigenului este situat intre 25-50 m adancime, unde continutul de oxigen scade pana la 33-70%. Un salt mai brusc se intalneste la limita convectie verticale care se produce intre orizonturile de 50-70 m, unde oxigenul scade pana la 4-10 %. Mai jos de orizontul de 150 m adancime distributia verticala a oxigenului este mult mai uniforma (Cociosu si al., 1983). Circulatia sudica imbogateste cu oxigen apele de langa tarm, pe cand circulatia nordica introduce in paturile de fund apa cu cantitati mai mici de oxigen, adusa din adancurile marii.. Uneori masele de apa marina sarace in oxigen ajung pana la mal in conditiile circulatiei sudice, formand un brau lat de circa 4 km de-a lungul tarmului (Rapoarte INCDM, 1998-2006).
Se poate mentiona ca o caracteristica a oxigenului dizolvat, slabirea dependentei de regimul termic si cresterea dependentei de procesele biologice. In acest sens s-a observat suprasaturarea stratului superficial, surplus de oxigen de natura biologica, determinat de cresterea productiei primare planctonice si deficitul de oxigen din vecinatatea fundului, creat prin consumul intens de oxigen in procesele de mineralizare a materiei organice moarte. Cunoscut sub numele de hipoxie, acest deficit de oxigen intalnit in special in perioada calda (vara), de regula intre 10-40m, reprezinta
cauza mortalitatilor in masa semnalate la nivelul comunitatilor bentale (Gomoiu, 1985).
Scaderea gradului de oxigenare a apelor marine litorale, particularitate mult mai pronuntata in extremitatea sudica, semnifica inrautatirea calitatii apelor marine costiere. Afirmatia este confirmata de valorile saturatiei in oxigen care reflecta permanetizarea deficitului de oxigen in ultimii ani, starea de subsaturare fiind mai accentuata in extremitatea sudica (Rapoarte INCDM, 1998-2006). Starea de subsaturare intalnita ocazional in anii anteriori extinzandu-se la nivelul intregului ciclu evolutiv.
Continutul mare de hidrogen sulfurat, in straturile de adancime ale Marii Negre, reprezinta una din cele mai importante particularitati ale acestei mari. Hidrogenul sulfurat imparte masa de apa a Marii Negre in doua parti, partea inferioara numita 'zona moarta' si partea superioara numita 'zona vie'. Granita hidrogenului sulfurat in paturile centrale ale marii este situata la circa 125 m adancime, iar in paturile periferice, la 175 m (Black Sea Pilot). Stratul de intrepatrundere reciproca a oxigenului cu hidrogenul sulfurat din zona moarta are o grosime de 50 m, osciland in diverse sectoare ale marii intre 7 si 50 m. Distributia plana a hidrogenului sulfurat are aceiasi caracteristica ca si distributia altor elemente hidrochimice examinate.
Principalele surse de producere a hidrogenului sulfurat sunt: descompunerea continutului de sulf din substante organice si prin reducerea sulfatilor sub influenta bacteriilor. Descompunerea substantelor organice se realizeaza prin participarea bacteriilor genului Bacterium si Bacillus, iar
reducerea sulfatilor cu bacteriile Microspira. Astfel, ambele surse de formare a hidrogenului sulfurat sunt procese biochimice. In Marea Neagra, hidrogenul sulfurat se formeaza prin descompunerea organismelor care contin sulf, sub influenta bacteriilor. Totusi, o importanta primordiala o are si cel de-al doilea proces, reducerea sulfatilor de catre carbonul din substantele organice, sub influenta bacterilor Microspira. Procesul formarii hidrogenului sulfurat, prin reducerea sulfatilor, conduce la randul lui la formarea unor cantitati si mai mari de sedimente pe fundul marii. Prin formarea hidrogenului sulfurat, pe baza descompunerii sulfatilor, se formeaza oxizi cu miros neplacut.
In ciuda relativei stabilitati a distributiei hidrogenului sulfurat, nivelul interfetei ce separa apa oxigenata de apele saracite de oxigen are fluctuatii in concordanta cu oceanografia fizica a regiunii (Aubrey si al., 1996).
Observatii sistematice efectuate la litoralul romanesc asupra fosforului total dizolvat, au scos in evidenta faptul ca apele marine contin cantitati semnificativ mai importante de fosfor organic decat anorganic. Se poate estima ca fractiunea organica poate constitui o importanta rezerva nutritiva disponibila, in conditiile diminuarii drastice a nivelului fosforului mineral.
Scaderea stocului de fosfati se manifesta la nivelul intregului platou continental romanesc. Elocvente pentru sustinerea acestei afirmatii sunt concentratiile extrem de scazute, in jurul valorii de 0,1-0,15 µM, inregistrate in anul 1997, in sectoarele deschise ale marii, precum si frecventele cazuri de epuizare sau valori apropiate de zero. Procesul, surprins in urma cu 4-5 ani pe arii restranse, s-a extins considerabil, afectand uneori intreaga coloana de apa 0-50 m, asa cum s-a intamplat in timpul ultimelor veri (Rapoarte INCDM, 1998-2006).
In contrast cu fosfatii, evolutia silicatilor, asa cum rezulta din analiza datelor inregistrate in statiile din dreptul Constantei (Raport IRCM, 1998), a fost marcata de o evidenta crestere a nivelului actual de concentratii in raport cu nivelul mediu de referinta, cele mai mari abateri pozitive fiind inregistrate in lunile ianuarie, februarie, mai si decembrie. Daca luam in consideratie mediile anuale, reprezentand 22,3 µM in anul 1998 si 12,3 µM in intervalul 1980-1997, se poate constata aproape o dublare a continutului actual de silicati in mare, in conditiile in care cantitatile transportate de Dunare au fost doar usor mai ridicate decat cele din anii anteriori. Acumularea actuala de silicati din mare se poate pune, in principal, pe seama repunerii sale in circuit din depunerile sedimentare, compensand astfel utilizarea sa de catre organismele consumatoare (diatomee).
In Marea Neagra, factorul dominant care guverneaza productivitatea primara este disponibilitatea de azot. Examinarea sarurilor minerale de azot incepe cu azotul amoniacal, care reprezinta forma predominanta de azot din apa marii. Datorita utilizarii sale de catre fitoplanctonul marin, in stratul eufotic continutul de azot amoniacal suporta importante variatii sezoniere. Sub adancimea de 150-200 m continutul de azot-amoniacal creste sensibil, ca urmare a intensificarii activitatii bacteriilor nitrificatoare. La 2000 m, continutul de azot amoniacal inregistreaza valori de 1300-1400 µg / l N. O astfel de distributie a azotului amoniacal reprezinta o alta particularitate a Marii Negre, in comparatie cu marile si oceanele bine oxigenate.
Una din caracteristicile principale ale Marii Negre o constituie continutul ridicat de amoniu, nu numai la adancimi, dar si in straturile superficiale. Dupa cum este cunoscut, oxidarea materiei organice are loc in majoritatea cazurilor in prezenta oxigenului liber, iar trecerea azotului organic in azotati este aproape totala. In bazinele bine aerate, oxidarea substantelor organice cu azot , la azotati (nitrati), are loc in toata coloana de apa, procesul extinzandu-se chiar si sub interfata apa-sediment (Holland, 1983).
In Marea Neagra, datorita existentei conditiilor de oxidare in stratul superior (0-150-200 m) si de reducere sub aceasta adancime, nitratii lipsesc complet in zona hidrogenului sulfurat. Aceasta reprezinta o alta particularitate prin care Marea Neagra se deosebeste de restul bazinelor bine oxigenate.
In zona eufotica, nitratii, asemenea celorlalte elemente nutritive, suporta importante variatii sezoniere, determinate in principal de ciclu evolutiv al consumatorilor (fitoplanctonului). Azotitii (nitritii) apar ca rezultat al reducerii azotatilor, prin dezintegrarea si mineralizarea detritusului organic, care cade din straturile de suprafata. De asemenea, el apare si ca rezultat al acidularii amoniacului, care se formeaza in procesul amonificarii substantelor azotoase ale organismelor planctonice care mor in stratul de suprafata. In Marea Neagra, distributia pe verticala a nitritilor este identica cu cea din oceane si din alte marii. Pe intreg acvatoriul Marii Negre, se constata absenta nitritilor in stratul de fotosinteza si concentrarea lor in stratul de 50-100 m.
Pe ansamblu, sarurile de azot au avut nivele de concentratii si evolutii diferite de la o zona la alta, Dunarea ramanand principala sursa de azotati pentru apele marine de pe platoul contimental romanesc. Spre deosebire de perioada 1988-1992, cand nivelul azotatilor din Dunare depasea de circa 30-40 ori nivelul lor din zona marina costiera, in ultimii ani, aceste decalaje s-au redus considerabil. Aceasta situatie a fost determinata de declinul produs in evolutia acestor saruri minerale de provenienta fluviala al caror nivel de concentratie s-a diminuat continuu din anul 1988, ajungand sa reprezinte, in prezent, aproximativ ¼ din nivelul mediu de la inceputul perioadei de referinta. Desi la scara mai mica, acelasi proces de reducere al stocului de azotati s-a inregistrat si pe platoul continental, nivelul actual de concentratie situandu-se sub nivelul multianual.
Referindu-se in ansamblu la evolutia sarurilor de azot in sectoarele de mica adancime de la litoralul romanesc, trebuie sa mentionam faptul ca, spre deosebire de apele fluviale unde azotatul continua sa reprezinte forma dominanta de azot, in apele marine amoniul a devenit preponderent. Aceasta rasturnare a raportului intre cele doua forme de azotati si azotiti, s-a produs la nivelul anilor 1988-1989 si reprezinta inca una din consecintele negative ale eutrofizarii asupra factorilor de mediu abiotici.
Stabilirea factorilor de mediu cu impact asupra bioresurselor Marii Negre
Conceptia unitatii dintre organisme si mediul lor constituie principiul teoretic fundamental al ecologiei contemporane, inclusiv al ecologiei pestilor. Orice organism, populatie, specie, traiesc pe seama mediului lor specific; in absenta unei interactiuni cu mediul, organismul isi inceteaza existenta. Specificul viului consta tocmai in interactiunea sa adaptiva cu mediul care ii asigura dezvoltarea si existenta (Nikolski, 1962).
Raporturile dintre organism si diferitele elemente ale mediului sau abiotic si biotic nu exista izolat unele de altele, ele aflandu-se intr-un sistem de relatii unitar, indisolubil. Modificand in natura o anumita legatura biotica sau abiotica, provocam inevitabil o restructurare a intregului sistem de relatii reciproce dintre organism si mediu.
Studiile anterioare au accentuat comportamentul pestelui ca o reactie la stimulii mediului inconjurator, in timp ce lucrarile mai recente tind sa coreleze comportamentul pestelui si cu performanta uneltelor de pescuit. De asemenea, multe din studiile anterioare tind sa generalizeze comportamentul observat, in timp ce lucrarile mai recente remarca faptul ca fiecare specie si fiecare grup de lungime/varsta pot avea unele reactii specifice si distincte. Chiar stocuri diferite ale aceleiasi specii pot reactiona diferit ca raspuns sezonier la modificarile de mediu (ex. migratiile), de asemenea acest raspuns se poate modifica in timp (ex. rutele de migratie).
Comportamentul (migratiile verticale diurne si cele orizontale, precum si agregarea, dispersia, etc.) si in consecinta accesibilitatea pestelui este determinata de o multitudine de factori de mediu care si ei la randul lor sunt influentati si/sau determinati de conditiile meteorologice.
Cel mai usor si cel mai eficient mod de a imbunatati captura pe unitatea de efort este analiza conditiilor de mediu si prognozele. Pentru a realiza aceasta, trebuie cunoscut comportamentul (migratia, agregarea, etc.) pestelui in relatie cu conditiile si schimbarile de mediu. In plus, trebuie sa fie disponibile prognozele si analizele sinoptice ale conditiilor de mediu. Aceste analize sinoptice ale conditiilor de mediu si ale comportamentului pestilor ar trebui sa furnizeze o baza reala pentru prognoza pescuitului.
In multe zone de pescuit sunt necesare prognoze speciale pentru regiunile costiere, deoarece conditiile meteorologice in zona costiera pot fi considerabil diferite de cele de pe uscat sau din largul marii. Aplicarea oceanografiei pescaresti la solutionarea problemelor practice pescaresti nu inseamna numai imbunatatirea productiei pescuitului, dar si intelegerea biologiei pestilor si a dinamicii populatiilor.
Desi incercam sa rezumam comportamentul pestelui din punct de vedere al unei cauze, trebuie avut tot timpul in minte ca reactiile pestelui pot varia considerabil si ca mai multi stimuli pot fi prezenti la un moment dat. Relatia aparent simpla dintre un organism si orice componenta particulara a mediului sau nu este niciodata izolata de influenta si interactiunea cu alte componente de mediu.
Influenta factoriilor abiotici
1.1. Influenta curentilor marini asupra distributiei si comportamentului pestilor
Majoritatea cercetarilor arata ca advectia este unul dintre factorii cei mai importanti ce determina schimbari locale in conditiile de mediu prin raspandirea altor factori de mediu precum temperatura, salinitatea, etc. Este de asteptat ca pestele sa reactioneze la acele conditii de mediu care sunt determinate de curenti si de asemenea, la actiunea mecanica directa prin transportul pestilor, al larvelor si icrelor la distante uneori foarte mari.
Migratia pestilor adulti poate fi influentata de curenti prin faptul ca acestia servesc si ca punct de orientare, organul care percepe miscarea apei fiind linia laterala. De asemenea, prin curenti, indeosebi cei de flux, poate fi influentat comportamentul diurn al pestelui.
In special la marginile sale, un curent poate afecta distributia pestilor adulti, fie prin efectul direct asupra lor sau indirect prin agregarea hranei si producerea altor limite de mediu (temperatura, salinitate, turbiditate etc.) care influenteaza abundenta si limitele geografice de distributie a speciilor din lantul trofic.
Un impact deosebit asupra stocurilor de peste o are influenta curentilor asupra stadiilor timpurii de dezvoltare, respectiv icre si larve. Excelente investigatii asupra acestui subiect au fost realizate de Walford (1938), Carruthers, Lawford si Veley (1951), Fraser (1958). De cele mai multe ori, recrutarea unui stoc de peste depinde foarte mult de transportul icrelor si larvelor de la locurile de reproducere catre zonele de crestere si hranire.
La maturare, unele specii incep migratia activa, inotand impotriva curentului pana la locurile
de reproducere. Dupa reproducere pestele poate fi extenuat, slabit, fiind transportat de catre curent spre locurile de hranire, ciclul repetandu-se anual.
Comportamentul pestelui adult in legatura cu curentul apei, precum si cu alte conditii de mediu, depinde de starea fiziologica, marimea exemplarelor si de la specie la specie, fiind un rezultat al efectului combinat al mai multor factori. De exemplu, este simplu de corelat comportamentul diurn al pestilor cu ciclul diurn al conditiilor de lumina si efectul curentilor, indeosebi cei de maree, fiind multe specii care inoata activ impotriva curentului in timpul zilei, in timp ce noaptea plutesc antrenate de curent.
Temperatura apei afecteaza atat migratia pestelui, cat si transportul lor de catre curenti, observandu-se ca in apa rece, in apropiere de 0oC, pestele este putin activ si purtat de curent.
Unul dintre factorii principali care afecteaza periodic disponibilitatea pestelui in pescuit este dominanta curentului care, la randul sau, este influentata de variatiile in timp ale intensitatii si duratei vantului.
Rodewald (1960) a analizat fluctuatiile in debarcarile de peste si disponibilitatea diferitelor stocuri de peste in corelatie cu anomaliile vantului din Marea Barent, rezultand ca variatiile la scara mare in presiunea atmosferica determina disponibilitatea in pescuit a unui numar important de specii comerciale.
Prin prognoza presiunii atmosferice, a directiei si vitezei vantului, precum si a curentilor rezultati, vom fi capabili de a prognoza disponibilitatea in pescuit in zonele de activitate.
Zusser (1958) sustine o teorie conform careia exista o agregare semnificativa a pestilor comerciali in mod obisnuit in centrul curentilor circulari, unde pestele se odihneste sau se hraneste, aici existand o acumulare de plancton, icre si puiet de peste (Orton, 1937; Redfield, 1941).
Vartejuri locale sunt determinate de morfologia coastei si configuratia fundului. De aceea, cunoasterea in detaliu a curentilor locali joaca un rol important cand cautam concentratiile pescuibile de pesti.
Convergenta curentilor provoaca o agregare mecanica a organismelor ce constituie hrana pestilor (Laevastu, 1962), linia de convergenta dintre curentii calzi si reci fiind in mod special bogata in organisme.
Destul de frecvent, aglomerarile de pesti demersali sunt determinate de marginile curentului care, de cele mai multe ori depind de particularitatile locale precum: panta, stabilitatea in spatiu si timp a limitelor din apropierea fundului, disponibilitatea hranei de o parte si de alta a marginilor curentului. Marginile curentului isi schimba pozitia cu variatia conditiilor meteorologice sau sezonier, localizarea lor fiind cu subiect de interes pentru oceanografie si de mare importanta pentru pescarie.
Sedykh (1979) studind relatiile dintre conditiile hidrologice, in special a curentilor de upwellig si ciclu biologic al diferitelor specii de pesti pelagici, pune in evidenta raporturile directe existente intre variatiile intensitatii upwellingului cu variatiile perioadei de maturare, ritmul de crestere, abundenta numerica pe grupe de varsta si a biomasei.
Deosebirile calitative si cantitative ce se inregistreaza in pescuitul cu talienele din cele trei zone de la litoralul romanesc (sudica, centrala si nordica) se datoresc si influentei pe care o exercita curentii asupra conditiilor fizico-chimice si biologice ale mediului marin. In acelasi mod se explica de regula si deosebirile mari ce se semnaleaza intre cantitatile realizate in aceeasi luna, de la un an
la altul, chiar atunci cand capturile anuale sunt sensibil egale.
Pescuitul sprotului, specia cu cea mai mare pondere, este determinat in mare masura de frecventa curentilor reci de fund. Fiind un peste criofil ce nu suporta temperaturi mai ridicate de cca. 16sC, el se indeparteaza de tarm la inceputul verii si in tot cursul acestui sezon nu mai revine decat odata cu aparitia fenomenului de 'upwelling'. Cum in zona gurilor Dunarii, deplasarea apelor superficiale de la tarm spre larg este aproape permanenta (datorita actiunii curentului fluvial), urcarea maselor de apa de adanc catre tarm este mai accentuata aici decat in restul litoralului. De aceea cantitatile pescuite de sprot, vara in zona nordica, sunt incomparabil mai mari decat cele din zona centrala si sudica, unde scaderile de temperatura in stratul 0-10 m, sub 18sC, sunt destul de rare si se mentin pe intervale foarte scurte de timp.
1.2. Influenta valurilor
Obtinerea datelor privind influenta valurilor asupra miscarii verticale a pestilor este extrem de dificila dat fiind faptul ca aceste miscari sunt influentate de asemenea si de alti factori (lumina, temperatura, hrana, etc.).
Este cunoscut faptul ca pestele evita straturile superioare ale apei pe vreme rea, cu valuri puternice, coborand de obicei in straturile mai adanci ale apei, unde actiunea valurilor nu se resimte.
O influenta deosebit de mare exercita asupra pestilor valurile din zonele litorale ale marii, unde forta unui val poate atinge pana la 1,5 t/m2. Pestii care traiesc in zona litorala se caracterizeaza
prin adaptari speciale, care ii apara pe ei insisi, ca si icrele lor de bataia valurilor.
Valurile nu exercita numai o influenta mecanica directa asupra pestilor, ci si o mare influenta indirecta, contribuind la amestecarea apei si coborarea saltului termic.
In apele de mica adancime furtunile sporesc turbiditatea apei si limiteaza distributia catre tarm a unor specii care nu suporta conditii de turbiditate.
1. Influenta temperaturii apei marine
Dintre numerosii stimuli de mediu ce afecteaza pestele, temperatura apei s-a bucurat de cea mai mare atentie, fiind factorul de mediu cel mai usor de observat si de aceea majoritatea cercetarilor au incercat sa coreleze comportamentul si abundenta pestelui cu temperatura apei si fluctuatiile sale. Efectele sale asupra pestelui au fost rezumate printre altii de Laevastu si Hela (1970); Laevastu si Hayes (1981); Laevastu si altii (1996) si Nikolski (1962).
In cele mai multe cazuri, temperatura apei constituie indicatorul cel mai folositor pentru a determina schimbarile conditiilor ecologice, dar nu numai temperatura reala, cu domeniile sale de fluctuatii, ci si gradientii orizontali si verticali, ce variaza de la un loc la altul, trebuie sa fie luati in calcul cand folosim temperatura ca indicator ecologic sau indirect.
Sulivan (1954) a aratat efectele temperaturii apei asupra miscarii si distributiei pestelui, prezentand rolul si mecanismul de receptor al sistemului nervos central ca raspuns la temperatura. Aceasta poate actiona asupra pestelui ca: stimul nervos, modificator al proceselor metabolice si modificator al activitatii corpului.
Schmidt (1931) si Nikolski (1962) au aratat efectul temperaturii apei asupra caracterelor meristice ale pestilor, numarul vertebrelor si al radiilor crescand cand temperatura scade.
Efectele temperaturii asupra pestelui se manifesta in pescarie in diverse moduri. Cum aceasta afecteaza activitatea si mobilitatea corpului, atunci cand este scazuta poate afecta posibilitatea pestelui de a scapa de unealta de pescuit si de asemenea, abilitatea pestelui de a prinde hrana mobila.
Temperatura poate determina o diferentiere in distributia regionala a juvenililor si adultilor, deoarece acestia au cel mai adesea o preferinta si toleranta diferita pentru aceasta (Alverson, Pruter si Ronholt, 1964).
Concluzia este ca mai exista si alti factori alaturi de temperatura care influenteaza abundenta si distributia organismelor. In plus la influenta directa, schimbarea temperaturii apei poate indica schimbari si ale altor factori de mediu.
a. Influenta temperaturii apei marine asupra reproducerii, eclozarii, supravietuirii si dezvoltarii larvelor
Influenta temperaturii apei asupra comportamentului pestilor este cea mai pronuntata in timpul reproducerii. Insa si inainte de reproducere, are o importanta ridicata, deoarece influenteaza maturarea produselor sexuale. Fenomenele au fost descrise printre altii de Poulsen (1944), Hodder (l965), Nikolski (1962).
Fiecare stoc de pesti are un domeniu de temperatura optima pentru reproducere. Temperatura in afara limitelor optime face ca maturarea gonadelor sa intarzie, sau sa fie devansata. De aceea, factorul temperatura este decisiv pentru sosirea in timp a stocurilor de pesti pe locurile obisnuite de reproducere.
Temperaturi neobisnuite in zonele de reproducere si in timpul sezonului de reproducere obliga pestele sa se reproduca si in alte zone decat acelea in care el merge in mod obisnuit pentru aceasta (Simson, 1953).
Dezvoltarea icrelor si larvelor constituie indubitabil perioada cea mai critica din viata unui peste, deoarece acum sunt cel mai puternic afectati direct si indirect de conditiile fizice. Temperatura influenteaza direct rata de dezvoltare si in conjunctie cu salinitatea va determina densitatea apei, afectand astfel flotabilitatea icrelor. De asemenea, perioada de incubare si durata vietii larvare depind direct de temperatura mediului care, in conjunctie cu intensitatea curentilor, vor determina intensitatea eclozarii si rata de supravietuire a larvelor (Rounsefell, 1930; Uda si
Honda, 1934; Kurita, 1959).
In realitate exista mai multe feluri in care temperatura influenteaza supravietuirea larvelor.
Cel mai important fiind probabil efectul asupra disponibilitatii hranei. Este evident ca disponibilitatea hranei corespunzatoare pentru larve, la timpul potrivit, este legata de productia fitoplanctonica care, la randul ei, este legata de schimbarile sezoniere ale temperaturii si cantitatii de lumina. In general, temperatura prea inalta sau prea scazuta, poate pune perioada de dezvoltare a larvelor in afara intervalului optim.
Ca rezumat al celor aratate mai sus, se poate spune ca urmarind temperatura si schimbarile sale in zonele de reproducere, temperatura in zonele ocupate mai tarziu de larve si puiet, disponibilitatea hranei (plancton) si cunoscand temperatura optima a unui stoc de peste dat, se poate prognoza rata de supravietuire a larvelor eclozate si bogatia claselor anuale.
In general este acceptata ideea ca exista o variatie a succesului reproducerii si ca aceasta nu depinde numai de marimea stocului de reproducatori (Murphy, 1966), ci si de conditiile de mediu din timpul eclozarii si dezvoltarii larvelor.
b. Influenta temperaturii apei marine asupra hranirii, metabolismului si cresterii
Ratele de hranire, metabolism si crestere sunt afectate nu numai prin disponibilitatea hranei, ci si indirect prin temperatura apei. La o temperatura in afara limitelor optime, activitatea de hranire este in mod obisnuit redusa (Komarova, 1939).
Dupa Brett (1958), exista un nivel al temperaturii la care diferenta dintre ratele metabolice din timpul odihnei si din perioada activa este maxima. De aceea, exista o temperatura optima pentru rata metabolica (viteza de convertire a materialului nutritiv in materie vie) si implicit pentru o crestere optima. Desi cele mai multe specii de pesti au un domeniu larg al tolerantei fata de temperatura, in mod obisnuit, cresterea este optima intr-un domeniu mai limitat in special in partea superioara a domeniului (Wise, 1958).
Dat fiind nevoile fiziologice, pestele mare dintr-un stoc cauta temperaturi mai mici fata de restul exemplarelor. Acest lucru poate fi aplicat in interpretarea distributiei spatiale, exemplarele mari migrand spre marginile zonelor de distributie a stocului, evitand zona normala de reproducere, facand ca reproducerea sa nu fie tocmai un succes.
Cunoscand toate aceste elemente, le putem folosi pentru pescuitul mai intens a cohortelor mai varstnice.
c. Influenta temperaturii apei marine asupra abundentei, migratiei si formarii aglomerarilor de peste
Aproape toate stocurile de peste au temperaturi optime specifice. Cunoasterea exacta a aces-tora este necesara pentru prognoza concentratiilor de peste. Cu astfel de cunostinte si beneficiind de prognozele temperaturii (statistic sau sinoptic), putem sa prognozam abundenta sezoniera a unui stoc de peste. Problema se complica in mod suplimentar prin faptul ca cerintele fata de mediu se schimba in functie de etapa de crestere, precum si sezonier, cel putin in legatura cu activitatea de reproducere (Devold, 1959) si hranire.
Limitele superioare sau inferioare de temperatura la care pestele poate supravietui depind de aclimatizarea sa anterioara. De aceea, schimbari bruste ale temperaturii sunt in mod normal mult mai periculoase decat schimbarile lente care-i dau posibilitatea de a se aclimatiza.
d. influenta temperaturii apei marine asupra migratiei de primavara a pestilor la litoralul romanesc
Toate speciile industriale de pesti de la litoralul romanesc al Marii Negre, parasesc zonele de iernat si migreaza spre tarm sau in Dunare pentru reproducere sau hranire, mai devreme sau mai tarziu, in functie de existenta unui regim termic al apei marine. Speciile ce executa migratii de reproducere (sturionii, scrumbia de Dunare) suporta temperaturi mai scazute decat celelalte specii de pesti.
La specia sprot ce executa primavara o migratie de hranire, rolul temperaturii este mult mai important, fenomenul fiind conditionat de dezvoltarea unor organisme planctonice ce constituie hrana lor de baza. Observatiile efectuate de-a lungul mai multor ani au demonstrat ca indiferent de particularitatile celorlalti factori mediali, chiar daca ating valorile optime, aceasta specie nu apare la tarm decat atunci cand s-a produs o incalzire a apei pana la un anumit grad si anume de cca. 7oC in cazul sprotului, 9oC. De remarcat ca aceasta valoare este superioara celor existente in stratul de minima temperatura, valori pe care le suporta in timpul iernarii.
Din analiza pescuitului cu talienele se constata ca sprotul apare in pescuit chiar in ultima parte a lunii martie, in anii cu primaveri foarte timpuri.
In luna iunie, asistam in continuare la schimbari si mai importante ale raportului dintre cele trei specii pelagice. Incalzirea accentuata a apei determina o retragere a sprotului din zona costiera, in schimb apar carduri mari de hamsie. Rezulta, ca in conditiile iernarii acestor specii in fata litoralul romanesc si migratiei lor timpurii ca urmare a existentei unor conditii termice favorabile, se pot realiza capturi bune la taliene, in sezonul de primavara.
e. Influenta zonei saltului termic asupra distributiei pestilor pelagici in masa apei
In sezonul de vara, distributia sprotului si hamsiei este determinata de zona saltului termic.
Sprotul, specie prin excelenta criofila, se indeparteaza de litoral cand temperatura apei depaseste 18 C si revine numai in cazul existentei curentilor reci de fund (Cautis, 1958, Serpoianu, 1964). Sprotul se mentine numai in stratul de apa de sub saltul termic. In schimb, hamsia, fiind specie termofila, executa migratia de hranire catre tarm mai tarziu decat sprotul, iar pe timpul verii se disperseaza in vederea reproducerii pe intreg platoul continental, mentinandu-se in patura superficiala a apei, deasupra saltului termic.
Stratul de apa in care se situeaza saltul termic reprezentand totodata zona limita de contact a organismelor planctonice termofile si criofile este in general mai bogat in hrana. Astfel, orizontul unde se inregistreaza marile diferente ale temperaturii intereseaza in mod cu totul deosebit, furnizand indicatii pretioase asupra adancimii la care diferite specii de pesti se mentin in sezonul cald al anului.
In luna iunie saltul termic se situeaza intre minimum 5-10 m si maximum 15-20 m, diferentele de valori pentru orizonturile mentionate reprezentand 8 C si respectiv 7 C. Procesul de incalzire al apei din lunile urmatoare (iulie si august) determina o crestere simtitoare a valorilor temperaturii din stratul superficial si o coborare a saltului termic, care nu depaseste insa orizontul 25-30 m. In cazul dominarii vanturilor de vest, concomitent cu formarea curentilor superficiali de est, la fund iau nastere curenti de directie contrara, ceea ce face ca saltul termic sa se intalneasca pentru intervale foarte scurte intre numai 5 si 10 m, chiar in lunile iulie si august.
f. Influenta temperaturii apei marine asupra starii de sanatate a pestilor
Dintre factorii abiotici, temperatura apei reprezinta pentru pesti, animale poikiloterme lipsite de sisteme termoreglatoare, unul dintre principalii factorii de viata. Scaderea sau cresterea in afara limitelor normale, suportabile de catre pesti, dar mai ales schimbarile bruste de temperatura, semnalata in apele costiere romanesti, provoaca tulburari grave ca: paralizia aparatului branhial, a inimii si tulburari sanguine.
Factorul termic are asupra pestilor nu numai o actiune directa ci si indirecta, modificand alti factori de mediu. La temperaturi ridicate, semnalate deseori in ultimii ani, scade continutul in oxigen, care determina hipoxia. Efectul hipoxiei, manifestat prin asfixie, se accentueaza datorita faptului ca la temperaturi ridicate metabolismul se mareste, crescand si necesarul de oxigen.
1.4. Influenta salinitatii apei marine
a. Influenta salinitatii asupra starii si comportamentului pestilor
Un factor deosebit de important, care determina posibilitatea adaptarii pestilor la apa cu salinitate variata, este constanta mai mult sau mai putin accentuata a mediului intern. Aceasta constanta se realizeaza prin dezvoltarea diferitelor adaptari de osmoreglare, care asigura posibilitatea reglarii presiunii osmotice a lichidelor din corp.
Variatiile salinitatii in zonele de larg sunt relativ mici, insa in zonele costiere sunt considerabile datorita aportului apelor curgatoare. Aceste variatii afecteaza reglarile osmotice ale pestelui si determina flotabilitatea icrelor pelagice.
Desi exista numeroase articole ce descriu aparenta relatie dintre comportamentul pestelui si distributia si concentratia salinitatii apei, aceste relatii nu sunt neaparat directe. Cel mai adesea, variatiile salinitatii indica schimbarea in conditia de stabilitate a maselor de apa. Influenta directa a salinitatii asupra celor mai multe specii poate fi considerata minora. Acest lucru a fost demonstrat de Holliday si Blaxter (1960) care au gasit ca fertilizarea, dezvoltarea si eclozarea icrelor de hering exista in salinitati care variaza de la 5,9 la 52,5 . Toleranta la salinitate a larvelor eclozate atat primavara, cat si toamna s-a situat intre 2,5 si 52,5
b. Influenta salinitatii asupra repartitiei pestilor in sezonul de iarna
In mod obisnuit toate speciile de pesti cu importanta economica, fie ele pelagice (sprot, hamsie, rizeafca, gingirica), bentonice (sturioni, calcan) sau nectonice (scrumbia de Dunare), ierneaza la adancimi cuprinse intre 45-90 m. Mentinerea lor la adancimi mai mici sau mai mari, intre curbele batimetrice mentionate este in functie de conditiile termice specifice fiecarei ierni.
Se cunoaste ca pentru zona cu adancimi de pana la 55 m salinitatea apei marine prezinta oscilatii foarte mici in raport cu stratul superficial al apei. Limitele valorilor intalnite de-a lungul mai multor ani au oscialt intre minim 17,57 si maximum 18,75. De regula, insa la orizontul de 25 m, salinitatea depaseste 17. Sub 50 m salinitatea apei creste in mod lent. Chiar la limita platoului continental, unde efectul curentilor poate determina uneori cresteri simtitoare ale valorilor, s-a constatat ca maximele semnalate nu depasesc totusi la 100 m, 20,63.
Deoarece toate speciile la care ne-am referit se mentin in unele perioade ale anului la salinitati mult inferioare minimelor semnalate la 50 m, rezulta ca in nici o situatie existenta in stratul 50-100 m, a unor valori ale salinitatii apropiate limitelor inferioare, nu pot influenta in vreun sens repartitia pestilor in sezonul de iarna.
c. Influenta salinitatii asupra migratiilor de reproducere si hranire
La litoralul romanesc, actiunea apelor dulci se resimte foarte activ numai in stratul 0-10 m, avand o puternica influenta asupra speciilor pelagice ce populeaza acest strat de apa. Dintre speciile industriale, sunt afectate in special hamsia. Cum aceasta migreaza primavara pentru hranire in zona costiera, posibilitatea mentinerii si deci a pescuitului ei, este in stransa dependenta de particularitatile regimului salin. De-a lungul litoralului romanesc oscilatiile salinitatii de la suprafata sunt foarte mari, procesul de indulcire reducandu-se totusi pe masura ce ne departam de zona gurilor Dunarii. Deosebirile existente de-a lungul litoralului se reflecta clar in rezultatul pescuitului cu talienele.
1.5. Oxigenul si alte gaze dizolvate
a. Influenta oxigenului si a altor gaze dizolvate asupra starii si comportamenlui pestelui
In conditii normale ale marii, continutul de oxigen dizolvat al apei nu devine un factor limitator in distributia pestelui. Totusi, in unele cazuri, oxigenul poate afecta comportamentul pestelui, fiind un factor de intarziere a depunerii icrelor sau controland migratiile pestilor demersali catre zonele cu adancime mica.
O influenta deosebita asupra distributiei pestelui si mortalitatii in masa o are upwellingul stratului de apa putin oxigenata. Un strat de apa putin oxigenat poate exista indeosebi in zona de fund unde oxigenul este consumat prin descompunerea sedimentelor organice si apele sunt slab ventilate.
Cauza upwellingului o constituie vanturile si topografia coastei si a selfului continental in relatie cu dinamica maselor de apa (curentii). In mod obisnuit, pestele se hraneste la marginea zonei cu upwelling normal si migreaza catre coasta in timpul incetarii upwellingului. Cand upwellingul incepe din nou, apele de fund, acum saracite in oxigen, sunt aduse catre suprafata pana in apropierea coastei, avand ca posibil rezultat mortalitatea in masa a pestelui.
Un exemplu extrem de elocvent il constituie situatia de la litoralul romanesc in vara anului 1999. Urmare a persistentei fenomenelor de inflorire algala din lunile iunie si iulie, a temperaturilor caniculare prelungite si a unei slabe agitatii a apelor, in zona costiera s-au inregistrat frecvente situatii de subsaturatie in oxigen, cu o concentratie a acestuia sub 5 cm3/l. Peste efectele prelungite ale acestei stari de fapt, s-au suprapus in prima parte a lunii august efectele aparitiei la coasta a fenomenului de upwelling . Sub influenta unor vanturi din sectorul sudic, a caror actiune s-a resimtit in tot intervalul 06-11.08.1999, in perioada 10-12.08.1999 s-a produs un puternic upwelling manifestat prin impingerea spre larg a apelor de coasta si aparitia la tarm, in compensatie a unor mase de apa rece, cu salinitate ridicata si continut slab de oxigen.
In raport cu perioada 06-09.08.1999, in intervalul 10-12.08.1999, s-a produs o drastica scadere a temperaturii apei (cu pana la 11oC in 3 zile) si o diminuare a concentratiei de oxigen pana la valori de 3,64-4,78 cm3/l. Urmare a acestui fenomen, in Baia Mamaia s-au semnalat cantitati importante de pesti morti sau in stare letargica, care au fost impinse de val spre linia tarmului. Numai pentru jumatatea sudica a baii Mamaia, mortalitatea a fost apreciata la 800-1000 kg/zi.
In afara de oxigen, o deosebita importanta pentru pesti prezinta si alte gaze. Bioxidul de carbon (CO2) chiar si in doze relativ mici provoaca moartea pestilor. Mecanismul actiunii lui asupra pestilor, ca si a altor animale, se reduce la diminuarea capacitatii de a fixa oxigenul. Este adevarat ca in apa, in special in apa de mare, cantitatea de bioxid de carbon este mica, deoarece magneziul, calciul si alti cationi il fixeaza rapid. Cantitatea de CO2 in apa este in legatura directa cu concentratia ionilor de hidrogen, a carei variatie exercita de asemenea asupra pestilor o influenta apreciabila.
Paralel cu cresterea aciditatii apei, scade intensitatea metabolismului gazos, in schimb ritmul respirator se accelereaza in mediul acid, in comparatie cu mediul neutru si alcalin.
Hidrogenul sulfurat exercita de asemenea asupra pestilor o puternica influenta negativa. El se poate forma si acumula in bazine numai in absenta oxigenului in apa, deoarece in prezenta oxigenului, hidrogenul sulfurat se oxideaza rapid.
Gradul de influenta a hidrogenului sulfurat asupra pestilor difera de la specie la specie si de la un stadiu de dezvoltare la altul. Intensitatea actiunii hidrogenului sulfurat, ca de altfel si a altor multe gaze, variaza apreciabil in functie de temperatura. La temperaturi ridicate moartea survine mai repede decat la temperatura scazuta.
b. Influenta oxigenului si gazelor solvite in apa marina asupra starii de sanatate a pestilor
Dintre gazele solvite in apa, rezultate in urma unor procese naturale care au loc in apa marina, cu influente directe asupra sanatatii pestilor mentionam: oxigenul, hidrogenul sulfurat si pH-ul.
Oxigenul este cel mai important gaz solvit in apa de care depinde viata si sanatatea organismelor marine. Provenit din aerul atmosferic sau ca rezultat al fotosintezei plantelor acvatice, inclusiv a microalgelor (fitoplancton) este indispensabil in procesul de respiratie a pestilor. Valori reduse, sub 4 mg/l, combinate cu cresteri ale temperaturii apei, dezvoltarii fitoplanctonului (in timpul fenomenelor de inflorire), conduc la aparitia starii patologice de asfixie la pesti.
Hidrogenul sulfurat, gaz rezultat din procesele microbiologice de descompunere a substantei organice si de reducere a sulfatilor, provoaca iritatii si intoxicatii care se agraveaza foarte mult in conditiile in care oxigenul este redus.
Ph-ul apei, constituie un element chimic ce poate influenta de asemenea viata pestilor. Apele acide ca si cele alcaline (sub 6,5 si peste 9,0), rezultate in urma deversarilor industriale, produc leziuni grave la nivelul pielii si bronhiilor finalizata cu moartea pestilor.
1.6. Influenta luminii asupra comportamentului si abundentei pestelui
Lumina, iar intr-o masura mai mica si alte forme de energie radianta joaca un rol foarte mare in viata pestilor. Gradul de luminozitate are importanta, atat directa cat si indirecta, in viata pestilor. La majoritatea pestilor, organul vazului joaca un rol esential in orientarea in timpul locomotiei, orientarea fata de prada, de rapitori si de alte exemplare ale aceleiasi specii din bancul respectiv, fata de obiectele imobile, etc.
De gradul de luminozitate este legat intr-o mare masura comportamentul pestilor, in special ritmul nictimeral al activitatii si multe alte aspecte ale vietii (Nikoski, 1962). Lumina exercita o anumita influenta si asupra metabolismului la pesti, precum si asupra maturatiei produselor sexuale.
Astfel, pentru majoritatea pestior lumina constituie un element indispensabil al mediului.
Patrunderea luminii in apa marii este dependenta de intensitatea luminii la suprafata care, in mare parte, este determinata de gradul de acoperire cu nori, de inaltimea soarelui, activitatea valurilor si turbiditatea apei. Diferite lungimi de valuri fac ca lumina sa aiba rate diferite de extinctie in apa, astfel incat compozitia spectrala a luminii se schimba cu adancimea.
Distanta maxima de observatie a unui peste variaza cu conditia luminii in apa si cu turbiditatea. Este cunoscut faptul ca pestii care se hranesc prin vizualizarea hranei, precum tonul, evita apele cu turbiditate ridicata.
Atractia catre lumina poate exista din motive variate precum: fototropism, hranire, curiozitate, dezorientare. Gradul fototropismului pozitiv poate varia de la specie la specie si de la sezon la sezon.
Marimea suprafetei de pe care pestele este colectat cu lumina de pescuit depinde de intensitatea luminii si in special de turbiditatea apei (Laevastu, 1996). De aceea, pescuitul de noapte cu lumina este efectiv, in principal, in apele limpezi.
Distributia verticala si comportamentul diurn al celor mai multe specii variaza de la specie la specie si in cadrul aceleiasi specii, cu varsta, stadiul de maturare al gonadelor, sezon si distributia geografica. Comportamentul poate fi de asemenea afectat de factori de mediu ca: structura termica, curentii de apa si disponibilitatea hranei.
Lumina si vederea pestelui sunt doua elemente de baza exploatate de pescariile cu unelte capcana, mai ales in proiectarea aripii conducatoare, fiind necesar de a spori contrastul vizual al aripii conducatoare prin fire mai groase si colorate. A fost determinat faptul ca galbenul sporeste contrastul (Nomura, 1980). Evitarea uneltei de pescuit descreste cu cat lumina este mai slaba, sub 0,5-0,05 lux (Woodhedd, 1965). Pentru a descreste vizibilitatea si contrastul setcilor si traulului, sunt folosite culorile albastru si verzui pentru plasa.
2. Influenta factorilor biotici
2.1. Relatiile biotice ale pestilor, influenta lor asupra comportamentului si abundentei
Relatiile biotice, atat cele interspecifice, cat si cele intraspecifice, sunt conditionate reciproc si legate in modul cel mai strans de mediul abiotic. De aceea, trebuie sa se aiba in vedere ca legaturile biotice ca si cele abiotice nu exista izolat in natura, ele fiind indisolubil legate (Nikolski, 1962).
Caracterul relatiilor interspecifice ca si al relatiilor intraspecifice se elaboreaza in procesul genezei speciilor ca adaptare la noi conditii de viata. Formarea noilor specii se desfasoara pe calea elaborarii de adaptari la conditiile abiotice si biotice ale zonei geografice in care are loc formarea speciilor respective si poarta de obicei un caracter de grup, formandu-se asa-numitul complex faunistic.
In procesul de formare a complexului faunistic se stabilesc anumite legaturi firesti intre speciile care formeaza complexul si anume: evitarea contradictiilor ce ar putea apare in conditiile consumarii unei hrane asemanatoare prin marirea diversitatii spectrelor de nutritie, in special la pestii de varsta mai inaintata; prin aparitia anumitor adaptari ale pradatorilor si prazii etc. Speciile prada isi elaboreaza prolificitatea necesara, precum si o serie de adaptari de protectie cum sunt: culoarea, tepii, toxicitatea, etc., care le apara intr-o oarecare masura impotriva dusmanilor.
2.2. Relatiile intraspecifice la pesti
Legaturile intraspecifice au scopul de a asigura existenta speciei in conditiile in care ea a aparut si traieste. La pesti, una dintre cele mai importante forme de relatii biotice intraspecifice o constituie formarea bancurilor ce au importanta protectoare, trofica, reproductiva in cautarea rutelor de migratie si a locurilor de iernare.
Rolul protector al bancului de pesti este indiscutabil, pestii solitari sunt devorati de rapitori mult mai repede decat cei din banc. Pestii aflati in banc observa de la distanta mai mare pericolul (rapitorul sau unealta de pescuit). Uneori bancul de pesti are si o importanta protectoare directa impotriva rapitorilor. Astfel, la apropierea unui rapitor, de exemplu a stavridului, hamsia din Marea Neagra se ingramadeste intr-o aglomerare compacta, incepand uneori sa se miste in cerc. Atat timp cat stavridul nu reuseste sa sparga aceasta aglomerare, el nu poate prinde nici macar un singur exemplar (Nikolski, 1962).
Importanta adaptiva trofica a bancului se manifesta cel mai puternic la pestii gregari pelagici, care se hranesc cu zooplancton. In banc pestele descopera mai repede aglomerarile de organisme trofice si pastreaza mai usor contactul cu ele, datorita carui fapt se hraneste mai intens decat in stare solitara (Welty, 1934).
Bancul de pesti are o importanta adaptiva si in timpul migratiilor. Chiar si pestii care duc de obicei o viata solitara, in timpul migratiilor se grupeaza in bancuri, gasind mai repede drumul de migratie, descoperind mai repede diferitele puncte de orientare. Caracterul raporturilor reciproce intraspecifice la pesti este foarte variat. Pestii se influenteaza reciproc si prin modificarea conditiilor abiotice, degajand produsele sale metabolice, fiecare peste exercita prin aceasta o influenta asupra celorlalti pesti din bazinul acvatic.
O foarte mare importanta au raporturile trofice reciproce dintre exemplarele aceleiasi specii. Consumarea indivizilor din propria specie constituie un fenomen destul de larg raspandit la pesti. Canibalismul asigura o slabire a tensiunii relatiilor intraspecifice de ordin trofic, tensiune care se inregistreaza cand in populatie intra o generatie numeroasa. Adeseori prin consumarea puietului propriu, indivizii adulti pot asimila o hrana la a carei consumare directa nu sunt adaptati.
2. Relatiile interspecifice la pesti
Relatiile interspecifice la pesti au un caracter foarte variat, ele putand fi reprezentate prin pradatorism, parazitism, comensalism, raporturi reciproce contradictorii pe baza nutritiei cu hrana asemanatoare, precum si alte tipuri de legaturi.
Pe baza relatiilor pradator-prada , la pestii pradatori se elaboreaza adaptari pentru capturarea prazii si asimilarea ei, iar la pestii prada se elaboreaza in mod corespunzator diferite mijloace de aparare. Parazitismul si comensalismul sunt fenomene interspecifice destul de rare intre speciile de pesti. Relatii extrem de complicate exista insa intre speciile de pesti pe baza nutritiei cu o hrana asemanatoare. Coincidenta spectrelor de nutritie survine la pestii adulti numai pe seama obiectelor trofice secundare pentru una dintre speciile de pesti si indeosebi atunci cand conditiile de mediu se modifica in asa fel incat una din speciile de pesti este fortata sa treaca la nutritia cu aceeasi hrana, ceea ce in conditiile unui deficit de hrana poate provoca ascutirea relatiilor trofice.
2.4. Relatiile interspecifice dintre pesti si alte organisme
Relatii foarte variate au loc in natura intre pesti si alte organisme (de la virusi si bacterii pana la om), acestea avand o importanta esentiala in viata pestilor (Nikolski, 1962).
a. Bacteriile si virusurile
Afara de importanta uriasa pe care o au bacteriile in viata pestilor ca si a altor organisme, prin participarea lor la circuitul substantei organice in natura, bacteriile au pentru pesti si o importanta speciala. O serie de boli grave ale pestilor sunt de origine bacteriana sau virotica.
Pe cat se pare, bacteriile prezinta importanta si ca hrana pentru pesti, cantitatea de bacterii inghitita o data cu materialul substratului este in partea anterioara a tubului digestiv mai mare decat in partea posterioara, ceea ce arata ca bacteriile sunt asimilate de catre pesti.
O deosebita importanta prezinta bacteriile ca hrana pentru nevertebratele care servesc ca hrana pentru pesti.
In apa marina costiera s-au evidentiat bacterii potential patogene pentru pesti din genurile: Actinomices, Nocardia, Flexibacter, Aeromonas, Pseudomonas si Vibrio. Aceste bacterii afecteaza pestii, in conditiile instalarii unor stari de stres, cauzate de factorii de mediu improprii, determinand infectii. Astfel s-au semnalat imbolnaviri infectioase la pestii bentonici (cambula, calcan si guvizi) cantonati in apropierea coastei in sezonul de primavara-vara.
b. Plantele
Dintre plantele inferioare, o importanta deosebita in viata pestilor o au ciupercile, indeosebi cele patogene. Acestea ataca icrele si pestii numai in conditiile slabirii organismului pestilor sau ale perturbarii metabolismului, fiind de cele mai multe ori un fenomen secundar.
O importanta enorma in viata pestilor o au plantele cu clorofila. Degajand la lumina oxigen si absorbind bioxidul de carbon, planta contribuie la crearea unor conditii mai favorabile pentru viata pestilor. In absenta luminii, atunci cand fotosinteza nu are loc, plantele degaja numai bioxid de carbon, acest proces putand ajunge uneori la asemenea proportii incat provoaca moartea pestilor.
Plantele au o mare importanta si ca substrat pentru icrele multor pesti. Depunand icrele pe plante, pestii le apara, pe de o parte, intr-o masura oarecare de pradatori, iar pe de alta parte, plaseaza icrele in conditii hidrochimice mai favorabile pentru dezvoltarea lor.
Multe specii de pesti sunt adaptate in mod special la viata pe pajistile submerse. Ele poseda un colorit protector corespunzator sau o forma speciala a corpului care le aseamana cu plantele printre care traiesc. Plantele (algele planctonice, epibioza, cat si macrofitele) au de asemenea importanta ca hrana pentru pesti.
Evolutia componentei micoplanctonice s-a caracterizat prin dominanta in structura populatiilor a grupului filamentos aflat intr-un procent mai ridicat in anii 1998-1999 decat in anii anteriori. In masele de apa tarmurale din spatiul predeltaic dar si inspre larg, in zona Constanta, s-au identificat in lunile aprilie si mai puseuri cu caracter de inflorire fungica.
Acumularile maxime de ciuperci in aceste luni au coincis cu concentratiile ridicate de substanta organica din masa apei, iar formele de masa au fost Penicillium chrysogenum (31,00%), Cladosporium algarum (22,6%), Candida diddensii (21,00%), Rhodotorula glutinis (17,22%).
Si in perioada de vara s-a evidentiat o bogata flora fungica, cu elemente consumatoare de substanta organica. Din punct de vedere cantitativ, productia totala de sporiti a fost de pana la trei ori mai mare fata de anii precedenti.
In perioada 1994-1999, in conditiile in care silicatii si fosfatii au fost in scadere comparativ cu anii anteriori, cantitatile totale de fitoplancton au fost scazute in raport cu cele din perioada de crestere a eutrofizarii, dar au ramas net superioare in raport cu cele din perioada anterioara intensificarii procesului respectiv (Rapoarte IRCM, 1994-1999).
Schimbari calitative si cantitative au avut loc in zona de coasta. Alga mare, bruna, perena, Cystoseira barbata, care nu tolereaza eutrofizarea, virtual a disparut. In anii 1970 a avut loc o scadere majora a biomasei de Cystoseira. In 1971, biomasa totala in lungul costei romanesti a fost estimata la 5.400 t; in 1972 ea a scazut la 900 t; in 1973 la 755 tone, in 1979 la 120 t si in 1980-1981, virtual a disparut.
Alte specii de macroalge continua sa se dezvolte intensiv de-a lungul granitelor eutrofizate, speciile de alge verzi, brune si rosii sunt mai bine adaptate la apele eutrofizate. Astfel, au aparut unele alge polisaprobice , care nu se gaseau in trecut in apa marii care nu era eutrofizata. Alga bruna Cladophora este un indicator al apelor saprobii
c. Crustaceii
In ceea ce priveste importanta lor in viata pestilor, crustaceii ocupa indiscutabil unul dintre primele locuri printre celelalte animale, mai cu seama ca organisme trofice.
Majoritatea speciilor de pesti atat rapitori, cat si pasnici (bentofagi, planctonofagi sau fitofagi), parcurg in dezvoltarea lor etape in care consuma crustacei planctonici, in primul rand din grupul Copepoda. Dar pestii se hranesc cu crustacei nu numai in stadiul larvar, crustaceii constituind baza nutritiei unor pesti planctonofagi adulti (exemplu: hamsia, sprotul, etc.).
Unii crustacei pot exercita o anumita influenta asupra bazei trofice a pestilor, consumand aceleasi organisme nutritive, iar altii provoaca daune ca pradatori, mai cu seama larvelor pestilor, atunci cand dezvoltarea prezinta o oarecare intarziere in raport cu fazele corespunzatoare ale dezvoltarii planctonului.
Pe pesti paraziteaza reprezentantii a trei ordine de crustacei: Copepoda, Brachiura si Isopoda. Toti crustaceii care paraziteaza pe pesti sunt paraziti externi ataca pielea sau branhiile pestilor. Crustaceii au un rol important in viata pestilor si ca vectori ai invaziilor helmintice, constituind gazde intermediare pentru unii viermi paraziti. In perioada 1988-1990, pe baza analizei unor parametri structurali inregistrati, se pot desprinde trasaturi ce au marcat evoutia populatiilor zooplanctonice in conditiile cresterii gradului de eutrofizare a mediului marin costier.
Comunitatile zooplanctonice au prezentat o situatie precara, cu o scadere evidenta a valorilor de biomasa trofica. Chiar in perioada de vara, cand in anii de dupa 1980, se consemna dezvoltarea uneori exuberanta a unor specii cu o frecventa mare in comunitatile costiere ca de exemplu Acartia clausi sau Pleopis polyphemoides, in vara ultimilor ani si aceste specii cu rol trofic pentru pestii planctonofagi, au fost in scadere.
In perioada 1991-1999, din Rapoartele IRCM, reiese ca specia Noctiluca scintillans, care provoca in lunile de vara ale deceniului anterior adevarate infloriri zooplanctonice, a avut acum dezvoltari moderate, diminuandu-si populatiile.
Biomasele zooplanctonului trofic inregistrate in vara anului 1999 au avut valori apropiate cu cele din lunile de vara ale ultimilor 2-3 ani, cand s-a evidentiat o usoara revenire a capacitatii trofice zooplanctonice in acest sezon. In timpul verii tendinta de scadere a valorilor biomasei trofice nu mai este atat de clar descrescatoare fata de primavara.
Comunitatile zooplanctonice identificate in spatiul marin din fata gurilor Dunarii au fost tot timpul verii mai ridicate, cantitatile fiind in scadere inspre sudul litoralului.
d. Viermii
Viermii au o importanta mare in viata pestilor marini. Foarte multi reprezentanti ai diferitelor grupuri de viermi servesc ca hrana pentru pesti; in bazinele marine un rol esential in hrana pestilor revine polichetelor.
Pe langa importantul rol pozitiv ce revine viermilor ca hrana, foarte mare este si influenta lor negativa, in primul rand ca paraziti ai pestilor (trematodele cestodele, nematodele, acantocefalii).
e. Molustele
Legaturile dintre pesti si moluste sunt foarte variate, ele putandu-se manifesta in relatia consumator-hrana (atat pestii se hranesc cu moluste, cat si molustele cu pesti), ca fenomene de parazitism si de comensalism, etc.
In apele marine, molustele constituie un obiect foarte important al nutritiei pestilor. Importanta trofica primordiala au pentru pesti molustele bentonice care constituie un procent insemnat in hrana principalelor specii demersale.
O serie de pesti folosesc molustele lamelibranhiate ca substrat pentru depunerea icrelor. Aceasta asigura protectia icrelor impotriva dusmanilor, precum si apararea lor de uscare in cazul variatiilor nivelului apei, deoarece cea mai mare parte dintre moluste reusesc in cazul scaderii nivelului sa se refugieze mai la adanc. Aceasta forma de legatura dintre pesti si moluste se incadreaza in notiunea de comensalism. Importanta molustelor ca paraziti ai pestilor este foarte mica. Pe pesti paraziteaza numai unele larve-glochidii, ale lamelibranhiatelor.
Domeniul bental reprezinta compartimentul vietii marine in care s-au produs in ultimii 20 de ani cele mai profunde modificari. Reducerea numarului de specii, saracirea calitativa progresiva reflectata de numarul tot mai mic de tipuri de organisme prezente in comunitati, reprezinta una din principalele modificari structurale. O alta caracteristica este aceea ca o serie de specii cu o frecventa mare in trecut au devenit rare, crescand in schimb frecventa speciilor tolerante.
Cronicizarea mortalitatilor in masa a organismelor de fund, consecinta a infloririlor tot mai frecvente, a condus la o saracire generala a fondului faunistic, reducerea indeosebi a populatiilor de bivalve avand drept consecinta diminuarea biofiltrului natural. Asa de exemplu, midiile ale caror stocuri prezentau in trecut chiar un interes economic de exploatare si aveau rolul lor de biofiltru in ecosistem, au pierdut 70% din biomasa ce o formau; in biocenoza midiilor unde se intalneau 49 specii in 1960, numai 11 specii macrobentale s-au intalnit in 1988 (Rapoarte IRCM).
Perpetuarea si accentuarea in timp a conditiilor nefavorabile a afectat in cele din urma chiar si populatiile speciilor tolerante care au inceput sa-si diminueze treptat efectivele, incat pe ansamblu, macrobentosul s-a redus de la an la an (de exemplu in 1987, densitatile si biomasele macrobentosului au fost cu 75% si respectiv 30% mai mici decat in 1985).
In perioada dintre 1973 si 1990, pierderile de vietuitoare au totalizat 60 milioane tone de animale de fund, incluzand 5000 tone de pesti (Zaitsev, 1992). Disparitia vechilor comunitati inseamna ca zone vaste cu apa de mica adancime au pierdut functiile lor anterioare de areale de hranire pentru puietul si adultii unor specii cum ar fi cei de calcan, cambula, sturioni, gobiide si barbun.
In ceea ce priveste tendintele manifestate in evolutia comunitatilor bentale, desi au indicat o usoara refacere in zonele sedimentare care in ultimii ani au fost puternic saracite ca urmare a mortalitatilor, aceasta refacere a fost mai mult realizata pe seama speciilor oportuniste.
In 1994 (Raport IRCM), s-a facut o evaluare a stocurilor de midii de mal din zona Constanta - Mangalia, intre izobatele de 20 - 50 m. Comparativ cu evaluarile anterioare, stocul din 1994 este de peste 2 ori mai mic decat in 1970, de 1,36 ori mai mic ca in 1981, dar de aproape 2 ori mai mare ca in 1982 cand au fost inregistrate mortalitati in masa ale organismelor bentale.
In perioada 1995-1999, tendintele pozitive manifestate raman inca putin semnificative, starea ecologica actuala fiind marcata in continuare de consecintele negative ale numerosilor ani anteriori, cu infloriri de mare amploare.
f. Celenterate
Ca obiect al nutritiei pestilor, celenteratele nu au mare importanta, intalnindu-se in hrana la putine specii de pesti.
Printre celenterate insa, exista o serie de specii precum meduzele si ctenoforele care provoaca mari daune resurselor de pesti. Ca obiecte nutritive pentru acestea servesc de cele mai multe ori icrele si larvele, dar sunt si specii care ataca pesti de dimensiuni mai mari. Pe de alta parte, la celenterate si pesti se intalnesc si exemple de simbioza, unii pesti traind sub clopotul unor meduze sau printre tentaculele unor anemone marine.
Unele celenterate, consumand aceeasi hrana ca si pestii, influenteaza asupra bazei trofice a acestora. Caz cu totul aparte, patrunderea accidentala in Marea Neagra a ctenoforului Mnemiopsis leidyi, semnalata in anul 1982, a cunoscut dezvoltari masive astfel incat dupa 1987 in perioadele de vara-toamna s-a ajuns la valori ale biomasei de 109tone pe intreaga mare. In unele zone de coasta abundenta sa a fost de 10-12 kg/m2, dar nu a depasit 1,5-3 kg/m2 in largul marii.
Concurent la hrana pestilor zooplanctonofagi si un vorace consumator de icre si larve de peste, acest organism este considerat factorul care a avut o influneta catastrofala asupra comunitatilor pelagice si asupra pestilor industriali planctonofagi.
Datele din literatura de specialitate privind biologia si fiziologia ctenoforului Mnemiopsis leidyi, precum si principalele modificari aparute in comunitatile pelagice odata cu dezvoltarea exploziva a sa au constituit elemente ce au determinat inceperea cercetarilor romanesti asupra distributiei si biomasei ctenoforului si in paralel cu acesta a meduzei Aurelia aurita.
Pe ansamblu, tendinta generala este de diminuare a densitatii aglomerarilor de ctenofor atat in sezonul de primavara, cat si in sezonul cald, in sezonul de primavara scazand de la 3,2 t/Mm2 in 1994, la 0,276 t/Mm2 in 1997, iar in sezonul cald, de la 10,0 t Mm2 in 1993 la 0,503 t/Mm2 in 1997.
Pentru meduza Aurelia aurita, pana in anul 1995, suprafata de raspandire este mult mai mica decat a ctenoforului Mnemiopsis leidyi, concentratiile maxime situandu-se in apropierea tarmului, acolo unde aglomerarile ctenoforului erau mai putin dense. Incepand din anul 1995, indeosebi in sezonul de primavara, suprafata de raspandire si densitatea pe unitatea de suprafata a meduzei au crescut ajungand la 40,4 t/Mm2 in anul 1998, fata de 1,79 t/Mm2 in anul 1994 (Radu si Maximov, 1996, Rapoarte INCDM, 1998-2006).
g. Pradatorii de varf, mamifere si pasari marine
In ultimele doua secole am asistat la decimarea unor populatii de pasari marine si mamifere care interactioneaza direct sau indirect cu recoltarea de catre om a produselor marine (Northridge, 1984). In anumite cazuri, de exemplu, mai multe populatii de foci s-au refacut odata cu introducerea legislatiei corespunzatoare si ajutate evident de regenerarea populatiilor speciilor mici ce constituie hrana lor. Fluctuatiilor naturale in abundenta coloniilor de pasari ce traiesc in apropierea stocurilor pelagice exploatate (Anon, 1995) se poate adauga si actiunea directa a oamenilor prin exploatarea speciilor pradate de ele.
Pradatorii de varf care includ mamiferele marine precum focile, delfinii si balenele si pestii mari ca rechinii, tonii, etc., joaca un rol deosebit in controlul speciilor pradate de ele, dar au fost printre primele specii care si-au redus numarul prin recoltare. Totusi, dat fiind complexitatea lanturilor trofice, eliminarea speciilor principale de pradatori de varf rezulta adesea din pescuitul altor organisme, suprapescuitul lor, sau prin schimbarea compozitiei hranei lor (Kerr, 1977; Ursin, 1982, Glasser, 1978).
Complexitatea lanturilor trofice face dificila cuantificarea impacturilor umane (May si al., 1979; Caddy si Sharp, 1986). Nu intotdeauna eliminarea unui pradator de varf a condus la sporiri majore in productia speciilor pradate de el (Pauly, 1979 a).
Pe de alta parte, daca luam in considerare marimea stocurilor speciilor pradatoare inainte de reducerea acestora constatam ca exista un grad de competitie intre om si pradatorii de varf pentru speciile ce constituie hrana lor comuna.
Astfel, o serie de calcule (Katona si Whitehead, 1988) au aratat ca populatiile de mamifere, chiar daca acum multe sunt reduse in marime, pot inca consuma din unele specii prada mai mult decat este recoltat de catre om.
Unele populatii de mamifere si pasari sunt vectori pentru parazitii ce infesteaza speciile comerciale de pesti (McClelland si al. 1983; FAO, 1978 b) si accidental pot cauza pagube uneltelor pescaresti. Aceasta explica partial comportamentul antagonist al pescarilor in unele parti ale lumii impotriva mamiferelor marine.
Calculele au fost facute prin prisma pierderilor pescuitului, insa nu este sigur ca astfel de calcule reprezinta efectele reale asupra pescuitului si nici ca ele pot fi aplicate simplist la estimarea productiei surplus ce ar putea fi realizata de om prin reducerea pescuitului natural al pradatorilor asupra pestilor. Astfel de calcule nu iau in seama legaturile ecologice, stabilitatea ecosistemului, rolul ecologic al mamiferelor marine, semnificatia lor pentru om datorita inteligentei lor, modalitatile de comunicare si comportamentul social.
Unul dintre obiectivele dezvoltarii durabile a stocurilor periclitate de pradatorii de varf este refacerea lor la marimi ce sa le asigure reproducerea fara pericolul colapsului populatiei. Totusi, marimi necontrolate ale populatiilor pradatorilor de varf nu sunt necesare fara repercursiuni adverse asupra disponibilitatii pestilor pentru hrana umana.
Starea actuala a celor trei specii de delfini din Marea Neagra (Delphinus delphis ponticus Barabasch - Nikiforov, 1935 - delfinul comun; Tursiops truncatus ponticus Barabasch - Nikiforov, 1940 - delfin cu bot gros, delfin mare, afalin; Phocoena phocoena relicta Abel, 1905 - marsuin, porc de mare, focena), nu este suficient cunoscuta, nu numai in apele romanesti, dar si in intregul bazin pontic, apartinand celor sase tari riverane (Romania, Bulgaria, Turcia, Georgia, Rusia si Ucraina), cu tot efortul depus de-a lungul anilor. Krotov (1949) si Zenkevich (1963) cred ca populatiile de delfini consumau aproximativ 500.000 tone de pesti anual.
O evaluare a stocurilor de delfini realizata in perioada 1983-1984, utilizand observatiile aeriene si navale, a estimat populatia totala la 60.000-100.000 exemplare (Vinogradov si Simionov, 1989). Observatiile realizate in nordul Marii Negre, in particular in nord-vestul Marii Negre, in perioada 1980-1990, au raportat o reducere de mai multe ori a populatiilor de delfini. Dupa ultimele estimari efectuate in anul 1987, in Marea Neagra, stocul de delfini a fost apreciat la 96 tone / 30 mii exemplare de Delphinus delphis ponticus, 7 tone / 3 mii exemplare de Tursiops truncatus ponticus si 10 tone / 3 mii exemplare de Phocoena phocoena relicta.
Datele despre foca din Marea Neagra lasa putin spatiu optimismului. Foca este usor deranjata si in primul rand a incetat sa apara in locurile aglomerate de oameni.
Pentru multi ani cea mai mare colonie de foci de Marea Neagra a fost localizata langa Capul Caliacra - Bulgaria. Doua exemplare de foca au fost observate la Trabzon in 1994, dar se crede ca aceasta specie nu are nici un viitor in Marea Neagra din cauza ca tarmurile sale sunt foarte populate.
Doi factori sunt critici in destinul populatiilor de foci. Primul este disponibilitatea locurilor adecvate pentru reproducere in zona tarmului si al doilea factor este hrana adecvata.
Influenta factorilor antropici gradul lor de influenta asupra resurselor marine vii
1. Determinarea factorilor antropici
Asa cum s-a mai aratat, pana nu demult, impactul omului asupra ecosistemelor marine a fost considerat in mare masura limitat la efectele pescuitului. Capacitatea mediului marin si a resurselor vii de a absorbi efectele altor activitatii umane a fost considerata efectiv nelimitata.
Cu timpul, s-a demonstrat ca schimbarile in conditiile de mediu ca rezultat al altor activitati umane (altele decat pescuitul) ce intra sub titulatura 'dezvoltarii' afecteaza de asemenea resursele vii. Acestea sunt:
activitati care au loc in interiorul sau adiacent la mediul marin precum dezvoltarea costiera, navigatia, deversarile in mare, turismul costier, mineritul submarin, extractia de gaze si petrol;
activitati umane din interiorul uscatului care afecteaza bazinul hidrografic marin si implicit sistemele marine prin intermediul raurilor (transportul materialului rezultat din eroziunea uscatului, nutrienti, pesticide, metale si alte materiale toxice) sau prin intermediul atmosferei (cloruri organice, metale grele, etc.).
a. Poluarea marina
Definitii variate ale poluarii marine trebuie sa fie luate in considerare in discutarea diverselor efecte induse de om asupra resurselor marine. GESAMP (1983) a definit poluarea ca: introducerea de catre om, direct sau indirect, de substante sau energie in mediul marin (inclusiv estuare), avand ca rezultat efecte dauntoare asupra resurselor vii, riscand sanatatea oamenilor, oprind activitatile marine (inclusiv pescuitul) si slabind calitatea pentru utilizare a apei marine.
In articolul 1 al Conventiei de la Londra privind Deversarile, partile contractante au pledat pentru luarea tuturor masurilor practice pentru a preveni poluarea marii prin aruncarea de deseuri si alte materii care sunt susceptibile de a crea riscuri asupra sanatatii umane, de a deteriora resursele vii si viata marina, de a interfera cu alte utilizari legitime ale marii.
Substantele chimice toxice, intre care enumeram metalele grele, produsele petroliere, detergentii, ajunse in apele marine litorale din apele Dunarii si din deversarile menajere si industriale din zona litorala au o mare influenta asupra sanatatii pestilor exploatabili. Metalele grele, indeosebi zincul si plumbul sunt foarte periculoase pentru pesti. La valori de peste 0,03 si respectiv 0,05 mg/l, aceste metale produc dereglari grave, ireversibile in functionarea organelor interne. Pesticidele si detergentii, in concentratii mari produc tulburari de metabolism, iar produsele petroliere determina
leziuni grave, indeosebi leziuni structurale ale organelor interne (ficat, rinichi, splina).
Poluarea chimica poate influenta productia de peste in numeroase feluri: reducerea stocurilor prin mortalitati in masa; declinul gradual sau schimbari in compozitia populatiilor sau a ecosistemelor intregi; sporirea existentei bolilor; deteriorarea calitatii alimentare a pestilor; micsorarea ratelor de crestere, etc.
Impactul deversarii deseurilor in mare, fie deliberat sau accidental, asupra resurselor marine, depinde intr-un grad semnificativ de localizarea si intensitatea deversarii. Astfel, deversarea petrolului in apele oceanice de larg este de asteptat a avea mai putine efecte serioase (desi nu necesar neglijabile) decat in apele din apropierea tarmului.
In multe medii marine, diversitatea si productivitatea sunt limitate de relativa uniformitate a topografiei fundului. In cazurile in care in astfel de zone cu fundul plat are loc aruncarea de materiale (dupa unele criterii convenite) pentru a forma recifi artificiali, pot fi in contradictie cu Conventia de la Londra, dar pot fi considerate benefice dezvoltarii resurselor marine si protejarii habitatelor marine impotriva traularilor. Astfel, este necesar a se discuta exact despre felul aruncarii, locul si intensitatea ei pentru a fi considerate daunatoare si daca conventiile existente trebuiesc modificate in scopul promovarii dezvoltarii durabile.
Se recunoaste ca:
mediul are o capacitate limitata de a gazdui deseuri fara a deveni semnificativ alterat de ele;
o astfel de capacitate poate fi cuantificata;
pana la un anumit nivel de contaminare nu se va produce un impact inacceptabil asupra resurselor marine vii.
Consumatorii au inceput sa devina tot mai constienti de calitatea produselor din peste si de necesitatea stringenta de introducere de catre guverne a reglementarilor pentru a asigura o astfel de calitate, lucru destul de dificil pentru tarile in curs de dezvoltare de a realiza un sandard minim din Codexul Alimentar FAO-WHO (FAO/WHO, 1993).
Deversarile in mediu si concentrarea ulterioara in lantul trofic a unor contaminanti precum: metalele grele, dioxinele, etc. pot avea impact asupra calitatii produselor din peste. Sunt in crestere in intreaga lume, problemele contaminarii produselor acvatice cu bacterii patogene si virusi continuti in apele de canalizare si deversarile domestice necontrolate din zonele urbane (Ahmed, 1991). Nutrientii din aceleasi surse favorizeaza de asemenea dezvoltarea exploziva a dino-flagelatelor toxice, contaminand pesti si scoici, indiferent daca sunt de cultura sau salbatice.
In zonele de exploatare a petrolului, capturile si uneltele de pescuit pot fi intinate cu petrol.
b. Constructiile costiere
Dat fiind faptul ca populatiile umane costiere sunt in crestere, constructiile au sporit in consecinta sub forma locuintelor, hotelurilor, porturilor, etc. si infrastructura asociata (statii de tratare a apelor de canalizare, rampe de gunoi, magazine, banci, oficii postale, etc.) si instalatii industriale. Asezarile umane aduc cu ele nu numai problema contaminarii, insa ameninta habitatele multor specii cu viata marina precum: broastele, mamiferele, pasarile si alte organisme care isi au habitatele in zonele costiere (in mare sau pe uscatul din apropierea marii).
Constructiile costiere au impact semnificativ asupra ecosistemelor marine locale si adesea forteaza pescariile de a-si schimba uneltele, metodele si speciile tinta.
c. Industria costiera
Multe industrii sunt stabilite in zonele costiere in principal datorita necesitatii unui loc de a arunca deseurile sau de a utiliza marea ca sursa de apa rece. Cele mai multe au de deversat deseuri chimice provenind din procesele de prelucrare (exemplu, componenti de mercur) si uneori deseuri fizice precum noroiul (de la prelucrarea minereurilor de aluminiu sau titan).
Uzinele de desalinizare prin definitie sunt situate la tarm, ele putand deversa saramura fierbinte. Industria costiera, nu numai ca poate afecta mediul marin local unde deverseaza, dar ea ocupa si mediul terestru din vecinatate pe seama altor habitate critice importante biologic pentru speciile marine si costiere.
Suma tuturor deversarilor industriale costiere, neabsorbite pe deplin de mare, au potentialul de a afecta advers pescariile marine costiere. Deci, industria costiera (inclusiv fermele pescaresti la scara industriala) au potentialul de a prejudicia interesele pescuitului costier si necesita o reconci-liere cu pescariile in contextul ICZM pentru a evita conflictul de interese.
d. Mineritul marin
Mineritul marin include forarea pentru petrol si gaze si dragarea pentru nisip si pietris. Cand astfel de minerit devine larg raspandit si in special daca unele prelucrari industriale sunt facute in mare, consecintele pentru pescarii pot deveni importante (descarcarea pulberilor de roca in mare poate creste turbiditatea sau sporesc concentratiile de elemente care in mod normal sunt rare in apa marii, dar care devin toxice in concentratii ridicate).
In prezent, extragerea de petrol si gaz pentru energie si prelucrare chimica este o activitate importanta, dar o data ce faza cercetarilor seismice, a forarilor exploratorii s-au incheiat si platformele de productie s-au amplasat, pescuitul este posibil afectat numai daca aceste platforme sunt amplasate prea aproape de zona de pescuit, sau in zona de pescuit, putand duce la inchiderea unei parti din aceasta. Totusi, astfel de parcuri de platforme pot constitui un refugiu pentru pesti si o oportunitate pentru pescuitul cu undita (Duglas si al., 1979; Reggio, 1987).
Accidentele de pe platforme duc la deversarea de cantitati substantiale de petrol in mare si au efecte deosebite asupra pescuitului stationar, dar si asupra altor activitati precum turismul.
Extragerea de pietris si nisip afecteaza negativ organismele bentonice prin distrugerea habitatelor, prin distrugerea zonelor de reproducere a pestilor demersali si interfereaza cu traularile si alte metode de pescuit. Totusi, durata efectelor, o data ce aceasta forma de minerit a incetat, poate fi relativ scurta.
e. Turism si recreere
Recreerea orasenilor si turistilor este o activitate economica importanta in multe tari. Oriunde sunt plaje curate, soare si infrastructura locala (hoteluri, locuri de divertisment, instalatii sanitare, banci, oficii postale, magazine, etc.) exista oameni carora le da mana sa vina aici si care platesc pentru aceasta. In multe zone litorale, in sezonul estival, populatia locala este multiplicata de mai multe ori. Aceasta cere investitii semnificative in infrastructura si servicii, chiar daca aceste facilitati vor fi folosite numai o parte din an. Autoritatile locale trebuie sa organizeze serviciile, indeosebi cele ce se ocupa de deseurile sporite din sezonul estival, pentru a evita poluarea mediului.
f. Acvacultura costiera
Acvacultura marina costiera constituie o solutie la criza de specii cu un pret ridicat si la cererea ridicata ce nu poate fi satisfacuta prin pescuit. Insa, hranirea intensiva a speciilor de cultura prin adaugarea de hrana in mediul natural, poate, in anumite conditii, sa duca la incarcarea anormala cu materie organica a marii costiere locale, conducand la eutrofizare (Enell si Ackefors, 1991), cauzand dezvoltarea exploziva daunatoare a planctonului (Whitei al., 1984).
Dezvoltarea acvaculturii marine nu poate fi vazuta favorabil de toate pescariile din cauza competitiei pentru piete, dar este de dorit a se asigura, unde este posibil, ca pescarii neangajati sau surplusul de forta de munca sa-si gaseasca un rol in acest sector.
g. Transportul naval costier
Rutele comerciale de transport naval nu sunt prea usor compatibile cu pescuitul industrial. Tancurile petroliere, pe langa faptul ca sunt foarte mari si greu manevrabile in apropierea tarmului, pot deversa din petrolul din tancuri, sporind astfel poluarea cu petrol ce poate fi nefavorabila, degradand mediul marin, intinand carnea pestilor, imbacsind uneltele de pescuit.
Deversarile majore de petrol pot constitui, din punct de vedere pescaresc, adevarate catastrofe locale, dar per ansamblu, riscul transportului naval a fost cel mai adesea exagerat.
h. Impactul agriculturii asupra bazinului hidrografic marin
Practicile agricole necorespunzatoare pun probleme serioase pescariilor costiere, atat prin pesticide, cat si prin pierderile de ingrasaminte.
Pesticidele, care sunt aplicate in cantitati excesive, pot fi spalate si duse direct in mare, sau indirect via rauri ce se varsa in mare si au efecte potential adverse asupra faunei marine costiere si asupra speciilor de interes. Exista de asemenea un transport substantial de pesticide prin atmosfera, ele putand ajunge in mare, dar pe o zona mai larga, afectand ecosistemele de la distanta.
Efectele ingrasamintelor chimice spalate de ploi direct in mare sau deversate via rauri, sunt potential foarte importante, adesea cu efect advers in vecinatatea punctelor sursa si posibil pozitiv mai departe dupa ce a avut loc dispersia.
i. Introducerea speciilor exotice si aspectul biodiversitatii
Numarul speciilor straine care au fost introduse deliberat sau accidental, in special in mediul marin, a crescut considerabil si este clar ca schimbarile din mediu datorate activitatilor umane pot facilita acest proces.
FAO a colaborat cu ICES in pregatirea unui cod de practici si manual de proceduri care sa stea la baza introducerii si transferului de organisme marine si de apa dulce (Turner, 1988).
Conservarea biodiversitatii si diversitatii genetice a organismelor acvatice poate fi influentata in doua moduri:
prin schimbarea mediului, suprapescuit sau competitia speciilor introduse;
prin canale facute de om pentru a lega doua bazine acvatice sau prin apa de balast din tancurile navelor de transport.
Schimbarile dramatice din fauna Marii Negre datorita introducerii de specii exotice mai bine adaptate la conditii de eutrofizare decat speciile bastinase (Ivanov si Beverton, 1985; Caddy si Grifiths, 1990), sunt un exemplu al influentei introducerii accidentale de noi specii.
2. Influenta pescuitului marin
Ihtiofauna marilor, in general si populatiile de interes comercial in mod particular au cunoscut in ultimele decenii mutatii severe si de cele mai multe ori imprevizibile. Cauza acestei situatii este pusa in principal pe seama degradarii accentuate a conditiilor de mediu in zonele litorale si indeosebi in aria selfului de nord-vest ce reprezenta principalul teren de reproducere si crestere a puietului speciilor comerciale.
Dependent de specificul biologic al fiecarei specii de pesti, aceste mutatii se manifesta atat in structura si potentialul ihtiofaunei, cat si in aspectele etologice ale populatiilor.
Factorii care au un caracter pur ecologic sunt completati pentru unele specii (sprot, hamsie, stavrid, calcan, sturion, rechin, etc.) cu un pescuit nereglementat si o captura necorespunzator de mare. Exploatarea excesiva a unor specii de pesti a dus si va duce la o scadere a biomasei acestora, iar prin folosirea unor unelte si tehnici de pescuit neadecvate, este periclitata starea rezervelor prin pescuirea puietului sau distrugerea comunitatilor bentale ce constituie hrana pentru multe specii.
Un factor care nu este in legatura directa cu eutrofizarea, dar are un efect similar asupra zonei bentice a Marii Negre, este practica moderna a traularilor de fund pe self (Zaitsev si al., 1992). Traularile comerciale cu traulul de fund in Marea Neagra,interzise din anii '70, au fost reluate, oficial s-au numit traulari in apropierea fundului, in scopul sporirii capturilor de sprot care se aglomereaza in straturile de fund ale selfului. Ca rezultat, in anii '80 capturile de sprot au fost de 15-20 de ori mai mari decat in anii '60 (Novikov si Serobaba, 1989).
Efectele cele mai drastice asupra fundului marii ale traularilor de fund, nu se datoreaza efectului mecanic asupra biocenozelor, ci mai ales, tulburarii apei care antreneaza mase mari de material sedimentar fin, asa-numita fractiune pelitica
Este usor de calculat ca la o deschidere de 25-50 m a unui traul ce este tractat cu o viteza de 3-4 noduri, intr-o ora de activitate, va tulbura malul pe o suprafata de cateva zeci de hectare. Deoarece exista mai mult de 200 traulere in Marea Neagra (Maximov, 2004) si fiecare nava lucreaza mai multe ore pe zi, este clar ca volumul de particule pelitice ce se ridica in coloana de apa este suficient pentru a avea un impact major asupra sistemului ecologic al Marii Negre. Aceste particule contin nu numai minerale, dar si substante organice care se adauga la eutrofizarea zonei, precum si substante toxice depozitate pe fund. Modul de miscare al particulelor pelitice care sunt rascolite de pe fund este catre sud, din zona vestica de pescuit si catre nord, din zona estica de pescuit.
Studiile geologice au aratat ca particulele antrenate de traule, in regiunile estice, sunt carate de curenti la o distanta de 150-200 km si depozitate pe o suprafata mai mare de 5.000 km2, la adancimi de 10-50 m. Grosimea stratului de sedimente variaza in unele zone de la 2-5 cm la 40-50 cm (Zaitsev si Mamaev, 1997).
Diversitatea specifica in zonele inmalite a scazut cu aproape 71%, abundenta cu aproximativ 60% si biomasa cu aproape 95%. Pierderile totale de pe o suprafata de 300 km2 de fund de mare in golful Karkinitsky, intens inmalit, s-au estimat la 800.000 tone de biomasa, in marea majoritate fiind midii. Aceasta reprezinta pierderea unei activitati de filtrare de 22-24 x109 m3 de apa de mare la fiecare 24 de ore (Kiseleva, 1979), precum si pierderea functiei anterioare ca zona de hranire pentru puietul si adultii unor specii de pesti.
In anii '80 rapitorii pelagici mari au disparut din capturile comerciale, ele fiind compuse in special din pesti de talie mica precum hamsia si sprotul (75-80%), bacaliarul si stavridul (Maximov, 2004). Majoritatea cercetarilor atribuie disparitia totala a pestilor pelagici mari din nord-vestul Marii Negre unei combinatii de factori, printre care:
vulnerabilitatea deosebita a acestora la activitatea de pescuit a navelor turcesti in timpul migratiei lor sezoniere prin Bosfor (Bosforul este un pasaj pentru fauna piscicola de apa calda care se reproduce si se hraneste in Marea Neagra si ierneaza in Marea Marmara si Egee);
poluarea industriala si domestica, traficul puternic si eroziunea costiera din Bosfor sunt alti factori care impiedica migratia anuala a pestilor intre cele doua mari.
Captura de peste din tarile riverane Marii Negre a crescut pana in anii '85-'88, dupa care a scazut drastic.
Observatiile efectuate in perioada 1990-1998 au evidentiat o completare putin numeroasa a rezervei de hamsie in toti anii din perioada 1990-1992. Aceasta situatie este legata de diminuarea considerabila a noii generatii de catre ctenoforul Mnemiopsis leidyi prin concurenta trofica si eliminare directa (perioada de reproducere a hamsiei suprapunandu-se cu perioada de dezvoltare maxima a ctenoforului)(Radu, 2004). In acelasi timp, datorita lipsei unei reglementari internationale a pescuitului, concentratiile de iernare ale hamsiei au continuat sa fie supuse unui pescuit intens.
Intr-o perioada critica pentru alte forme ihtiologice din Marea Neagra, numai sprotul a prezentat o fluctuatie naturala aproape normala si un efectiv relativ bun.
Pentru a demonstra efectul efortului de pescuit asupra starii resurselor pescaresti trebuie aratat ca la nivelul anului 2003, pe intreg bazinul Marii Negre, existau 4510 nave de pescuit, dintre care: 179 traulere; 223 seinere; 2580 driftere; 816 paragatiere (Maximov, 2004). Dupa cum se observa, din cele 4510 nave de pescuit, 4268 nave apartineau Turciei, aceasta detinand suprematia ca efort de pescuit la toate categoriile de nave si avand un efect aproape devastator asupra starii resurselor pescaresti, fiind compusa din trei categorii distincte: seinere; traulere si pescaria artizanala.
Plasele punga folosite de flota de pescuit turceasca sunt de aproximativ 1 km lungime si aproximativ 150 m inaltime (Anton E., 1992). Acestea captureaza pestele prin inconjurarea aglomerarilor, fundul apei sau stratul de H2S impiedicand evadarea pestelui in adancime. Navele care opereaza cu plasa punga sunt de obicei echipate cu sonare puternice, capabile sa determine cu precizie pozitia aglomerarilor. Dupa cum a fost consemnat de Gücü, 1997, ca fiind raportat de pescarii turci, la o singura lansare se pot pescui pana la 300 tone.
In pescuitul pasiv, desi efortul de pescuit a fost relativ constant, productia a scazut drastic. Structura pe specii a capturilor la taliene evidentiaza de-a lungul anilor o evolutie care reflecta in mare masura atat situatia stocurilor principalelor specii de pesti, cat si mutatiile in calitatea factorilor de mediu. Singura specie care a asigurat o oarecare constanta in capturi este sprotul, alaturi de care a aparut bacaliarul. Hamsia si stavridul au ajuns sa dispara practic din capturi pana la jumatatea acestui deceniu. In ultimii 10 ani efortul a scazut constant, ajungandu-se la o medie de 29-35 de taliene, amplasate de-a lungul intregului litoral, intre Portita si Vama Veche, fiind angrenati in aceasta activitate cca. 150-200 pescari, fata de peste 500 in anul 198 Captura rezultata din pescuitul cu talienele este livrata in stare proaspata sau depozitata si prelucrata (prin sarare) in cele 15 cherhanale de unde este expediata fie pentru consum uman in stare sarata sau uneori afumata, fie pentru consum animalier numai in stare proaspata.
In pescuitul activ, dominanta in capturi a asigurat-o sprotul cu peste 85%, urmat de bacaliar. Din cele 21 nave existente la mijlocul deceniului trecut (12 la I.P.Constanta si 9 la I.P.Sulina), numarul navelor a scazut dramatic, ajungandu-se la numai 7 nave active in anul 2000 si sub cinci nave, in ultimii ani. Daca la inceputul anilor '90, numarul de persoane direct implicate in activitatea de pescuit era cca. 180-200, acesta a scazut simtitor ajungand la circa 50 de persoane. Pestele este adus de pe nave la tarm sub forma sarata, o parte fiind expediat pe piata pentru consum in aceasta forma, iar o parte este prelucrat sub forma de conserve. Cantitati limitate se livreaza si sub forma proaspata.
In afara celor doua forme de pescuit, la litoralul romanesc se practica un pescuit recreativ, mai intens indeosebi in sezonul turistic, iar in ultimul deceniu un pescuit de subzistenta practicat de catre localnici. Acesta ultima forma de pescuit nu exercita o presiune prea mare asupra stocurilor de pesti atat timp cat se practica la un nivel artizanal.
Copyright © 2024 - Toate drepturile rezervate