|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Studiu ecologic si etologic al populatiilor genului Acrocephalus (Aves) din zonele umede ale Moldovei
Introducere.
Paseriformele de stuf sunt pasari a caror prezenta trece de multe ori neobservta. Prin modul lor de viata, prin hrana pe care o consuma, prin locurile unde isi stabilesc cuiburile, ele ofera o imagine elocventa si completa a starii zonelor umede.
Pentru monitorizarea starii de fapt a ecosistemlor umede este necesara studierea pasarilor de apa. Intre acestea se incadreaza passeriformele umede care constituie veriga cea mai importanta in desfasurarea ciclului de viata in ecosistemele acvatice, alaturi de unele pasari rapitoare. Ele se incadreaza in ultima treapta a piramidei trofice, cu importanta deosebita in elucidarea relatiilor vietuitoarelor din ecosistemle acvatice.
Studierea paseriformelor de apa este dificila din cauza dificultatilor ce le presupune identificarea lor. In lucrarea de fata s-a recurs la o determinare acustica in sezonul de reproducere, iar in afara acestuia, acolo unde a fost posibil la o determinare vizuala.
Detalierea aspectelor privind dinamica, situatia capturilor, biometria, structura cantecului au fost facute cu credinta ca ele vor constitui date care sa reprezinte baza unor planuri de dezvoltare durabila.
Dinamica pasarilor cuprinde totalitatea modificarilor care se petrec in efectivele numerice, in biometria si comportamentul lor. De aceea, am considerat necesar sa surprindem mai multe aspecte ecologice privind distributia, morfometria si cantecul, ce reprezinta baza unei cunoasteri detaliate a paserifomelor stuficole.
Studierea comportamentului la pasari are o mare importanta stiintifica si practica, intrucat prin aceasta se pot gasi noi modalitati de protectie a faunei si florei, de identificare mai judicioasa a speciilor si sexelor la speciile monomorfice, asa cum este cazul genului Acrocephalus. Lacarii (genul Acrocephalus) se situeaza la capatul lantului trofic, intrucat sunt specii insectivore. De acea ei au importanta deosebita in cadrul ecosistemelor acvatice. Cu cat numarul si rata lor de supravietuire este mai mare in zonele umede cu atat este mai sanatos mediul respectiv. Pentru a gasi mijloacele de protectie, trebuie urmarite aspecte ale comportamentului. Astfel oamenii pot sa intervina in ecosistemele lor naturale, pentru a le proteja, numai la momentul oportun.
In decursul iesirilor in teren am incercat sa stabilim un inventar al actiunilor comportamentale a speciilor de lacari, pentru a alcatui o etograma care sa ne permite interpretarile privitoare la comportament.
In lucrarea de fata au fost surprinse aspecte indeosebi ale etologiei lacarului mare- Acrocephalus arundinaceus, de la migratia de primavara pana in cea de toamna. Comportamentul lacarului mare a fost comparat cu cel al lacarului de stuf- A. scirpaceus si cu cel al lacarului mic- A. schoenobaenus, la care s-au facut numeroase referiri..
Rezultate si discutii
Habitatele ocupate de paseriformele de stuf, in Moldova, sunt in cea mai mare parte omogene din punct de vedere al vegetatiei si influentei antropice. Unele zone, insa in ultimii 20 de ani au fost modificate mai ales prin amenajari hidroameliorative. Datorita acestui fapt, au aparut si modificari majore in zonele invecinate lacurilor sau cursurilor de apa. Un alt factor care a dus la alterarea habitatelor naturale umede in Moldova este practicarea agriculturii intensive. Aceasta a dus la disparitia unor habitate naturale din vecinatatea luciului de apa si la instalarea unor habitate antropogene dominate de culturi agricole uniforme. Cu toate acestea paseriformele de stuf si in special lacarii (genul Acrocephalus) s-au adaptat foarte bine la conditiile nou create, fiind speciile dominante de passeriforme in aceste ecosisteme acvatice.
Date fiind considerentele de mai sus am recurs la o descriere a habitatelor din zonele umede investigate de noi pana in prezent. Zonele investigate, considerate stationare, sunt: elesteele Larga Jijia- Vladeni (judetul Iasi); lacul Balta Lata; Lacul Iezar (judetul Botosani); elestele Carja (judetul Vaslui); elestele Vladesti (judetul Galati); lacul Beresti (judetul Bacau). Mentionam ca incursiuni au avut loc si pe lacul Lilieci, amenajat pe cursul raului Bistrita, la 10 km N de municipiul Bacau, dar datele adunate sunt neconcludente, de aceea nu am recurs la o descriere a zonei.
Observatiile s-au desfasurat pe parcursul a doi ani de zile. Datele din zonele investigate au fost comparate tinand seama de perioadele de observatii pentru fiecare zona, pozitia geografica, aspecte particulare privind fizionomia fiecarui habitat.
In ceea ce priveste habitatul natural al paseriformelor de stuf el este constituit de un luciu de apa invadat partial de stuf inconjurat de pajisti iar la distante de 20- 100 m sunt culturi agricole. Aproape toate zonele umede investigate de noi cu exceptia Iezarului Dorohoi se afla in vecinatatea unor paduri de foioase (la distante de peste 500 de m fiecare). Cadrul general al habitatului natural al paseriformelor de stuf se contureaza dupa cum urmeaza:
In jurul baltilor, ca si in interiorul lor, exista o vegetatie variata si abundenta, favorizata si de belsugul apelor. (Popovici, 1994). Cadrul baltii il formeaza stuful (Phragmites comunis) sau trestia (Typha angustifolia), rogozul (Carex riparia), stanjeneii de balta (Iris pseudacorus). In interiorul baltii gasim plante amfibii sau emerse: buzduganul de apa- Sparganium ramosum, sageata apei- Sagittaria sagittifolia, limbarita- Alisma plantago, cucuta de apa- Cucuta virosa, rachitanul- Lythrum salicaria. In apa sau la suprafata ei exista exista plante submerse ca: nufarul alb (Nymphea alba) si nufarul galben (Nuphar luteum). Dintre plantele plutitoare fixate prin radacini sunt mai frecvente: broscarita- Potamogeton natans, troscutul- Polygonum amphibium, cornaciul- Trapa natans. Pe suprafata apei plutesc plante nefixate prin radacini: pestisoara- Salvinia natans, otratelul de balta- Utricularia vulgaris, lintita- Lemna trisulca, iarba broastelor- Hydrocharis morsus ranae.
In apropierea apelor, in locurile de unde paserifomele de apa isi procura cea mai mare parte din hrana se gaseste o vegetatie tipica de stepa. In complexul vegetatiei de stepa (Popovici, 1994) din Moldova o mare raspandire o au gramineele: firuta- Poa bulbosa, obsiga- Bromus inermis, orzul soricesc- Hordeum murinum, paiusul- Festuca pseudovina- colilia- Stipa pennata, barboasa- Antropogon ischaemum, din familia Graminee. Din familia Compositae amintim: pelinita- Artemisia austriaca, cicoarea- Cichorium inthybus. Alaturi de aceste specii de plante in pajistile de la marginea apelor mai exista: salvia de camp- Salvia pratensis, mazarichea- Vicia stenophylla, - Coronilla varia, cosaci- Astragalus cicer, plevaita- Xeranthemum annum, ciulinul- Carduus natans, corovaticul- Verbascum thapsus. Pe langa plantele ierbacee se mai gasesc tufisuri si maracinisuri formate din porumbar- Prunus spinosa, visinel- Prunus fruticosa. Portiunile inundabile de la marginea baltilor sunt favorabile zavoaielor formate din: salcii, rachite si plopi.
Vegtatia palustra si cea de stepa este favorabila existentei speciilor de nevertebrate cu care se hranesc paseriformele de apa. In apa si in ariile inmlastinite se gasesc melci. In stufarii se gasesc efemeroptere, diptere, atat adulti cat si larve (Cramp, 1992), hemiptere, lepidoptere, trichoptere (Ion C., 2004) iar in vegetatia ierboasa se gasesc pe langa nevertebrate mentionate mai sus: paianjeni, coleoptere, precum si diverse materiale vegetale. Speciile de plante cu care hranesc paseriformele de apa nu au putut fi precizate in analizele de pana in prezent. (Cramp, 1992).
In conditiile existentei unui astfel de cadru natural traiesc paseriformele de apa cu mentiunea ca: stufarisurile dinspre maluri si terenurile invecinate sunt ocupate in special de paseriforme de stuf, pe cand in cele situate spre interiorul suprafetei acvatice sunt dominate de pasari de apa de talie mare.
In ceea ce priveste distributia speciilor analizate s-a observat ca aceasta se refera la modul de ocupare a unui microhabitat de pe intreaga suprafata a unei zone umede pentru satisfacerea cerintelor de cuibarit, cautare de hrana si adapost.
Exemplarele de pasari ce apartin speciilor de Acrocephalus arundinaceus (figura 18) si Acrocephalus scirpaceus (figura 19) au fost capturate in toate stationarele de proba. S-a observat ca Acrocepahlus scirpaceus are o distributie aglomerata in zonele umede investigate. Aceasta pasare isi stabileste locuri de popas sau de cuibarit in benzile de stuf, fara a circula pe distante mari de la un cordon de stufaris la altul in cautarea unui teritoriu. In zonele unde stuful ocupa suprafete restranse (lacul Beresti, Iezar), Acrocephalus arundinaceus are o raspandire grupata. Acest lucru se explica prin concurenta pentru spatiu cu alte specii de pasari acvatice, care sunt prezente intr-un numar mare de exemplare in zonele respective. In ecosistemele acvatice cu stufarisuri intinse (Larga Jijia, Carja, Vladesti), aceasta specie are o repartitie uniforma in stufarisuri, urmare a faptului ca suprafata unde isi desfasoara activitatea este mult mai mare si "interactiunea" cu alte specii mult mai scazuta. In zona lacului Balta Lata, lacarul mare- Acrocephalus arundinaceus are o dispersie intamplatoare (Poisson). Datele obtinute prin calcularea indicelui de agregare concorda cu observatiile din teren in sensul ca Acrocephalus arundinaceus apare intr-un numar mai mare in raport cu celelalte specii de lacari. Numarul exemplarelor de lacar mare- Acrocephalus arundinaceus este ridicat inaintea sezonului de reproducere in zona Balta Lata, dupa care numarul lor descreste. In perioadele de vara si in cea de migratie de toamna numarul de exemplare a acestor pasari este in scadere.
Distributia pe verticala se refera la spatiile de microhabitat preferate de paseriformele de stuf, in functie de inaltimea vegetatiei existente si a miscarilor pe care le executa.
In deplasarile facute in teren s-a urmarit si dispersia pe verticala a paseriformelor de stuf.
Lacarul mare (Acrocephalus arundinaceus) are un comportament mai "indraznet" decat al celorlate specii in sensul ca poate fi vazut adesea cantand pe varful firelor de stuf. De asemenea, in deplasarile sale in interiorul stufarisurilor se catara cel mai adesea in treimea superioara a firelor de stuf, iar prezenta sa catre varful firelor de stuf poate fi pusa atat pe seama taliei sale mai mari, in comparatie cu ceilalti lacari, dar si a explorarii unui spatiu cat mai larg pentru prinderea insectelor.
Lacarul mic (Acrocephalus schoenobaenus) executa miscari de mica amploare in regiunea mediana a firelor de stuf. Este o specie mai "rezervata". Miscarile sale se concentreaza in interiorul stufariilor si sunt executate pe distante scurte.
Lacarul de stuf (Acrocephalus scirpaceus) are un comportament mai tolerant, adesea acceptand apropierea omului la distante destul de mici. Isi cauta hrana sau se ascunde, mai ales, la baza stufului. In momentele cand observa apropierea unui eventual pericol se prinde de varful firelor de stuf. In perioada de cuibarit, comportamentul sau in interiorul stufarisurilor este asemanator lacarului mic, adica face deplasari de mica amplitudine. Prezenta sa se concentreaza in zonele cu stufarii compacte la 1-1,5 m de sol.
In cadrul studiului nostru asupra avifaunei paseriformelor acvatice din unele zone umede ale Moldovei, am incercat sa surprindem dinamica compozitiei specifice a comunitatilor de pasari de-a lungul aspectelor ecologice. Pasarile sunt cunoscute ca animale extrem de mobile care se deplaseaza dintr-un habitat in altul, in cautarea hranei si a conditiilor propice realizarii procesului de reproducere. Motivul acestor deplasari este dublu: 1. sporirea sanselor de supravetuire a acestor pasari care parasesc ariile lor de hranire inainte ca sezonul nefavorabil sa vina; 2. permite pasarilor sa utilizeze temporar resursele de hrana disponibile, asa cum ar fi semintele existente din abundenta in anumite sezoane, pesti, insecte, ceea ce sporeste numarul juvenililor care pot supravetui. (Gache Carmen, 2002, a)
Beneficiile migratiei trebuie privite prin prisma costului unui zbor extenuant, in timpul caruia pasarile pot muri din cauze variate: epuizarea energiei, vremea nefavorabila, pierderea simtului de orientare s.a. Acest fapt confera termenului de migratie beneficii si costuri individuale si nu reprezinta doar un simplu fenomen populational.
Pe parcursul unui an putem delimita doua perioade de pasaj: pasajul de primavara care se desfasoara pe parcursul lunilor martie, aprilie si mai si pasajul de toamna ce tine de la sfarsitul lunii iulie si pana la sfarsitul lunii octombrie.
Speciile genului Acrocephalus sunt oaspeti de vara. (Munteanu D, 1999).
Studiul nostru a urmarit dinamica paseriformelor de stuf punand accentul, mai ales, pe speciile genului Acrocephalus, care reprezinta prezenta cea mai comuna..
Studiul dinamicii paseriformelor de stuf s-a realizat pe trei zone umede: Iezar (nordul Moldovei), Larga Jijia(zona central-estica a Moldovei) si Beresti (zona de vest Pentru a avea o imagine completa a dinamicii anuale a efectivelor paseriformelor de stuf am recurs la ilustrarea grafica a efectivului mediu. Metodele folosite in determinarea efectivului, au fost transectele vizuale si auditive pe distante insumand 1km de-a lungul benzilor de stuf. In zonele marginite de dealuri (Iezar) si diguri (Beresti) s-a folosit si metoda observatiei din punct fix. (Metode de evaluare a abundentei pasarilor, 2000). In cazul transectelor vizuale randomizate si a celor din punct fix am folosit binoclu 10x50 si luneta 40x60. Itinerariile s-au efectuat intre orele 6- 9 dimineata sau intre orele sase si noua seara in sezonul prereproductiv si reproductiv. In sezonul postreproductiv cantecele speciilor de passeriforme acvatice sunt razlete sau chiar absente. Pentru a determina efectivul mediu al pasarilor investigate inaintea migratiei de toamna s-a procedat la identificarea vizuala, acolo unde acest criteriu este posibil si unanim acceptat de ornitologi. Pentru a nu fi confundate speciile genului Acrocephalus cu alte specii ale aceluiasi gen, semnalate in Romania, dar inca nesemnalate in Moldova s-a realizat si studierea sunetelor scurte emise de aceste pasari care au fost, in cele mai multe cazuri, edificatoare in determinarea lor. Aceasta metoda a fost utilizata, intrucat timpul avut la dispozitie a fost scurt si pentru ca determinarea acustica este mult mai exacta decat cea vizuala. (Svensson, 1978). Determinarea cantecelor emis de lacari (genul Acrocephalus) dupa sunetele emise a fost posibila dupa un "antrament" realizat pe baza audierii unor cantece inscriptionate pe Cd-uri de specialisti ornitologi din strainatate.
Dinamica sezoniera a paseriformelor de stuf in cele trei zone umede luate in studiu, inregistreaza mari variatii numerice in stationare din regiunile Larga Jijia si Iezar. Aici se inregistreaza un maxim numeric in aspectul prevernal corespunzator migratiei de primavara, pentru ca apoi numarul lor sa scada in celelalte sezoane. Explicatia rezida in faptul ca in primavara, inaintea perioadei de reproducere, exista un numar mare de insecte, componenta principala a hranei acestor pasari. Ca urmare, in zonele umede ale Moldovei se aduna un numar mare de indivizi ce apartin paseriformelor de stuf care-si fac rezerve energetice, ce sunt necesare in perioada cuibaritului.
Ca efect al concurentei pentru hrana ori a fidelitatii locului de cuibarit (Hasselquist, 1998) unele paseriforme de stuf migreaza mai catre nord, iar altele raman pe loc pentru cuibarit. Daca teritorialitatea devine accentuata atunci unele pasari se intorc in ecosistemele acvatice mari din sud- est, in Delta Dunarii.
Observatiile noastre arata ca toamna, in zonele din nordul si nord-estul Moldovei raman mai ales juvenilii si mai putini adultii de lacar mare sau lacar mic. Din aceasta cauza numarul de paseriforme de stuf este relativ mic. In timpul verii s-a inregistrat seceta prelungita ce a avut ca efect retragerea apelor (2003), si aridizarea terenurilor. Insectele au fost mai putine si active doar seara si mai putin dimineata. O alta cauza a prezentei acestei speciilor intr-un numar scazut toamna o reprezinta si conditiile de mediu, cel mai adesea nefavorabile. La sfarsitul verii, inceputul toamnei se inregistreaza adesea ploi torentiale si vanturi puternice, scara 4-5 Beaufort (asa cum a fost in anul 2003).
In zona umeda a lacului Beresti, paseriformele de stuf inregistreaza un numar relativ constant de-a lungul celor trei sezoane luate in calcul. Diversitatea habitatelor, impactul antropic relativ scazut, existenta hranei asigura conditii pentru existenta acestor pasari. Dinamica lor numerica in aceasta zona nu inregistreaza variatii mari. Primavara, suprafetele restranse de stufarii nu ofera posibilitati pentru popasul indelungat a unui numar crescut de exemplare si apare o concurenta interspecifica a paseriformelor de stuf, fie o concurenta cu pasari de apa ce exploateaza aceleasi nise ecologice de hranire si reproducere. Teritorialitatea este accentuata vara, astfel incat inclinam sa credem ca se respecta regula "primul venit cuibareste". Toamna, adultii pleaca mai devreme, iar juvenilii nu pornesc in calatoria lor spre cartierele de iernare pana nu acumuleaza un strat suficient de grasime care sa le asigure combustibilul pentru calatoria spre sud. (vezi Fig. 1)
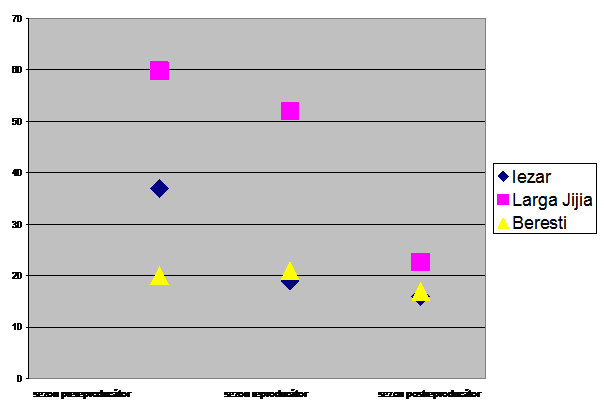
Fig. 1: Dinamica sezoniera a paseriformelor de stuf in zonele umede din Moldova: Iezar, Larga Jijia, Beresti.
In cele ce urmeaza am recurs la o analiza a comunitatilor paseriformelor de stuf, a relatiilor intraspecifice si interspecifice ce se stabilesc si raportul relatiilor intre diversele specii si mediul in care traiesc. Analiza complexa a paserifomelor ne permite aprecierea starii generale a habitatelor, si ofera indicii valoroase in vederea valorificarii durabile a resurselor din zonele umede atat pentru viata animalelor cat si pentru om.
Analiza sinecologica ne poate oferi o imagine despre relatiile ce se stabilesc intre diversele comunitati de specii si ierarhiile ce se stabilesc intre diferitele specii. Aceasta analiza ne permite sa vedem care este importanta unor specii intr-un ecosistem sub aspectul schimburilor energetice cu mediul, care sunt speciile caracteristice unui biotop, care sunt speciile ce au ajuns intamplator in zonele cercetate si in acest fel putem stabili interrelatiile intre diferitele specii care alcatuiesc biocenoza.
Datele acumulate, in urma observatiilor efectuate, au permis analiza sinecologica, prin calcularea unor indici ecologici analitici: abundenta, constanta si dominanta si a unor indicatori ecologici sintetici, cum ar fi: indicele de semnificatie ecologica si indicele de afinitatea cenotica.
Caracterizarea speciilor in cadrul comunitatilor paseriformelor de apa se poate face in urma analizei autecologice si sinecologice iar investigarea gradului de organizare a comunitatilor biocenotice se poate aprecia prin studiul diversitatii speciilor.
Abundenta propriu-zisa reprezinta numarul total al indivizilor unei specii dintr-o anumita zona si se exprima in valoare absoluta. (Gomoiu., 2001). Pentru aprecierea abundentei paseriformelor din zonele umede s-a folosit metoda transectelor auditive, pe fasii lungi de 1 km cu o latime maxima de 40 m. Intodeauna numaratoarea exemplarelor observate a fost facuta spre interiorul suprafetei de proba. Latimea maxima a transectelor executate au fost conditionate de distanta maxima de recunoastere dupa cantec a paseriformelor de stuf dar si de latimea suprafetelor de stuf, ce depaseste rareori 40 m. Transectele au fost parcurse randomizat in functie de configuratia terenurilor (de modul de dispunere a malurilor sau a digurilor ce separa lacurile intre ele), pentru a acoperi o varietate cat mai crescuta de microhabitate din cadrul ecosistemelor umede investigate. Observatiile au fost efectuate intre orele 6-9 dimineata si 18-21, seara. Pentru identificarea speciilor de pasari, mai ales in cazul lacarilor, am folosit metoda determinarii acustice. Avand in vedere ca numai masculul canta (Cramp, 1992), rezultatele obtinute s-au estimat sub forma numarului de perechi de clocitoare. (The EBCC Atlas of European Breeding Birds, 1997).
Estimarea efectivului, care este abundenta propriu-zisa, numerica, a fost realizata in perioada de cuibarit, in lunile mai , iunie, iulie. In etapele premergatoare acestei perioade sau cele care se succed acesteia se inregistreaza mari fluctuatii ale numarului de indivizi. Fluctuatiile numerice a paseriformelor in perioadele prereproductive apar pentru ca la indivizii clocitori se adauga exemplare aflate in pasaj. Dupa terminarea perioadei de reproducere la populatia clocitoare de pasari se adauga si puii zburatori. Rezultatele obtinute pe transectele efectuate au fost estimate pentru o suprafata standard de 10 ha asa cum sugereaza The EBCC Atlas of European Breeding Birds, 1997.
Pentru a reliefa pozitia lacarilor (genul Acrocephalus) in cadrul ecosistemului am recurs la estimarea parametrilor ecologice pentru toate paseriformele acvatice inventariate.
Pe baza probelor prelevate s-a recurs la calculul efectivului de paseriforme de stuf pentru toate zonele si pentru fiecare specie in parte, conturandu-se o imagine de ansamblu asupra situatiei actuale acestor pasari. Datele acumulate ne-au permis a efectua o analiza statistica astfel ca in tabelul 1, am calculat separat pentru cele sapte specii de paseriforme studiate din cele sase zone umede o serie de parametri statistici: media aritmetifca, eroarea standard a mediei (Er. std.), limita superioara (LS), limita inferioara (LI) a efectivelor speciilor. Intervalul limita inferioara si limita superioara contine media populatiei investigate din care a fost extrasa proba, cu o probabilitate de 95% (α 0,050, adica posibilitatea de a fi facut o eroare este de 5%). In unele cazuri au rezultat mari amplitudini ale intervalului de confidenta limita superioara, limita inferioara (LS- LI), pe baza probelor prelevate. In acest caz au rezultat limite inferioare negative care nu au fost prelucrate ulterior, nefiind reale. Ele s-au datorat, probelor inegale ca marime si probabil, inca insuficiente. Existenta acestor deficiente in estimarea mai corecta a populatiilor s-a datorat in principal diferentelor in ceea ce priveste configuratia terenului, marimea habitatatelor studiate.
Tabel 1: Efectivul estimat pe baza observatiilor efectuate la paseriformele de stuf din unele zone umede ale Moldovei in perioada de cuibarit.
|
Statistica descriptiva |
Specia |
|||||||
|
Parametrii |
Acr aru |
Acr sci |
Acr sch |
Loc lus. |
Pan bia |
Rem pen |
Emb sch |
|
|
Larga Jijia |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Balta Lata |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Vladesti |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Beresti |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Iezar |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Carja |
Media | |||||||
|
Nivel de confidenta | ||||||||
|
LS | ||||||||
|
LI | ||||||||
|
Total |
Media | |||||||
|
LS | ||||||||
|
LI | ||||||||
Locustella fluviatilis semnalata de noi in zonele umede investigate nu a fost cert inventariata ca fiind cuibaritoare de aceea nu a fost inclusa in tabelul de mai sus.
In lucrarea de fata am folosit indicele Bray Curtis pentru a grupa speciile in functie de valorile efectivului mediu, intrucat acest indice tine seama nu numai de prezenta sau absenta speciilor, dar si de numarul de indivizi a fiecarei specii. Avand in vedere ca indicele Bray Curtis este un indice de disimilaritate (FIg. 2) si ca similaritatea este data de 1 − similaritatea, speciile Emberiza schoeniclus si Acrocephalus scirpaceus prezinta o similitudine (corelatie) crescuta in ceea ce priveste efectivele medii in zonele cu stufaris. Cele doua specii au fost semnalate intr-un numar relativ constant de-a lungul investigatiilor noastre. Ambele specii prefera intinderile de stuf compacte pe care rareori le parasesc pentru a-si procura hrana. Ele se deplaseaza mai rar pe distante lungi.
Panurus biarmicus si Remiz pendulinus sunt grupate pe clustere (grupuri) apropiate in ceea ce priveste efectivele. Cele doua specii se gasesc acolo unde pe langa stufarisuri se gasesc salcii, arbori caracteristici zonelor umede, pajisti intinse, locuri de unde isi procura hrana. Numarul exemplarelor celor doua specii se afla intr-un usor echilibru. Panurus biarmicus inregistreaza efective mai crescute iar Remiz pendulinus un numar de indivizi mai scazut. Cand unul din efective creste celalalt scade sau nu este reprezentat prin nici un individ si invers. Speciile de pitigoi de stuf (Panurus biarmicus) si laicus Remiz pendulinus), se intalnesc mai putin in acele zone umede marginite de campuri agricole sau unde impactul uman este crescut.
Intre speciile genului Acrocephalus (A. arundinaceus si A. schoenobaenus) se observa similitudine relativ crescuta. Lacarul mare (A. arundinaceus) se intalneste in numar mare in toate ariile umede investigate. Lacarul mic (A. sch.) se observa in zonele umede mai ales in momentele cand, activitatea lacarilor mari este mai putin intensa sau cand efectivele sale se concentreaza in alte zone.
Locustella luscinioides- greluseul de stuf, se semnaleaza rar in stufarisurile din zonele umede ale Moldovei de aceea similitudinea in privinta efectivelor inregistrate de aceasta specie este slaba. (Fig. 2).
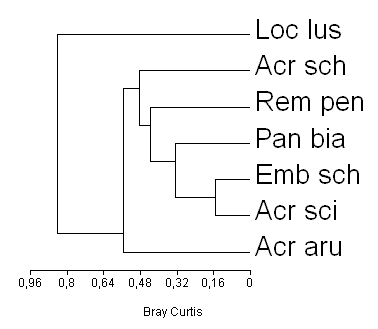
Fig. 2: Gruparea speciilor in functie de valorile efectivului mediu in perioada de reproducere
Pentru a observa corelatiile si asemanarile ce se stabilesc intre paseriformele de stuf si intre caracteristicile zonelor umede in ceea ce priveste prezenta pasarilor de apa am realizat cu ajutorul unui program de specialitate (XLstat) analiza corespondentei intre preferintele de mediu ale speciilor analizate si zonele in care au fost gasite. Astfel, in urma efectuarii aprecierii privind efectivul, datele au fost transpuse in fig. 3. In acest grafic se ilustreaza abundenta fiecarei specii pe fiecare zona umeda iar fiecare zona este analizata din punct de vedere al abundetei fiecarei specii de paseriform de stuf.
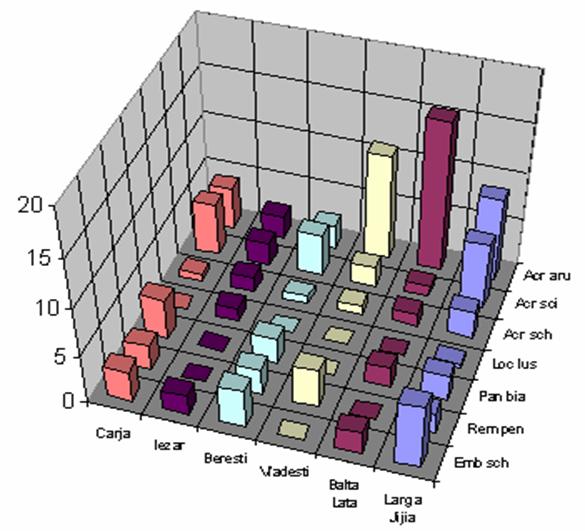
Fig. 3: Efectivul speciilor de paseriforme de stuf in unele zone umede ale Moldovei
Cele mai mari efective inregistrate de noi (vezi grafic 9) au fost in cazul speciei Acrocephalus arundinaceus. In zonele umede aflate pe valea Prutului (Larga Jijia, Balta Lata, Vladesti, Carja) se inregistreaza un numar crescut de exemplare. Acrocephalus scirpaceus si Emberiza schoeniclus, prezinta efective crescute si chiar egale, in zonele Larga Jijia, Beresti si Carja. Pentru Remiz pendulinus si Panurus biarmicus se observa o distributie echilibrata a efectivelor de-a lungul habitatelor analizate. Lacarul mic (Acrocephalus schoenobaenus) este prezent ca specie cuibaritoare in toate habitatele analizate, insa in numar mic. Locustella luscinioides prezinta un numar mic de perechi clocitoare, 5 perechi, in nordul Moldovei, la Iezar.
Avand in vedere ca nodurile dendrogramei (Fig. 2) sunt pivotante, se pot roti, am incercat sa aflam in ce masura speciile grupate in dendrograma sunt reprezentate de-a lungul habitatelor analizate si daca exista vreo corespondeta intre caracteristicile habitatelor analizate si prezenta paseriformelor de stuf. Pentru aceasta am recurs la construirea graficului bidimensional Bray- Curtis.
Axa 1 reprezinta variabilitatea maxima, iar Axa 2 reprezinta variabilitatea cea mai mica. Acrocephalus arundinaceus prezinta o mare variabilitate a efectivelor in zonele analizate. Acrocephalus scirpaceus, Emberiza schoeniclus si Panurus biarmicus prezinta o mare variatie numerica a efectivelor. Aceste specii sunt prezente inconstant in habitatele analizate : Panurus biarmicus lipseste in zonele Vladesti si Iezar, Emberiza schoeniclus este absenta in zona Vladesti, iar toate cele trei specii mentionate anterior prezinta mari variatii ale efectivului lor in habitatele analizate datorita diferentelor in rezervele de natura trofica si datorita configuratiei diferite a ecosistemelor analizate. Acrocephalus scirpaceus este prezent in numar mare numai in acele zone unde stufariile sunt intinse (Larga Jijia, Beresti, Carja- si resursele trofice foarte bogate ; Panurus biarmicus se gaseste in zonele in care Remiz pendulinus este aproape absent. Cele doua specii intra in concurenta pentru hrana mai ales in perioada prereproductiva si in perioada ce urmeaza eclozarii puilor. Ambele specii au un numar ridicat de pui si intrucat stufariile sunt ocupate de lacari, aceste specii isi cauta hrana prin pajistile si luncile de la marginea apelor. Emberiza schoeniclus prefera cuibaritul in stufarii intinse ca si Acrocephalus scirpaceus. In lipsa resurselor de hrana majoritatea populatiilor de presura de stuf- Emberiza schoeniclus se retrag in Delta Dunarii, unde, in anii favorabili, raman si peste iarna. Acrocephalus schoenobaenus si Remiz pendulinus sunt prezente in mod constant in zonele umede ale Moldovei.
Lacarul mic presupunem ca intra in concurenta pentru teritoriu cu lacarul de stuf cu care se aseamana in ceea ce priveste talia, comportamentul teritorial, modul de reproducere si de procurare a hranei.
Prezenta numerica scazuta a pitigoiului pungar (Remiz pendulinus) se datoreaza numarui scazut de zavoaie cu salcii de-a lungul elesteelor analizate, locuri preferate pentru cuibarit. Salciile au fost taiate, in cea mai mare parte, fie au fost acoperite de ape, in cazul inundatiilor de primavara,.
Locustella luscinioides prezinta o mare variatie a efectivului ea fiind o specie rar cuibaritoare in zonele umede ale Moldovei.
In ceea ce priveste deosebirile existente intre zonele umede in privinta abundentei specifice numarului de paseriforme de stuf se observa o mare variatie in zonele Vladesti si Balta Lata (vezi Fig. 4). Cele doua zone se afla la granita tarii. Influenta factorului antropic este relativ crescuta din cauza pescuitului si agriculturii intensive ce se practica in zonele umede inconjuratoare. Stufariile ocupa o suprafata mai mica de jumatate din luciul de apa, iar resursele trofice sunt impartite cu multe alte specii de pasari ce consuma insecte: prigorii, drepnele, grauri, ciori, ciocarlii, codobaturi, randunele, lastuni s.a.
O constanta mare in ceea ce priveste prezenta paseriformelor de stuf se intalneste in ecosistemul de la Larga Jijia- Vladeni, aici suprafetele de stuf sunt intinse si resursele trofice sunt foarte bogate si diverse in tot timpul anului.
Stationarele Beresti si Carja prezinta o variabilitate numerica a efectivelor speciilor scazuta si o situatie asemanatoare in ceea ce priveste abundenta passeriformelor de stuf (vezi Fig. 4). Daca numarul exemplarelor de paseriforme de stuf creste, atunci celalte specii de paseriforme, in cele doua zone, scad numeric din cauza limitarii spatiului de activitate. Ambele zone au o suprafata relativ mica de stuf ce acopera intinderile de apa. Alaturi de paserifomele insectivore de apa se afla specii de pasari acvatice cu care intra in concurenta pentru spatiul vital : rata mare, corcodel mare, lisite, gainusa de balta s.a, dar diversitatea este relativ scazuta.
In ce priveste zona Iezar din nordul Moldovei, daca urmarim axa 1 ce exprima 48% din variabilitatea valorilor efectivelor paseriformelor de stuf, deci o variatie negativa scazuta, adica numarul exemplarelor de paseriforme acvatice variaza foarte putin de la o specie la alta. Daca numarul de exemplare creste pentru o anumita specie (Acrocephalus scirpaceus), spre exemplu, numarul indivizilor de lacar mic (Acrocephalus schoenobaenus) scade. (a se vedea si Fig. 4). Zona Iezar, prin caracteristicile habitatului existent ofera conditii prielnice pentru coexistenta paserifomelor de stuf. Fluctuatiile numerice existente in ceea ce priveste aceste paseriforme de stuf se explica prin vremea capricioasa si o scadere a resurselor trofice. Un factor perturbator al vietii paseriformelor de stuf il reprezinta pescarii sportivi, mai ales braconierii care deranjeaza pasarile de la activitatile obisnuite.
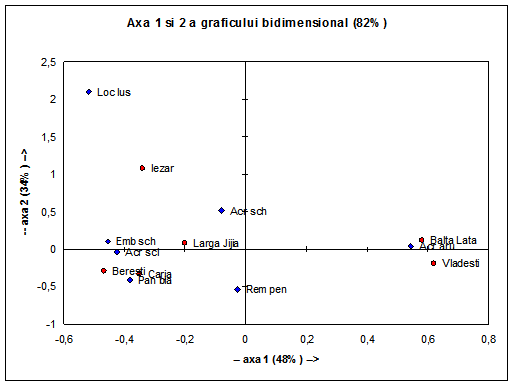
Fig. 4: Grafic bidimensional privind corespondeta intre speciile de paseriforme de stuf si zonele umede
Dominanta exprima abundenta relativa a unei specii, reprezentand raportul dintre efectivele unei specii si suma efectivelor celorlalte specii din zona studiata. Dominanta reprezinta un indicator al productivitatii, adica arata proportia in care participa fiecare specie la realizarea productiei de biomasa dintr-o biocenoza. In functie de valoarea procentului, speciile se impart in functie de dominanta (Gomoiu M, T., 2001) in: specii eudominate, cand procentul este > 10,1%; specii dominate, cand procentul este cuprins intre 5,1-10%; specii subdominante cand procentul este cuprins intre 2,1-5%; specii recedente cand procentul este cuprins intre 2,1-2% si specii subrecedente, cand procentul este sub 1,1%.
Din analiza indicelui de dominanta (vezi tabel 2) rezulta ca Acrocephalus arundinaceus este o specie eudominnata in toate zonele studiate, fiind principala specie de paseriform de apa ce se gaseste in stufariile din Moldova.
Acrocephalus scirpaceus este eudominanta in aproape toate zonele umede din Moldova, cu exceptia nord-estului Moldovei, la Balta Lata, unde este dominanta.
In general, specia Acrocephalus schoenobaenus este o specie dominanta in habitatele studiate, dar in zona Iezar, alaturi de celelalte doua specii de lacari, este o specie eudominanta, iar la Beresti efectivele inregistrate sunt mici. Analizand valoarea indicelui de dominanta putem confirma ca este o specie subdominanta.
Trebuie subliniat faptul ca toate speciile de silviide de apa gasite la Iezar- Dorohoi sunt eudominante, intrucat si valoarea dominantei pentru Locustella luscinioides este peste 10%. Zona Iezar este prielnica paseriformelor de apa datorita configuratiei aproape uniforma a habitatului. In zona Balta Lata, greluselul de stuf- Locustella luscinioides este o specie putin intalnita in sezonul de reproducere, fiind subrecedenta, iar la Larga Jijia este o specie recedenta.
Pitigoiul de stuf (Panurus biarmicus), care in cele mai multe cazuri inregistreaza efective relativ crescute, este o specie dominanta la Larga Jijia si la Balta Lata, iar in zonele Beresti si Carja este eudominanta.
Boicusul- Remiz pendulinus ca si in cazul speciei precedente prezinta efective crescute in zavoaiele umede, de aceea aceasta specie este eudominanta la Vladesti, Beresti si Carja, iar la Larga Jijia unde se inregistreaza un relativ echilibru intre paseriformele de stuf este dominanta.
Presura de stuf (Emberiza schoeniclus) , desi o prezenta discreta in stufariile zonelor umede inregistreaza in perioada de cuibarit efective crescute, de aceea, in toate zonele in care ea a fost observata este o specie eudominanta. (vezi tabel 2)
In general paseriformele de apa isi desfasoara ciclul de viata in stuf si in preajma lui, de aceea si valorile indicelui de dominanta indica caracteristici de specie eudominanta.
Tabel 2: Indicele de dominanta la paseriformele de stuf in zonele umede ale Moldovei
|
Specia |
|||||||
|
Zona |
Acr aru |
Acr sci |
Acr sch |
Loc lus |
Pan bia |
Rem pen |
Emb sch |
|
Larga Jijia | |||||||
|
Balta Lata | |||||||
|
Vladesti | |||||||
|
Beresti | |||||||
|
Iezar | |||||||
|
Carja | |||||||
In ecosistemele umede cercetate, din Moldova, speciile de paseriforme de stuf se concentreaza in jurul a doi clusteri sau puncte nodale. (Fig. 5) Unul este reprezentat de speciile Locustella luscinioides si Acrocephalus schoenobaenus care prezinta efective reduse in general si razlete. Al doilea mare grup de similaritate a indicelui de dominata il formeaza Acrocephalus arundinaceus si celelalte speciii de paseriforme de stuf.
Lacarul mare (Acrocephalus arundinaceus) domina in toate habitatele studiate fiind o specie raspandita pe suprafete intinse. Emberiza schoeniclus si Acrocephalus scirpaceus domina in teritoriile din interiorul suprafetelor cu stuf, de aceea si afinitatea ei este crescuta. Panurus biarmicus si Remiz pendulinus se concentreaza mai ales in habitatele de la marginea suprafetelor cu stuf, dar patrund uneori si in interiorul acestora, de aici si similaritatea lor ridicata cu lacarul de stuf (Acrocephalus scirpaceus) si presura de stuf (Emberiza schoeniclus). (vezi Fig. 5). Trebuie remarcat ca gruparea speciilor in functie de dominanta este in linii mari aceeasi cu cea in functie de valorile efectivului mediu (vezi Fig. 5) cu o singura deosebire. Acrocephalus schoenobaenus are valori ale efectivului asemanatoare cu ale speciilor Remiz pendulinus, Panurus biarmicus si de aceea prezinta o similaritate crescuta in ceea ce priveste numarul de indivizi. In schimb, in privinta dominantei Acrocephalus schoenobaenus nu este o specie prezenta in mod constant, in numar mare, in stufariile din zonele umede investigate de noi. De aceea Acrocephalus schoenobaenus prezinta a afinitate ridicata cu Locustella luscniodes (vezi Fig. 5) in ceea ce priveste dominanta, intrucat ambele specii prezinta efective relativ scazute si inconstante.
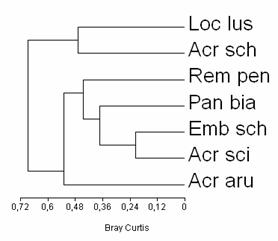
Fig. 5: Gruparea speciilor de paseriforme de stuf in functie de dominanta.
Constanta (Frecventa) este un indicator ce exprima continuitatea unei specii intr-un anumit teritoriu si reprezinta raportul procentual dintre numarul de probe in care apare o anumita specie si numarul total de probe analizate. In functie de valoarea constantei (Gomoiu M., T., 2001) in probe, speciile se pot imparti in urmatoarele categorii: specii accidentale, prezente in 1-25% din probe; specii accesorii, prezente in 25,1- 50% din probe; specii constante, prezente in 50,1- 75% din probe; specii euconstante, prezente in 75,1-100% din probe.
In ce priveste prezenta speciilor urmarite de noi in probe (vezi tabel 3), lacarul mare este o specie euconstanta, cu exceptia stationarelor Beresti si Iezar, unde, conform valorii indicelui de constanta se incadreaza in categoria speciilor constante. Lacarul mare este o pasare, ce poate fi vazuta in toate zonele umede cu stuf din Moldova. La Beresti, suprafetele compacte de stuf se amesteca cu papura, catina, tufarisuri, diverse plante ce creaza un mediu mai putin propice existentei acestei specii, prezenta in 57,14% din probe. (vezi tabel 3). Valoarea numerica, mai redusa a lacarului mare la Iezar din totalul observatiilor se datoreaza, in special, faptului ca spre sfarsitul perioadei de cuibarit indivizii clocitori se deplaseaza catre cartiere sudice. In nord, nord- estul Moldovei raman indivizi izolati. Uneori, cand conditiile sunt propice poposesc efective restranse numeric , care vin din nord, nord-estul Europei.
Ca si lacarul mare, lacarul de stuf- Acrocephalus scirpaceus este o specie euconstanta in majoritatea zonelor umede analizate. La Iezar, aceasta specie isi imparte teritoriul cu specii ca: Acrocephalus arundinaceus, Locustella luscinioides, Emberiza schoeniclus si se prezinta ca o specie constanta. Lacarul de stuf, la Vladesti, prezinta efective mai mici si o prezenta mai rara in stufarii, fiind conform valorii indicelui de constanta o specie constanta.
Lacarul mic, ca si celelalte specii de lacari, este o specie euconstanta in zonele Larga Jijia, Balta Lata si Iezar, unde aceasta specie poate fi observata in perioada de reproducere in mod constant. In zonele Vladesti si Carja, lacarul mic se incadreaza in categoria speciilor constante. Pe valea Siretului la Beresti, lacarul mic, apare sporadic, de aceea analizand valoarea indicelui de constanta (sub 25%) este o specie accidentala.
Locustella luscinioides inregistreaza o prezenta constanta doar in zona Iezar, unde de altfel am inregistrat si cea mai mare abundenta de paseriforme de stuf. In zona Larga Jijia prezenta sa o incadreaza ca o specie accesorie. In nord-estul Moldovei, la Balta Lata, apare foarte rar in perioada de cuibarit si de aceea se incadreaza in categoria speciilor accidentale.
In zonele umede in care temperaturile inregistrate de noi au fost mai ridicate, Panurus biarmicus- pitigoiul de stuf este o specie euconstanta. La Larga Jijia insa prezenata sa in perioada de reproducere am constatat-o doar in anii 2001 si 2003, nu si in anul 2002. Consideram ca prezenta sa in timpul sezonului de reproducere este mult legata si de valorile temperaturii si conditiile meteorologice. In anul 2002, in zona Larga Jijia- Vladeni, in perioadele iunie-iulie s-au inregistrat ploi repetate si temperaturi medii, de multe ori, sub 200C, ceea ce a determinat si absenta acestei specii.
Boicusul- Remiz pendulinus este o specie euconstanta in zonele umede in care se afla zavoaie de salcii (Carja, Beresti, Vladesti), dar in zona Larga Jijia, se incadreaza in categoria speciilor accesorii, pentru ca salciile ce constituie nisa preferata de cuibarit sunt rare.
Emberiza schoeniclus este o specie ce inregistreaza efective crescute in toate habitatele analizate. In zonele Larga Jijia si Iezar este o specie constanta, iar in celelalte zone este o specie euconstanta.
In ceea ce priveste constanta se observa ca speciile de paseriforme de stuf in zonele in care au fost gasite se incadreaza in clasele superioare de constanta. Locustella luscniodes face exceptie de la regula, in zona Balta Lata, unde se intalneste accidental in perioada reproducerii.
Tabel 3: Indicele de constanta la paseriformele de stuf in zonele umede ale Moldovei
|
Specia |
|||||||
|
Zona |
Acr aru |
Acr sci |
Acr sch |
Loc lus |
Pan bia |
Rem pen |
Emb sch |
|
Larga Jijia | |||||||
|
Balta Lata | |||||||
|
Vladesti | |||||||
|
Beresti | |||||||
|
Iezar | |||||||
|
Carja | |||||||
Lacarii, genul Acrocephalus, constituie prezente constante in habitatele studiate, ceea ce explica incadrarea lor intr-un grup cu similaritate crescuta. (vezi Fig. 6). Un alt grup cu similaritate crescuta in ceea ce priveste prezenta speciilor de paseriforme de stuf, in habitatele studiate, il formeaza pitigoiul de stuf (Panurus biarmicus) si presura de stuf (Emberiza schoeniclus), care in habitatele de pe valea Siretului si in bazinul mijlociu al Prutului constituie prezente obisnuite, dar nu intotdeauna regulate (vezi tabel 3). Ambele specii nu se regasesc in zona Vladesti.
Remiz pendulinus si Locustella lusciniodes prezinta o similaritate scazuta (vezi Fig. 5) cu celelalalte specii in ceea ce priveste constanta intrucat aceste doua specii constituie prezente destul de rare in habitatele studiate (vezi Fig. 5).
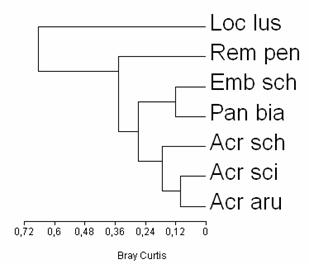
Fig. 5: Gruparea speciilor de paseriforme de stuf in functie de constanta
V.5 Indicele de semnificatie ecologica
Indicele de semnificatie ecologica Dzuba (W) reflecta pozitia paseriformelor de stuf in cadrul biocenozelor, deci relatia dintre indicele structural (constanta) si cel productiv (dominanta). Valorile acestui indice (Gomoiu M. T., 2001) determina o ierarhie a speciilor intr-o biocenoza. Dupa valorile acestui indice, speciile se subimpart in urmatoarele categorii: W1- specii subrecedente, cand acest indice are valori <0,1%; W2- specii recedente, cand acest indice este cuprins intre 0,1-1%; W3- specii subdominate, cand acest indice este cuprins intre 1,1-5%; W4- specii dominate, cand valorile indicelui sunt cuprinse intre 5,1- 10%; W5- specii eudominante, cand acest indice are valori > 10%.
Lacarul mare este o specie prezenta in mod frecvent, in numar mare, in habitatele analizate, de aceea se incadreaza in categoria W5, a speciilor eudominante caracteristice (tabel 4). In zona Beresti, deoarece prezenta sa este inconstanta iar efectivele mai scazute ca in celelalte zone, este o specie dominanta.
Lacarul de stuf este o specie caracteristica in toate zonele studiate, inregistrand atat efective crescute cat si o prezenta regulata in probele noastre.
Lacarul mic este o specie eudominanta in zona Iezar, unde aceasta specie constituie o prezenta obisnuita. In zona Larga Jijia este o specie dominata. In ambele zone anterior mentionate, care constituie excelente adaposturi si locuri de hranire pentru paseriformele de stuf lacarul mic- Acrocephalus schoenobaenus este o specie caracteristica. In biotopurile de la Balta Lata, Vladesti, Carja este o specie subdominata accesorie. In zona lacului Beresti, lacarul mic este o specie recedenta.
Ca si in cazul speciilor de lacari, in zona Iezar, Locustella luscinioides este o specie dominanta caracteristica. In ariile de studiu de la Balta Lata si Larga Jijia este o specie recedenta accesorie. Importanta acestei specii in biocenozele cercetate este redusa.
Pitigoiul de stuf se concentreaza numai in anumite zone si intr-un numar destul de ridicat. In zonele Beresti si Carja este o specie eudominata caracteristica. In zona lacului Balta Lata este o specie dominanta, iar la Larga Jijia este o specie subdominanta.
Remiz pendulinus prezinta un rol important mai ales in habitatele ce marginesc stufarisurile si luciul de apa, de aceea este o specie eudominanta caracteristica in zonele studiate. In zona Larga Jijia este specie accesorie.
Emberiza schoeniclus in toate biocenozele cercetate, incadrata in categoria W5- specie euconstanta caracteristica.
Tabel 4: Indicele de semnificatie ecologica Dzuba (W) la paseriformele de stuf in zonele umede ale Moldovei
|
Specia |
|||||||
|
Zona |
Acr aru |
Acr sci |
Acr sch |
Loc lus |
Pan bia |
Rem pen |
Emb sch |
|
Larga Jijia | |||||||
|
Balta Lata | |||||||
|
Vladesti | |||||||
|
Beresti | |||||||
|
Iezar | |||||||
|
Carja | |||||||
Din analiza graficului cu indicele de semnificatie ecologica (Fig. 6) reiese ca rolul cel mai important in biocenoza paseriformelor de stuf este indeplinit de lacarul mare- Acrocephalus arundinaceus. Un rol deosebit il prezinta in toate habitatele studiate lacarul de stuf- Acrocephalus scirpaceus si presura de stuf- Emberiza schoeniclus, care constituie pasari indicator pentru starea de sanatate a mediilor umede. Panurus biarmicus, Remiz pendulinus si Acrocephalus schonobaenus, a caror prezenta nu este intotdeauna regulata, efectivele inregistrate sunt destul de scazute.
Locustella luscinioides reprezinta un slab indicator al biocenozelor formate de paseriformele de stuf. Ponderea, in ceea ce priveste semnificatia ecologica a speciei Locustella lusciniodes este mica.
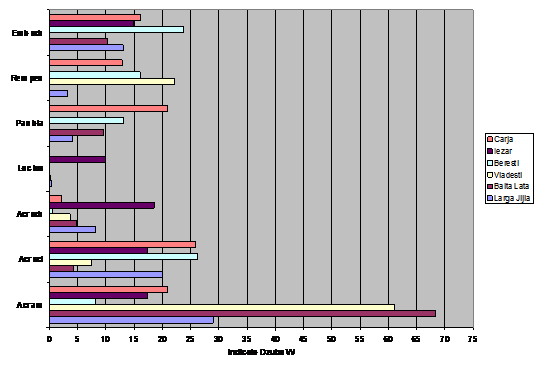
Fig. 6: Indicele de semnificatie ecologica Dzuba al paseriformelor de stuf in unele zone umede din Moldova
Indicele de afinitate cenotica reflecta legaturile existente intre speciile unei biocenoze. In functie de valorile acestui indice pot fi identificate speciile caracteristice, adica acestea au afinitatile cele mai mari.
Pentru evidentierea afinitatii cenotice am folosit indicele Bray- Curtis, deoarece acest indice tine seama atat de prezenta sau absenta unei specii dar si de numarul de indivizi al fiecarei specii. In cadrul matricei de calcul a dendrogramei Bray- Curtis privind afinitatea cenotica (Fig. 7) am inclus si celelalte paseriforme observate in zonele cu stufarii care stabilesc o serie de interrelatii cu paseriformele de apa. In elucidarea preferintelor de biotop am tinut seama de legaturile ce se stabilesc intre paseriformele de stuf si celelalte pasari observate in aceasta zona.
Indicele Bray- Curtis este un indice de disimilaritate (Gomoiu M. T., 2001) ce ia valori cuprinse intre 0 (corelatie maxima) si 1 (necorelatie).
Afinitatile cenotice existente intre speciile de paseriforme de stuf ne permit sa determinam care sunt speciile caracteristice stufariilor din zonele umede studiate.
Prin analiza dendrogramei, grafic 21 se observa ca cea mai mare afinitate este inregistrata intre speciile: Acrocephalus scirpaceus si Emberiza schoeniclus. Acestea sunt speciile caracteristice pentru ecosistemele acvatice. Afinitatea privind conditiile de mediu intre cele doua specii este foarte mare, indicele calculat avand valorile cele mai mici (indicele Bray-Curtis este un indice de disimilaritate ;100-BC ne da indicele de similaritate), care ne arata ca ambele specii coexista foarte bine in stufarii.
Din analiza dendrogramei (Fig. 7) reiese ca speciile genului Motacilla: M. alba si M. flava manifesta aceleasi preferinte de mediu cu speciile mai sus mentionate. Observatiile directeconfirma acest lucru. Aceste specii isi impart domeniul de hranire, situat la marginea stufariilor, unde codobaturile- Motacilla consuma prada (insecte) de dimensiuni mari, iar presura- Emberiza si lacarul de stuf- Acrocephalus scirpaceus consuma hrana de dimensiuni mai mici. Codobaturile se constiuie in specii indicatoare a biotopurilor situate la marginea stufariilor.
Remiz pendulinus si Panurus biarmicus prezinta o afinitate cenotica ridicata, intrucat ambele specii patrund mai rar in stuf, si viziteaza in multe cazuri tufarisurile si subarboretul de la marginea apelor. Coexistenta lor este asigurata si de locurile diferite de cuibarit, Remiz pendulinus cuibareste in salcii, iar Panurus biarmicus cuibareste in stufarii, la marginea acestora. Afinitatea cenotica ridicata este explicabila prin aceea ca cele doua specii prefera o varietate mai larga de habitate. Ambele specii au fost observate in terenuri deschise si in papurisuri.
Acrocephalus schoenobaenus formeaza cu toate speciile mai sus mentionate un alt grup in cadrul dendrogramei. Lacarul mic- Acrocephalus schoenobaenus este prezent in numar mai mic in stufariile din Moldova. Efectivele sale se concentreaza pe anumite habitate umede, si spre deosebire de ceilalti lacari poate fi observat realizand miscari active si rapide si in habitate eterogene. Adeseori am semnalat-o in papurisiri, palcuri de catina sau chiar terenuri deschise. Observatiile privind aceasta specie sunt mai putine decat pentru celelalte specii apartinand genului Acrocephalus, insa numarul exemplarelor identificate per observatii este mai mare in cele mai multe din iesirile in teren.
Nodul central al dendrogramei este format de lacarul mare- Acrocephalus arundinaceus si celelalte specii amintite anterior. Lacarul mare constituie in fapt specia de legatura intre paseriformele tipice zonelor umede si paseriformele mai putin caracteristice (Motacilla cinerea, Luscinia luscinia, Anthus campestris si Locustella luscinioides), un grup aparte de paseriforme apartine mai degraba mediului sinantrop dar viziteaza deseori habitatele acvatice (Passer domesticus, Passer montanus, Delichon urbica, Hirundo rustica si Riparia riparia).
Separarea paseriformelor de apa de celalte paserifome intalnite preponderent in pajistile de la marginea apelor nu este una stricta, intrucat multe din pasarile granivore ori insectivore intalnite in biotopuri cu vegetatie formata din ierburi si subarbusti pot fi vazute adeseori zburand deasupra intinderilor de stuf si papura in cautarea hranei- insecte. Capturarea acestora este dificila deoarece ele zboara la inaltimi mai mari ca paseriformele acvatice si numai rareori patrund in stufarii ori papurisuri.
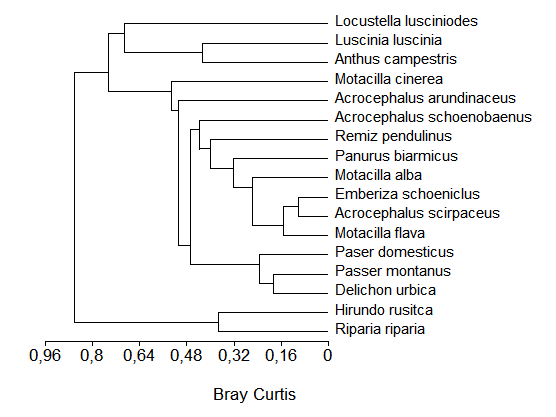
Fi. 7: Dendrograma afinitatii cenotice intre speciile de paseriforme in unele zone umede ale Moldovei
Identificarea speciilor genului Acrocephalus, urmarind caracteristicile morfologiei externe nu este elocventa. De aceea, in practica, se recurge la etaloane morfometrice. La fiecare specie limitele metrice sunt stabilite empiric. (Ion C, 2002).
In analiza biometrica am pornit de la ideea lui Darwin,1859, conform careia exista o stransa relatie intre morfologia unei specii si ecologia ei. In cazul lacarilor exista o stransa legatura intre lungimea aripii, a formei ei, a remigelor, si:distanta de zbor, miscarile ample pe care le realizeaza; intre greutate si disponibilitatile de hrana; intre greutate si sezonul fenologic. (Ion C, 2002).
Caracterele morfometrice calculate, indica adeseori diferente intre speciile de Acrocephalus de talie mica (A. scirpaceus, A. palustris, A. agricola, A. dumetorum, A. schoenobaneus, A. melanopogon si A. paludicola), precum si intre populatiile aceeleasi specii de Acrocephalus, situate in zone diferite sau capturate in sezoane distincte. (Ion C, 2002)
In cadrul studiului au fost capturate si masurate exemplare apartinand speciilor: Acrocephalus arundinaceus, A. scirpaceus, A. schoenobaenus). Datele obtinute au fost comparate cu unele rezultate existente in literatura de specialitate.
Caracterele metrice cercetate sunt: lungimea aripii (A), lungimea cozii (C), lungimea celei mai lungi remige primare (RI max; diferenta de lungime dintre cea mai lunga remige primara si lungimea: celei de-a doua remige primara (RI max- RII); celei de-a patra remige primara (RI max- RIV); celei de-a cincea remige primara (RI max- RV); celei de-a sasea remige primara (RI max- RVI), celei de-a saptea remige primara (RI max- RVII), a diferentei dintre lungimea celei mai lungi tectrice primara si lungimea primei remige primara (TP- R1), diferenta dintre lungimea primei remige primara si lungimea primei remige secundare (I-II). In afara caracterelor metrice s-a studiat si caracterul ponderal, greutatea (G).
Parametrii statistici calculati sunt: media, eroarea standard, mediana, deviatia standard, varianta, minimum (valoarea cea mai mica), maximum (valoarea cea mai mare), limita inferioara si limita superioara a intervalului de confidenta.
Masuratorile au fost efectuate cu o rigla ornitologica. Eroarea de calcul am estimat-o la aproximativ 1 mm.
Pentru aprecierea greutatii corpului s-a folosit un cantar Pesola 100gx1g.
Capturile s-au realizat trei ani consecutiv, 2001-2003, in zonele Larga Jijia, Beresti, Iezar, Balta Lata, Carja, Vladesti.
Modelul pe care l-am utilizat a mai fost folosit de catre ornitologii grupului Milvus care au studiat alcatuirea formulei alare la specia Parus ater. Noi am adaptat formula respectiva pentru genul Acrocephalus, ceea ce ne-a permis determinarea mai exacta a speciei. Formula alara se refera la stabilirea caracteristicilor metrice ale penelor de la aripi foarte utile in determinarea paseriformelor (Svensson L., 1992)
Pe baza capturilor am estimat, la nivelul populatiilor de lacari, limita superioara (LS) si inferioara (LI) a parametrilor calculati printr-o analiza descriptiva. Datele le-am prelucrat cu ajutorul programului Excell. (vezi tabel 5, 6, 7).
In ce priveste greutatea corpului la lacarul mare (Acrocephalus arundinaceus) ea se situeaza in jurul valorii de 30g (Iberian Birds Guide, 2000). Rezulatele noastre prelucrate statistic arata ca indivizii capturati in zonele umede ale Moldovei au o greutate mai ridicata (vezi tabel 5), lucru explicabil prin faptul ca resursele trofice sunt deosebit de bogate in aceste locuri, dovada fiind si diversitatea mare de pasari (Gache, 2002, Feneru, 2002, Rang, 2002, Trelea, 2002, Munteanu, 1968). La aceasta se adauga si impactul antropic destul de scazut si faptul ca indivizii de lacar mare din estul si sud-estul Europei migreaza pe distante mari catre sud-estul Africii si au nevoie de rezerve energetice mai mari.
La lacarii de stuf (Acrocephalus scirpaceus), intr-un studiu realizat pe mai multi ani in statiunea ornitologica Rybach, la Marea Baltica, (Bolshakov, 2000) greutatea corpului a fost la 12,84±0,54 g. Rezultatele noastre (vezi tabel 6) sunt aproape identice cu cele aratate.
Greutatea corpului la lacarii mici (Acrocephalus schoenobaenus) se situeaza in jurul valorii de 11-12 g (Cramp, 1992). Rezultatele obtinute de noi arata ca la indivizii capturati in plasele ornitologice valorile greutatii sunt putin mai mari, ca urmare a disponibilitatii mari de hrana- afide (Ion C, 2004), hrana preferata de paseriformele de stuf, dupa cum arata Michael, 2001.
Rezultatele obtinute de noi in privinta valorilor variabilelor privind aripa au fost comparate cu datele din literatura. (Hall, 2004; Hasselquist, 1998; Paspaleva, 1977; Svensson, 1992, Western Ukrainean Ornithological Union, Avosetta, 1997). Datele noastre, prelucrate statistic, sunt apropiate de cele furnizate de cercetatorii enumerati mai sus.
Urmarind valorile morfometrice ce au permis alcatuirea formulei alare la Acrocephalus arundinaceus se constata ca ele se incadreaza in limite mult mai mici decat cele gasite in literatura de specialitate (Hall, 2004; Svensson, 1992). Populatiile analizate de noi in teritoriile Moldovei, prin masuratorile facute ne conduc la ideea ca apartin ssp. arundinaceus. (Svensson, 1992)
La specia Acrocephalus scirpaceus valorile limitelor intervalului de confidenta sunt apropiate de cele gasite in literatura bibliografica (Paspaleva, 1977; Svensson, 1992; Hall , 2004; Hasselquist, 1998;). Lungimea aripii se apropie de limitele superioare indicate de lucrarile de specialitate. Populatiile de Acrocephalus scirpaceus existente in zonele umede din Moldova sunt situate la distante destul de mari de locul lor de iernare. Lungimea aripii la aceasta specie este in corelatie cu distanta de la locul lor de migratie si cu timpul de supravietuire (Hall, 2004).
Intre varsta pasarilor si lungimea cozii si a aripii s-a gasit un model de regresie non-lineara, unde lungimea aripii, atat la mascul cat si la femela, creste semnificativ in primul an de viata si in al doilea, pentru ca dupa al patrulea, al cincilea an sa scada. (Nowakowski, 2003). Aceste date ne conduc la parerea ca indivizii capturati de noi erau in primii trei ani de viata, si puteau fi apti pentru reproducere.
La lacarul mic (Acrocephalus schoenobaenus) am observat limite destul de largi ale unor caractere metrice (vezi tabel 6, RI max, I-II). Asa cum s-a prezentat in capitolul privind situatia capturilor, conditiile de mediu sunt foarte schimbatoare in zonele de studiu. De altfel Nowakowski, 2000, afirma ca factorii de mediu determina variatii mari in ceea ce priveste lungimea aripii si a corpului, iar disponibilitatile pentru hrana se modifica.
Uniunea Ornitologica Ucraineana, intr-un studiu din 1997, pe baza lungimii aripii, imparte in doua grupuri exemplarele apartinand speciei Acrocephalus schoenobaenus, pe baza lungimii aripii. (Western Ukrainean Ornithological Union, Avosetta, 1997). Dupa datele obtinute de noi, populatiile de lacari mic- Acrocephalus schoenobaenus se incadreaza in cel de al doilea grup in care lungimea aripii este cuprinsa intre 67-69 mm.
In general, limitele intervalului de confidenta pentru parametrii calculati, pe baza datelor culese in teren, se incadreaza in cele oferite de literatura de specialitate, cu mentiunea ca limita inferioara este, cel mai adesea, mai mica decat cea data de specialisti in determinarea silviidelor. Trebuie tinut seama ca in cazul masuratorilor efectuate de noi, penajul pasarilor in cartierele de cuibarit avea un grad de uzura ridicat din cauza densitatii vegetatiei din habitat si a tipului ei (preponderent stufaris). De asemenea masuratorile pe care le-am efectuat s-au referit la populatiile clocitoare si la indivizi aflati in pasaj, fie in sezonul prereproductiv, fie in cel postreproductiv. De aceea, asa cum vom observa din analiza statistica nu se diferentiaza pregnant, populatii distincte pentru cele trei specii de lacari la care am efectuat masuratori biometrice si ponderale, cu exceptia situatiei intalnite la specia Acrocephalus arundinaceus.
Tabel 5: Analiza discriminata la unele caractere ale speciei Acrocephalus arundinaceus
|
Acr aru |
G |
A |
C |
RI max |
RI max- RII |
RI max- RIV |
RI max- RV |
RI max-RVI |
RI max- RVII |
TP-R1 |
I- II |
|
Nr. obs. (exemplare) | |||||||||||
|
Media | |||||||||||
|
Eroarea standard | |||||||||||
|
Mediana | |||||||||||
|
Deviatia standard | |||||||||||
|
Varianta | |||||||||||
|
Minimum | |||||||||||
|
Maximum | |||||||||||
|
Interval de confidenta (95,0%) | |||||||||||
|
LS | |||||||||||
|
LI |
Tabel 6: Analiza discriminanta la unele caractere ale speciei Acrocephalus scirpaceus
|
Acr sci |
G |
A |
C |
RI max |
RI max- RII |
RI max- RIV |
RI max- RV |
RI max-RVI |
RI max- RVII |
TP-R1 |
I- II |
|
Nr. obs. (exemplare) | |||||||||||
|
Media | |||||||||||
|
Eroarea standard | |||||||||||
|
Mediana | |||||||||||
|
Deviatia standard | |||||||||||
|
Varianta | |||||||||||
|
Minimum | |||||||||||
|
Maximum | |||||||||||
|
Interval de confidenta (95,0%) | |||||||||||
|
LS | |||||||||||
|
LI |
Tabel 7: Analiza discriminata la unle caractere ale speciei Acrocephalus schoenobaenus
|
Acr sch |
G |
A |
C |
RI max |
RI max- RII |
RI max- RV |
RI max- RVI |
RI max-RVII |
RI max- RVIII |
TP-R1 |
I- II |
|
Nr. obs. (exemplare) | |||||||||||
|
Media | |||||||||||
|
Eroarea standard | |||||||||||
|
Mediana | |||||||||||
|
Deviatia standard | |||||||||||
|
Varianta | |||||||||||
|
Minimum | |||||||||||
|
Maximum | |||||||||||
|
Interval de confidenta (95,0%) | |||||||||||
|
LS | |||||||||||
|
LI |
Datele din tabele prelucrate statistic, cu un prag de incredere de 95%, au fost supuse analizei multivariate. Noi am incercat sa gasim care din caracterele cercetate explica diferente ale analizei metrice. Prin analiza principalelor componente (PCA) a fost posibil acest lucru si s-a putut realiza un sumar al structurii de date cantitative si mentinerea doar a factorilor principali (Fowler J., 1998) si simplificarea numarului de variabile in probe.
Analiza pricipalelor componente (PCA) urmareste sa explice care din combinatiile de variabile luate in calcul explica in cea mai mare masura variatia valorilor existente in interiorul setului de date multivariate. Axa F1 explica variabilitatea maxima, iar axa F2 explica variabilitatea minima.
La populatiile de Acrocephalus arundinaceus, caracterele discriminante care explica variatia biometrica si ponderala dintre indivizii capturati sunt: -diferenta dintre lungimile celei mai lungi remige primare si remigea a IV- a (RI max- RIV)
- diferentele dintre lungimile celei mai lungi remige primare si remigea a V-a (RI max- RV)
- diferenta dintre lungimile celei mai lungi remige primare si remigea a VI- a (RI max- RVI)
- diferenta dintre lungimile celei mai lungi remige primare si remigea a VII- a (RI max- RVII
- lungimea cozii (C)
- lungimea aripii (A)
- lungimea celei mai lungi remige primare (RI max)
Valorile acestor variabile prezentate sunt puternic corelate intre ele.
Cercul din Fig. 8 reda corelatia intre variabilele urmarite. Cu cat variabilele sunt pozitionate pe graficul PCA mai aproape de marginile cercului, atunci coeficientul de corelatie are valori apropiate de 1 sau -1, deci corelatia este mare. Daca variabilele se inscriu mai aproape de centrul cercului, coeficientul de corelatie are valori apropiate de zero.
Valorile inregistrate la greutate (G) ne arata (Fig. 9) ca acest caracter explica intr-o mica masura variatiile dintre indivizii populatiei.
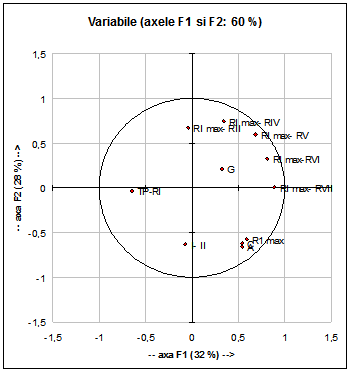
Fig. 8: Analiza principalelor componente (F1 si F2) privind variabilele analizate la specia Acrocephalus arundinaceus.
Valorile caracterelor analizate mai sus si prezentate in Fig. 9 explica doar 60% din variabilitatea urmarita. Din aceasta cauza am analizat separat numai variabilele ce au cea mai mare discriminanta si anume:- diferentele dintre lungimea celei mai lungi remige primare si lungimea remigii a IV- a (RI max- RIV), vezi Fig. 9.
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigii a V- a (RI max- RV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigii a VI- a (RI max- RVI)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigii a VII- a (RI max- RVII)
lungimea celei mai lungi remige primare (R1 max)
lungimea cozii (C)
lungimea aripii (A)
Ele explica 79% din variabilitate.
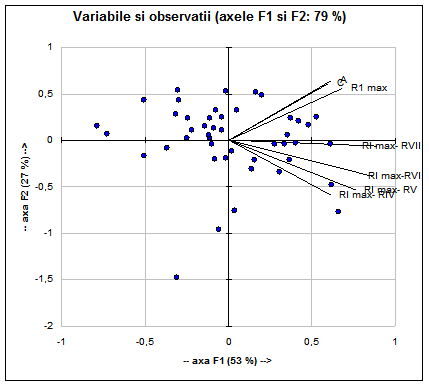
Fig. 9: Analiza principalelor componente (F1 si F2) privind variabilele si observatiile (ilustrate prin puncte negre) cu discriminanta cea mai mare la specia Acrocephalus arundinaceus
La populatiile de Acrocephalus scirpaceus (Fig. 10) caracterele discriminate care explica variatia biometrica si ponderala dintre indivizii capturati sunt: - lungimea cozii (C)
lungimea aripii (A)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a IV-a (RI max- RIV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RVI)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV)
Aceste caractere sunt corelate puternic intre ele (vezi Fig. 10) si explica doar 52% din variabilitatea valorilor obtinute de noi.
Variabilele greutate (G), diferenta dintre lungimea celei mai lungi remige primare si lungimea celei de-a II-a remige primare (RI- RII), diferenta dintre lungimea celei mai lungi tectrice primare si lungimea primei remige primare (TP- RI), diferenta intre lungimea celei mai lungi remige primare si lungimea primei remigi secundare (I-II), lungimea celei mai lungi remige primare (RI max) explica intr-o mica masura varibilitatea metrica si ponderala existenta in cadrul populatiei.
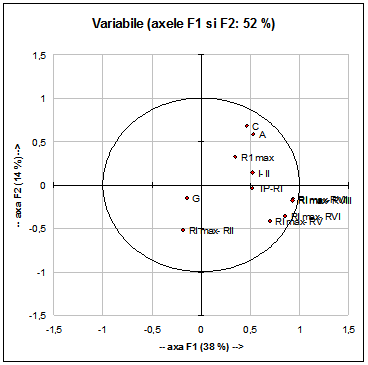
Fig. 10: Analiza principalelor componente (F1 si F2) privind variabilele analizate la specia Acrocephalus scirpaceus
Caracterele care au discriminanta cea mai mare sunt: lungimea cozii (C), vezi Fig. 11.
lungimea aripii (A)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a IV-a (RI max- RIV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RVI)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV).
Ele, singure, explica 77% din variabiliate
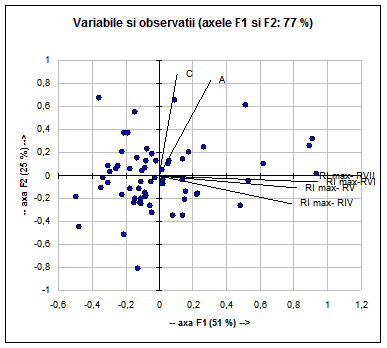
Fig. 11: Analiza principalelor componente (F1 si F2) privind variabilele si observatiile (ilustrrate prin puncte negre) cu discriminanta cea mai mare la specia Acrocephalus scirpaceus
Caracterele care detin informatia esentiala privind masuratorile efectuate pe exemplare capturate de Acrocephalus schoenobaenus in analiza discriminanta (vezi Fig. 12) sunt: -diferenta intre lungimea celei mai lungi remige primare si remigea a VII-a (RI max- RVII)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RVI)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a IV-a (RI max- RIV)
Aceste variabile explica insa o mica proportie variabilitatii caracterelor urmarite (50%).
Variabile ca: diferenta intre lungimea celei mai lungi remige primare si lungimea remigei a 2-a (RI Max- RII), diferenta intre cea mai lunga tectrice primare si lungimea primei remige primare (TP-RI), lungimea cozii (C), diferenta intre lungimea celei mai lungi remige primare si lungimea primei remige secundare (I-II) nu sunt corelate decat in mica masura si nu sunt importante in explicarea variabilitatii constatate la indivizii masurati.
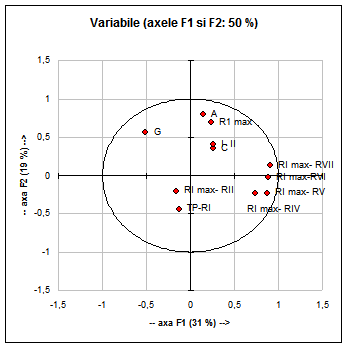
Fig. 12: Analiza principalelor componente (F1 si F2) privind variabilele analizate la specia Acrocephalus schoenobaenus
Deoarece caracterele descrise mai sus in cazul speciei Acrocephalus schoenobaenus nu explica decat 50% din variabilitate, am analizat separat caracterele cele mai discriminante: -diferenta intre lungimea celei mai lungi remige primare si remigea a VII-a (RI max- RVII), vezi Fig. 13.
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RVI)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV)
diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a IV-a (RI max- RIV)
Aceste variabile explica 88% din variabilitatea valorilor obtinute si sunt puternic corelate intre ele. O corelatie puternica exista intre diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RIV) si diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a V-a (RI max- RV), pe de-o parte si intre diferenta intre lungimea celei mai lungi remige primare si remigea a VII-a (RI max- RVII) si diferenta dintre lungimea celei mai lungi remige primare si lungimea remigei a VI-a (RI max- RVI)
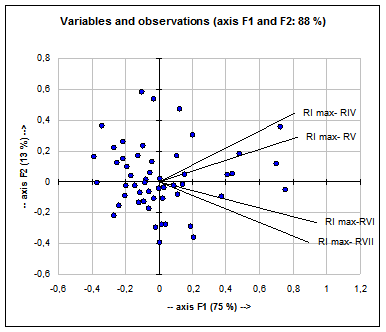
Fig. 13: Analiza principalelor componente (F1 si F2) privind variabilele si observatiile (ilustrrate prin puncte negre) cu discriminanta cea mai mare la specia Acrocephalus schoenobaenus
Observatiile efectuate de noi (vezi grafic 9, 11, 13) cuprind o plaja larga de valori. Ele nu se aranjeaza in grupuri izolate. Nu exista hiatusuri care sa indice anumite diferentierii nete intre variabilele masurate la exemplarele de lacari apartinand aceleasi specii. Datele adunate le consideram edificatoare si cuprind cea mai mare parte a variabilitatii caracterelor metrice si ponderale in interiorul populatiilor de lacari.
In cazul in care diferentele calculate se dovedesc a fi semnificative se recurge la comparea probelor, doua cate doua, cu ajutorul testului non-parametric Mann- Whitney (Varvara M, 2001), pentru un prag de semnificatie α
Variabilele care le-am luat in calcul sunt cele care permit alcatuirea unei formule alare complete si precise: RI max- RIV, RI max- RV, RI max- RVI, RI max- RVII.
Test Kruskal-Wallis pentru Acrocephalus arundinaceus
referitor la variabila RI max-RIV: H=5,117;
- referitor la variabila RI max-RV: H=16,662;
- referitor la variabila RI max-RVI: H=15,650;
- referitor la variabila RI max-RVII: H=18,346;
Rezultatele celor patru teste conduc catre aceeasi concluzie ca nu putem accepta ipoteza nula in care se afirma ca nu exista nici o diferenta intre cele patru populatii.
Prin compararea probelor doua cate doua, cu ajutorul testului Mann-Whitney, s-a dovedit ca intre populatiile de lacari de la Balta Lata si Beresti nu exista diferente semnificative in ceea ce priveste caracterul RI max-RIV: U observat=76,500, Ucritc=77,000, p=0,489,
In privinta diferentei RI max- RV pentru probele efectuate la Larga Jijia si Balta Lata, s-a constat ca U observat=52, 500, Ucritc=49,000, p=0,489, =0,05. Cu alte cuvinte diferentele intre populatiile analizate nu sunt semnificative. Pentru aceleasi zone s-a observat ca in ce priveste caracterul RI max- RVI nu exista nici o diferenta semnificativa intre grupurile de indivizi apartinand celor doua zone U observat=31,000, Ucritc=49,000), ca si pentru caracterul RI max- RVII (U observat=25, 000, Ucritc=49,000).
In celelalte cazuri diferentele statistice pe baza testului Mann- Whitney, intre populatiile investigate, pe baza caracterelor metrice sunt evidente si resping ipoteza nula conform careia nu exista nici o diferenta intre populatii, ceea ce ne determina sa afirmam ca in zonele cercetate se diferentiaza populatii locale, insa fara o diferentiere neta intre ele.
Combinand rezultatele testului Kruskal- Wallis cu testul Mann- Whitney, a reiesit ca se diferentiaza populatii locale in privinta caracterelor metrice pentru specia Acrocephalus arundinaceus.
Pentru zonele Larga Jijia si Balta Lata, se observa ca cele doua populatii de Acrocephalus arundinaceus sunt aproximativ asemanatoare in ce priveste formula alara. Lungimea remigelor primare (principalele pene de zbor) este in stransa corelatie cu caracteristicile habitatului si cu distanta de zbor (Ion C; 2003, a). S-a constat ca cele doua zone umede sunt asemanatoare in privinta habitatelor in sensul ca marginea baltilor si chiar si interiorul lor sunt dominate de stufarii compacte cu petece rare de papura.
Testele efectuate pentru specia Acrocephalus scirpaceus nu au luat in considerare si zona Balta Lata unde capturile au fost foarte putine.
Test Kruskal-Wallis pentru Acrocephalus scirpaceus:
- in ceea ce priveste variabila RI max-RIV: H=1,543;
- in ceea ce priveste variabila RI max-RV: H=0,980;
- in ceea ce priveste variabila RI max-RVI: H=0,534;
- in ceea ce priveste variabila RI max-RVII: H=0,455;
Conform rezulatatelor de mai sus, insemana ca decizia care trebuie luata cu o probabilitate de reusita de 95% este de a nu respinge ipoteza nula conform careia nu exista nici o diferenta intre populatiile de la Larga Jijia, Beresti si Iezar. Cu alte cuvinte diferenta intre cele trei populatii nu este semnificativa.
Aplicand testul Mann- Whitney s-a observat ca: pentru zonele Beresti si Iezar, in cazul parametrului RI max- RIV arata ca nu exista nici o diferenta semnificativa intre cele doua grupari de indivizi a celor doua zone. (Uobservat=57,000, Ucritic=72,000). Tot in zone Beresti si Iezar pentru parametrul RI max- RV calcule statistice (Uobservat=68,000, Ucritic=72,000) arata ca nu exista nici o diferenta intre populatiile celor doua zone.
Diferente semnificative nu s-au constat nici intre populatiile zonelor Larga Jijia si Iezar in ceea ce priveste caracterul RI max- RVI (Uobservat=318,000, Ucritic=333,000). Pentru acelasi caracter nu s-au observat diferente intre zonele Beresti si Iezar (Uobservat=58,000, Ucritic=72,000), la populatiile de lacar de stuf analizate metric.
Asociind rezulatele celor doua teste statistice utilizate se observa ca lipsa unei diferente nete intre populatiile celor trei zone investigate este semnalata doar in cazul factorului RI max- RVI. Populatiile lacarului de stuf, in cele trei zone umede prezinta foarte mici deosebiri.
Test Kruskal-Wallis pentru Acrocephalus schoenobaneus
- referitor la variabila RI max-RIV: H=7,811; =7,777, ceea ce inseamna ca nu putem accepta ipoteza nula conform careia nu exista nici o diferenta intre cele patru populatii, dar in alta ordine de idei, diferenta nu este semnificativa. (valoarea p caclulata este 0,05, α=0,05, ceea ce insemna conform ipotezei alternative ca poate interveni o eroare de tip α, adica diferenta dintre grupurile investigate nu este semnificativa
- referitor la variabila RI max-RV: H=9,579; =7,777, ceea ce inseamna ca nu putem accepta ipoteza nula conform careia nu exista nici o diferenta intre cele patru populatii.
- referitor la variabila RI max-RVI: H=10,820; =7,777, ceea ce inseamna ca nu putem accepta ipoteza nula conform careia nu exista nici o diferenta intre cele patru populatii.
- referitor la variabila RI max-RVII: H=11,707; =7,777, ceea ce inseamna ca nu putem accepta ipoteza nula conform careia nu exista nici o diferenta intre cele patru populatii.
Comparand populatiile cercetate doua cate doua prin aplicarea testului nonparametric non-parametric Mann- Whitney s-a observat ca:
-in privinta caracterului RI max- RIV in zonele Balta Lata si Beresti nu exista o diferenta semnificativa statistic. (U observat=11,500, Ucritc=11,000, p=0,457, =0,05); intre populatiile din zonele Balta Lata si Iezar (U observat=64,000, Ucritc=71,500), precum si cele de la Beresti si Iezar (U observat=10,500, Ucritc=13,000), diferentele constatate nu sunt statistic semnificative.
- in privinta caracterului RI max- RV nu s-au constatat diferente statistice intre populatiile de la Balta Lata si Iezar (U observat=70,500, Ucritc=71,000, p=0,476, =0,05) si cele de la Beresti si Iezar (U observat=5,500, Ucritc=13,000)
- in privinta caracterului RI max- RVII in zonele Beresti si Iezar (U observat=6,000, Ucritc=13,000) reiese ca nu exista nici o diferenta populatiile apartinand celor doua zone.
In cazul speciei Acrocephalus schoenobaenus comparand testele folosite in cazul factorului RI max- RIV se poate conchide ca populatia de la Larga Jijia este diferita de cea existenta in zonele Balta Lata, Beresti si Iezar.
Populatiile de la Beresti si Iezar apartin aceleasi populatii in privinta caracterelor metrice analizate, cu exceptia factorului RI max- RVI.
In cazul celorlalte variabile, populatiile nu sunt diferite, statistic semnificativ, cu o probabilitate de 95%, in cazul caracterelor metrice analizate.
Lungimea caracterelor metrice urmarite variaza la fiecare specie in functie de distanta pe care trebuie sa o parcurga in migratie si de talia corpului in unele cazuri, asa cum s-a explicat anterior.
In general, in zonele investigate se pot observa unele diferentieri ale populatiilor pe baza anumitor caractere metrice, dar fara a putea afirma ca in cele patru zone se afla patru populatii locale distincte morfologic, cu exceptia speciei Acrocephalus arundinaceus- lacarul mare, in cele patru zone se diferentiaza populatii locale in privinta caracterelor metrice. La lacarul de stuf se observa cele mai mici diferente intre populatiile din cele patru zone izolate spatial.
Cantecul la pasarile din familia Sylviidae, ordinul Passeriformes, din care face parte si genul Acrocephalus reprezinta un criteriu de identificare a speciei in sezonul de reproducere. Studiul cantecului la o anumita speciilor poate stabili rolul indivizilor in relatiile cu congenerii sai sau cu indivizii altor specii.
Structura cantecului la speciile genului Acrocephalus este destul de asemanatoare, iar de multe ori intre indivizii aceleeasi specii se inregistreaza unele diferente. Cantecele in sezonul de reproducere sunt fie teritoriale, fie de avertizare a partenerilor sexuali (Cramp S., 1992).
Inregistrarile sonore au fost realizate de noi cu ajutorul unui reportofon Sony si a unui microfon omnidirectional. Parametrii urmariti au fost intensitatesa (amplitudinea relativa), frecventa (inaltimea sunetului), si numarul de fonotoame (timbrul). Emisiile sonore au fost prelucrate apoi cu programe de specialitate (Spectraplus) si au fost realizate grafice ale amplitudinii relative medii, in functie de durata emisiilor sonore, spectograme, spectre ale amplitudinii relative medii si in final s-a realizat o analiza descriptiva pentru evidentierea caracteristicilor structurii cantecului speciei.
In cadrul unui singur cantec am evidentiat mai multe unitati distincte (Ion C, 2003, b) pe care le-am notat cu literele alfabetului latin. Unitatile au aceeasi forma, aceeasi freventa si durata in zona amplitudinii maxime. Intre unitatile ce compun o emisie sonora intervin pauze foarte scurte. O unitate poate fi considerata, o entitate ce se repeta de cel putin doua ori in repertoriul vocal al masculului. (Hasselquist D., 1998). Cu alte cuvinte cantecele analizate reprezinta semnale cu o structura complexa formate din note sau silabe, adica din unitati sonore distincte si indivizibile avand o durata medie de 0,1 secunde; notele sunt asociate in motive sau fraze. (Cociu M., 1999)
Pentru a reliefa diferentele mici existente in cantecul celor trei specii de lacari mai sus amintite am realizat graficul suprapus al spectrului mediu (relatia intre frecventa si amplitudinea relativa medie) Cantecele supuse acestui tip de analiza au fost prelevate in aceeasi perioada, aceeasi zi si in aceeasi ora. Asemanarea dintre cantecele diverselor specii de lacari este destul de mare. Se observa ca amplitudinile maxime ale cantecului sunt atinse, la cele trei specii, in jurul acelorasi valori ale frecventei 3-4 KHz.
Cantecele speciilor Acrocephalus arundinaceus si Acrocephalus scirpaceus sunt asemanatoare intre ele in comparatie cu cel al speciei Acrocephalus schoenobaenus. Spectrele medii ale cantecului la cele doua specii de lacari: lacarul mare- Acrocephalus arundinaceus si lacarul de stuf- Acr scirpaceus sunt, aproape identice. La frecvente mici amplitudinea creste, pentru ca dupa frecvena de 4 KHz aceasta sa scada brusc. La specia Acrocephalus schoenobanus amplitudinea scade mult catre valori mari ale frecventelor. Cu alte cuvinte, sunetele emise de lacarul mare si mic sunt mai rapide, mai subtiri. La lacarul mic- Acrocephalus schoenobaenus sunetele sunt mai ample, cu tonalitate mai groasa, joasa.
VII.1 Analiza cantecului la specia Acrocephalus arundinaceus (lacarul mare)
Pentru Acrocephalus arundinaceus am determinat sase unitati distincte ale cantecului. (Fig. 14).
D
Fig. 14: Spectograma cantecului de la lacarul mare
Unitatea de tip A reprezinta un sunet putenic, lung de aproximativ de 200 ms. Este cel mai lung tip de sunet inregistrat de noi. Varful amplitudinal corespunde unei frevente de 2 KHz. (Fig. 15, 16)
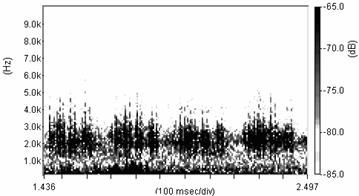
Fig. 15: Spectograma unitatii de tip A la lacarul mare
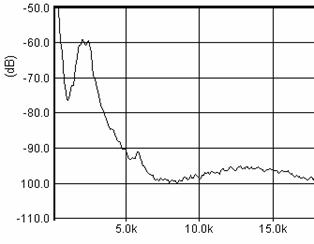
Fig. 16: Spectrul amplitudinii relative a unitatii de tip A la lacarul mare
Unitatea de tip B are o durata mai mica de 40- 50 ms, fiind un sunet foarte scurt. Energia sunetelor nu se concentreaza intr-un singur maxim, ci in mai multe asemanatoare cu amplitudini ce corespund unor frevente de 1,8 KHz, 3 KHz, 4,7 KHz, 6,3 KHz. (Fig, 17, 18)

Fig. 17: Spectograma unitatii de tip B la lacarul mare
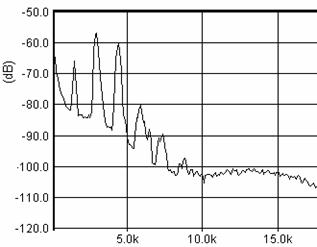
Fig. 18 Spectrul amplitudinii relative a unitatii de tip B la lacarul mare
Tipul B se aseamana foarte mult dar durata este mai mare, are mai multe varfuri amplitudinale ca si sunetul de tip B, dar mult mai bine conturate. Amplitudinile maxime corespund unor frecvente mai mari. (Fig. 19, 20)

Fig. 19: Spectogrma unitatii de tip B' la lacarul mare
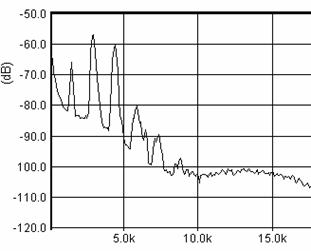
Fig. 20: Spectrul amplitudinii relative a unitatii de tip B la lacarul mare
Tipul C este o emisie sonora scurta , foarte subtiere. Durata este mica de 100 ms, iar varful amplitudial corespunde unei frecvente de 3KHz. (Fig. 21, 22)
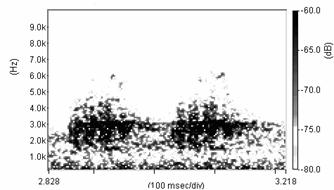
Fig. 21: Spectograma unitatii de tip C la lacarul mare
Fig. 22: Spectrul amplitudinii relative a unitatii de tip C la lacarul mare
Sunetul de tip D se asemana intrucatva cu cel de tip B, insa amplitudinea maxima este distribuita pe mai multe frecvente. Varful cel mai inalt al amplitudinii este de 4 KHz. Durata acestui tip de sunet este de 100 ms. (Fig. 23, 24)

Fig. 23: Spectograma unitatii de tip D la lacarul mare
Fig. 24: Spectrul amplitudinii relative a unitatii de tip D la lacarul mare
Unitatea de tip D nu este cu mult diferita de unitatea sonora de tip D, dar are mai multe varfuri amplitudinale. Varful amplitudinal maxim corespunde unei frecvente de 3 KHz. Sunetul de tip D' este putin mai slab, mai subtire ca cel de tip D. (Fig. 25, 26)

Fig. 25: Spectograma unitatii de tip D la lacarul mare
Fig. 26: Spectrul amplitudinii relative a unitatii de tip D la lacarul mare
Durata medie a cantecului inregistrat in perioada de reproducere este de 3,91± 0,672 s. Durata cea mai lunga a cantecului a fost de 8,59 s, in timp ce cel mai scurt a fost de 1,68 s. Numarul de unitati per cantec la Acrocephalus arundinaceus este 16,74± 3,13. Numarul maxim de unitati decelate intr-un singur cantec este de 40, ia cel mai mic de 8. (tabel 8)
Numarul de unitati distincte identificate intr-un cantec este de 3,696± 0,3596 pentru un prag de precizie de 95%. Numarul maxim de unitati diferite emise intr-un repertoriu sonor este de 5, iar minumum de 2. Parametrii estimati de noi sunt in concordanta cu cei consemnati in literatura de specialitate (Cramp S., 1992). Numarul de silabe calculat de noi, este putin mai mare ca cel calculat de alti cercetatori (Cramp S., 1992). Consideram ca aceasta diferenta apare datorita faptului ca inregistrarile au fost facute, in perioada considerata cea mai activa, de imperechere.
Tabel 8: Analiza descriptiva a cantecelor analizate la lacarul mare
|
Statistici |
Durata (s) |
Nr. unitati/cantec |
Nr. tipurilor/cantec |
|
Nr obs. (cantece) | |||
|
Medie | |||
|
Eroare stabdard | |||
|
Mediana | |||
|
Deviatie standard | |||
|
Amplitudine | |||
|
Minimum | |||
|
Maximum | |||
|
Er std. x t(0,05,22) |
Durata cantecului este corelata cu numarul de unitati dintr-un cantec, in proportie ridicata. (r2 ). Valoarea crescuta a coeficientului de determinare demonstreaza ca aproximativ 92% din valorile celor doua variabile calculate sunt corelate. (Fig. 27)
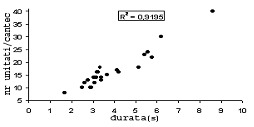
Fig. 27: Corelatia intre numarul de unitati dintr-un cantec si durata sa la lacarul mare
Pentru a contura o imagine de ansamblu asupra cantecului speciei Acrocephalus arundinaceus, in sezonul de reproducere, am realizat un spectru mediu pentru cantecele urmarite. Pentru fiecare cantec s-a extras o medie de 252 de frecvente cu amplitudinile relative corespunzatoare. Fiecarei frecvente i s-a calculat media valorilor amplitudinale pentru un interval de confidenta de 95 %. (Fig. 28)
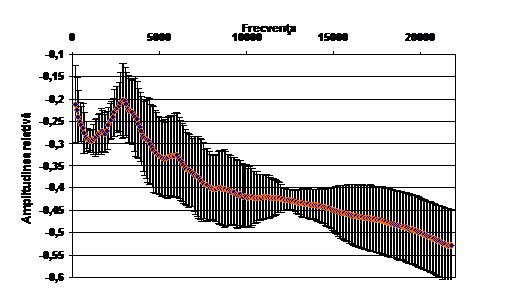
Fig. 28: Spectrul mediu al cantecelor la lacarul mare
Varful amplitudinal este atins la frecvente de 3 KHz. La frecvente mai mari se observa ca amplitudinile relative medii scad. O usoara crestere a amplitudinilor relative medii se observa intre frecventele de 2-3 KHz.
Analiza cantecului la Acrocephalus scirpaceus in timpul perioadei de reproducere a evidentiat cinci tipuri de silabe sau unitati: A, B, C, D, E.. Unitatile de tip B si D sunt foarte asemanatoare, numai ca valorile amplitudinii maxime se mentin ridicate in cazul silabei de tip D, pe mai multe varfuri ale frecventei comparativ cu tipul B. Tipul E se asemana si el intrucatva cu cu cel de tip B si D, ca forma, durata, dar inregistreaza un hiatus al amplitudinii in zona frecventei de 2.5 KHz. (Fig. 29)
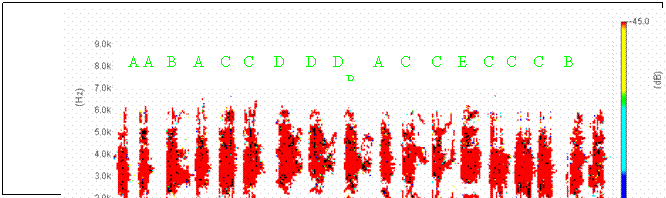
Fig. 29: Spectograma unui cantec la lacarul de stuf
Unitatea de tip A are o durata de aproximativ 100 ms, (Fig. 30), al carei frecvente se incadreaza intre 1- 4 KHz. Sunetul este unul destul de gros, iar amplitudinea maxima este atinsa la frecventa de 2,5 KHz. Sunetul se prezinta cu mai multe varfuri ale inaltimii, iar amplitudinea creste si descreste usor inspre si dinspre aceste valori maxime ale freventei. (Fig. 31)
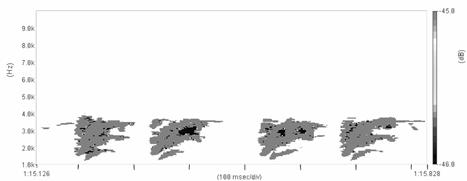
Fig. 30: Spectograma unitatii de tip A la lacarul de stuf
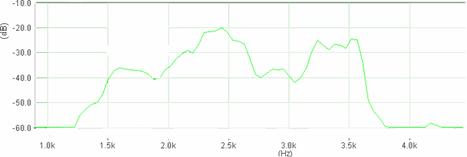
Fig. 31: Spectrul amplitudinii relative a unitatii de tip A la lacarul de stuf.
Sunetul de tip B are o valoare mai mica de 100 ms, (Fig. 32), iar valorile amplitudinale maxime sunt distribuite pana la valori mari ale frecventei 6 KHz. Unitatea de tip B inregistreaza trei maximuri amplitudinale mari in dreptul frecventelor de 1,7 KHz, 2,5 KHz , 3,1 KHz si un alt varf mai mic in dreptul valorii de 3,9 KHz. (Fig. 33)
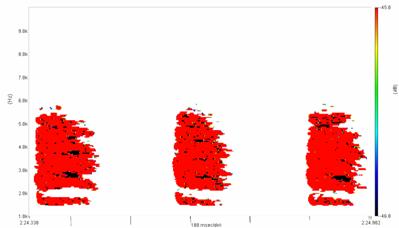
Fig. 32. Spectograma unitatii de tip B la lacarul de stuf
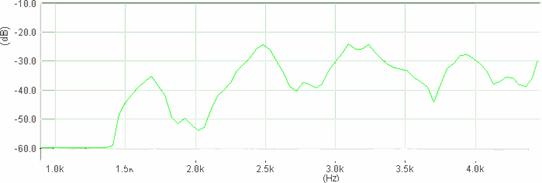
Fig. 33: Spectrul amplitudinii relative a unitatii de tip B la lacarul de stuf
Unitatea de tip C are o durata ceva mai mare de 100 ms si nu inregistreaza amplitudini crescute la frecvente scazute. Energia sunetului se distribuie pe trei varfuri amplitudinale din care cel mai mare este la 2,7 HKz. (Fig. 34, 35)
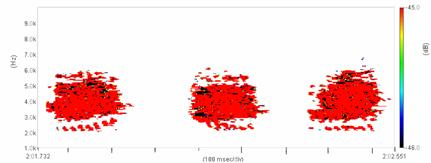
Fig. 34: Spectograma unitatii de tip C la lacarul de stuf
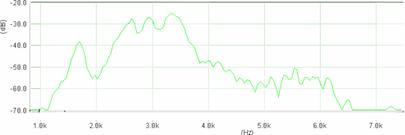
Fig. 35: Spectrul amplitudinii relative a unitatii de tip C la lacarul de stuf
Sunetul de tip D are o durata de aproximativ 200 ms, fiind astfel cel mai lung tip de unitate in cantecul speciei Acrocephalus scirpaceus. Amplitudinea maxima a cantecului se distribuie intre frecventele 2,8-4 KHz cu mai multe varfuri apropiate ca valoare. (Fig. 36, 37)

Fig. 36: Spectograma unitatii de tip D la lacarul de stuf
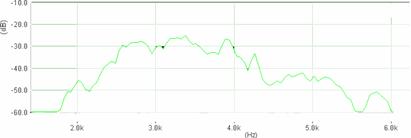
Fig. 37: Spectrul amplitudinii relative a unitatii de tip D la lacarul de stuf
Unitatea de tip E are durata asemanatoare cu sunetul de tip A si B, de 100 ms. Sunetul de tip E inregistreaza cea mai ridicata amplitudine ca si sunetul de tip A (-20DB), dar in dreptul frecvenei de 4 KHz (si nu de 2,5 KHz). Este un sunet mai subtire si mai putin bine conturat din punct de vedere al intensitatii la frecvente mici. (Fig. 38, 39)

Fig. 38: Spectograma unitatii de tip E la lacarul de stuf
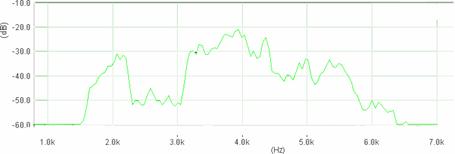
Fig. 39: Spectrul amplitudinii relative a unitatii de tip E la lacarul de stuf
Durata medie a cantecului la Acrocephalus scirpaceus este destul de ridicata: 4,135556±2,002135. Cel mai lung cantec inregistrat, de noi, a fost de 9,362 secunde. Cea mai scurta emisie sonora a fost de 0,4 secunde, inregistrata spre sfarsitul perioadei de reproducere. Durata cantecului la aceasta specie este putin mai mare ca la lacarul mare. Cantecul este asociat, in cele mai multe cazuri, miscarilor ample si mai agitate pe care le executa in perioada de reproducere.
Numarul unitatilor dintr-un cantec este de 11,33±4,846268. Maximum de unitati intr-un cantec inregistrat, in probele noastre, este de 25, foarte ridicat avand in vedere ca si emisiile sonore au durata lunga. Cel mai mic numar de unitati intr-un cantec a fost de 5, spre finalul perioadei de reproducere cand emisiile sonore sunt scurte, slabe si cu pauze destul de lungi, intermitente. Numarul tipurilor este de 3,11±0,600895. In niciunul din cantece nu am gasit numarul maxim (5) de tipuri de unitati in analiza cantecului. Rezultatele noastre (tabel 9) concorda cu cele din literatura de specialitate (Cramp S., 1992). Repertoriul vocal la Acrocephalus scirpaceus nu este foarte complex in ce priveste tipurile de sunete emise. Complexitatea cantecului rezida mai ales in alternanta randomizata de unitati sonore diferite, urmata de repetitii ale aceleeasi unitati, pauze si reluari de ansamble de sunete.
Tabel 9: Analiza descriptiva a cantecelor de lacar de stuf
|
Statistici |
Durata |
Nr unitati/cantec |
Nr. tipuri/cantec |
|
Nr. obs. | |||
|
Media | |||
|
Eroarea standard | |||
|
Mediana | |||
|
Amplitudine | |||
|
Minimum | |||
|
Maximum | |||
|
Er. std.x t(0,05,8) |
Corelatia observata (Fig. 40) intre numarul de unitati intr-un cantec si durata sa este ridicata 88%. Cu cat durata cantecului este mai mare cu atat numarul unitatilor este mai mare. Intre durata cantecului si numarul de unitati utilizate, ca si in cazul cantecului de la lacarul mare nu am observat o corelatie semnificativa. (R2=0,23). Asta inseamna ca structura, mai mult sau mai putin complexa a cantecului, depinde de alternanta variata a unitatilor dintr-un cantec si nu din numarul acestor unitati.
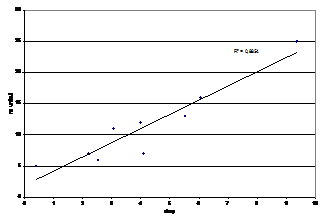
Fig. 40: Corelatia intre numarul unitatilor dintr-un cantec si durata la lacarul de stuf
Pentru a contura o imagine generala asurpra cantecelor speciei Acrocephalus scirpaceus am realizat un grafic al spectrului mediu cu interval de confidenta de 95%. Pentru fiecare cantec am extras 163 de frecvente cu amplitudinile relative corespunzatoare. La fiecare frecventa in parte am calculat media amplitudinilor relative procentuale pentru cele noua cantece. Amplitudinile relative procentuale redau in cazul fiecarei frecvente obtinute o imagine echivalenta si proportionala a tariei sunetului.
La frecvente foarte mici, aproape de 0 KKz- frecventa fundamentala, amplitudinile relative procentuale (intensitatea sunetului) sunt ridicate. In intervalul de frecvente 0,2-3 KHz procentul amplitudinal scade si are valori scazute. O usoara crestere se observa si in dreptul frecventei de 3 KHz. La frecvente mai mari de 3KHz amplitudinea relativa procentuala scade. (Fig. 41)
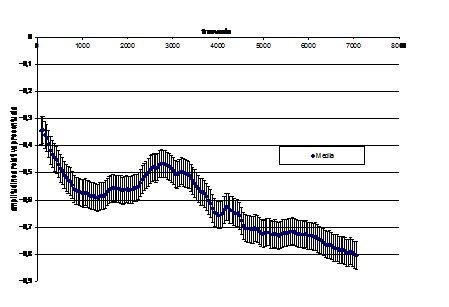
Fig. 41: Spectrul mediu al cantecelor la lacarul de stuf
In cazul lacarului mic- Acrocephalus schoenobaenus analiza cantecului s-a restrans doar la evidentierea unor unitati. Inregistrarile prelucrate au fost putine. Majoritatea cantecelor inregistrate au fost perturbate de zgomotul produs de vant.
Cantecul la Acrocephalus schoenobaenus este viguros, destul de rapid in succesiunea unitatilor. Timbrul sonor este mai melodios (Cramp S., 1992) ca la speciile precedente. In cadrul analizei cantecului se observa o repetitie metronomica a unitatilor (Cramp S., 1992), cu oarecare diferentiere spre final.
In analiza amplitudinii relative se observa existenta a patru faze mari intre care se interpun pauze scurte. Prima fraza este alcatuita la randul ei din doua subansamble. Partea de inceput este scurta, pentru ca apoi timbrul cantecului sa fie mai lung, compus dintr-o serie de vibratii diferite ca amplitudine.
Din relatia: inaltime sunet, timp si intesitatea cu care este transmis am determinat la Acrocephalus schoenobaenus sapte tipuri de unitati (silabe), diferite ca forma, intindere, interval de frecventa, energie de emitere. Analiza noastra corespunde in linii mari cu cea efectuata asupra cantecului de avertizare la adult (Cramp S, 2002). (Fig. 42)

Fig. 42: Spectograma unui cantec de lacar mic (in spectorama nu este redat si sunetul de tip F)
Unitatea de tip A are o durata mare, aproape 250 ms. Energia sunetului se contureaza cel mai bine in jurul frecventei de 5-6 KHz. In afara acestui interval de frecventa intensitatea emisiei sonore inregistreaza o curba descendenta. (Fig. 43, 44)
Fig. 43: Spectograma unitatii de tip A la lacarul mic
Fig. 44: Spectrul amplitudinii relative a unitatii de tip A la lacarul mic
Unitatea de tip B este foarte scurta, aproximativ 50 ms. Intesitatea sunetului se concentreaza pe un interval mare de frecventa 2-7 KHz. Amplitudinea relativa este ridicata cu inaltimi mari ale emisiei sonore, 7 KHz. Este cea mai ridicata frecventa a vreunei unitati la care amplitudinea este crescuta, constatata de noi. Intensitatea maxima a sunetului de tip B se evidentiaza in jurul frecventelor de 4,2-4,3 KHz. (Fig. 45, 46)
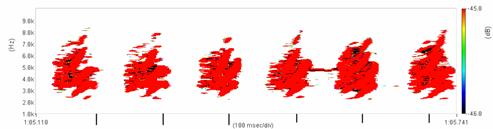
Fig. 45: Spectograma unitatii de tip B la lacarul mic
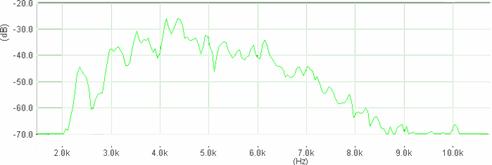
Fig. 46: Spectrul amplitudinii relative a unitatii de tip B la lacarul mic
Unitatea de tip C este cea mai lunga la Acrocephalus scirpaceus, peste 300 ms. Este un sunet subtire ascutit, cu un spectru amplitudinal maxim, la fel ca si in cazul unitatii de tip B , la 4,2-4,3 KHz. Caracteristica este aceea ca energia sunetului se resimte, chiar daca foarte slab si la frecvente de peste 8 KHz. (Fig. 47, 48)
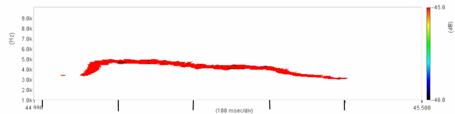
Fig. 47: Spectograma unitatii de tip C la lacarul mic
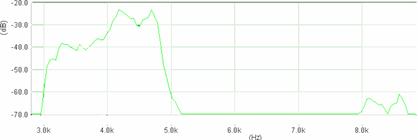
Fig. 48: Spectrul amplitudinii relative a unitatii de tip C la lacarul mic
Silaba (unitate) de tip D este de o tonalitate groasa, joasa. Durata sa este aproximativ 100 ms. Energia maxima a emisiei sonore se distribuie pe mai multe varfuri ale inaltimii sunetului: 3 KHz, 4,5 KHz, 5,1 KHz. (Fig. 49, 50)

Fig. 49: Spectograma unitatii de tip D la lacarul mic
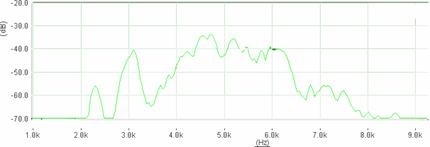
Fig. 50: Spectrul amplitudinii relativea a unitatii de tip D la lacarul mic
Sunetul de tip D este foarte asemanator cu cel de tip D. Pe langa faptul ca energia sunetului se concentreaza in mai multe varfuri de frecventa, forma spectrogramei este aproape aceeasi. Caracteristic este faptul ca silaba se compune din doua parti, intre care intervine o pauza scurta, de 10 ms, insesizabila pentru urechea umana. Desi amplitudinea este distribuita pe mai multe varfuri de frecventa se observa ca valorile intensitatii sunt mai scazute si ca panta spectrului amplitudinii relative este slaba. (Fig. 51, 52)
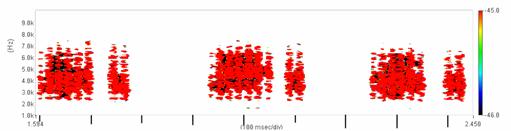
Fig. 51: Spectograma unitatii de tip D la lacarul mic
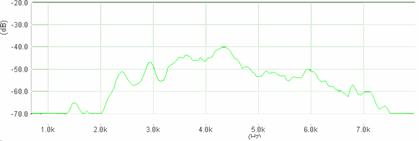
Fig. 52: Spectrul amplitudinii relative a unitatii de tip D la lacarul mic
Sunetul de tip E este mai lung si subtire asemanator celui de tip C. Durata de emisie, este mai scurta. Amplitudinea relativa atinge un maximum destul de ridicat in comparatie cu celelalte tipuri de unitati, aproape -20dB, in apropierea frecventei de 4 KHz. (Fig. 53, 54)
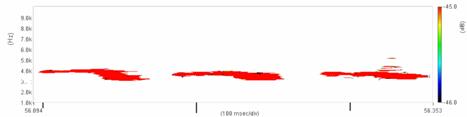
Fig. 53: Spectograma unitatii de tip E la lacarul mic
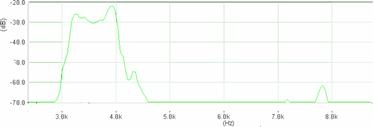
Fig. 54: Spectrul amplitudinii relative a unitatii de tip E la lacarul mic
Sunetul de tip F are aproape aceeasi durata ca si cel de tip B, adica de 50 ms. El a fost evidentiat numai spre finalul cantecului din perioada de reproducere. Energia sunetului este impartita pe mai multe varfuri de frecventa foarte apropiate ca valoare. Intervalul de frecventa in care se manifesta energia maxima este 3-6 KHz. (Fig. 55, 56)

Fig. 55: Spectograma unitatii de tip F la lacarul mic
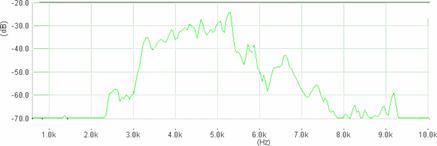
Fig. 56: Spectrul amplitudinii relative a unitatii de tip F la lacarul mic
In lucrarea de fata au fost surprinse aspecte indeosebi ale etologiei lacarului mare- Acrocephalus arundinaceus, de la migratia de primavara pana in cea de toamna. Comportamentul lacarului mare a fost comparat cu cel al lacarului de stuf- A. scirpaceus si cu cel al lacarului mic- A. schoenobaenus, la care s-au facut numeroase referiri..
Lacarul mare a fost cel mai bine studiat, intrucat aceasta specie este cel mai bine reprezentata (Ion C., 2003) in stufariile din zonele umede ale Moldovei, dar si in Delta Dunarii unde au fost intreprinse numeroase deplasari, si pentru ca miscarile si actiunile sale sunt mai usor de observat in conditii naturale.
Lacarul mare- Acrocephalus arundinaceus este specia cea mai prezenta dintre lacari in zonele umede ale Moldovei (Ion C., 2004). Comportamentul se caracterizeaza in linii mari prin indrazneala, curaj, activitate intensa s.a.
Aspectele privind comportamentul la lacari ridica si azi o serie de dificultati atat din pricina problemelor privind identificarea sexelor si a speciilor precum si pentru ca activitatea lor din interiorul stufariilor este greu de observat. Lacarul mare nu face exceptie, insa din cauza taliei mai mari si a faptului ca activitatile sale sunt mai usor de sesizat, am incercat ca pe parcursul a patru ani de zile 2000- 2004 sa adunam o serie de date care sa ne permita conturarea unei imagini cat mai complete a etologiei acestei specii.
Etograma speciei a fost alcatuita urmarindu-se principalele sezoane din cursul vietii in cartierele de vara: migratia de primavara, delimitarea teritoriului, formarea cuplului, depunerea oualor si ponta, incubatia, puii la cuib, emanciparea (cresterea si zborul) puilor si migratia de toamna.
Migratia de primavara
Lacarii mari apar in teritoriile de cuibarit din Moldova la jumatatea lunii aprilie, inceputul lunii mai, cel mai tarziu. Ei fac parte din categoria migratorilor de distanta lunga (Valcu M., 2003), venind din cartierele de iernare situate la sud de Sahara, tocmai din Africa de Sud. Populatiile din estul Europei, din ssp. arundinaceus, ierneaza mai ales in Africa de sud-est (1997- The EBCC Atlas of European Breeding Birds). Doar atunci cand vremea se incalzeste mai devreme, si disponibilitatile de hrana sunt abundente, lacarii mari pot veni in migratie si la sfarsitul lunii martie, in cartierele de cuibarit din zonele umede ale Moldovei. (Primul lacar mare, in anul 2004, a fost semnalat pe 12.04. la Larga Jijia- Vladeni).
Grosul populatiei se instaleaza in a doua decada a lunii aprilie. Observatiile efectuate in Delta Dunarii de catre noi au evidentiat ca deja la sfarsitul lunii aprilie, inceputul lunii mai migratia la lacarul mare inceteaza. In aceasta perioada incepe sa se formeze stuful nou. Observatiile noastre au reliefat, de altfel ca lacarii nu-si construiesc cuibul decat pe stuful nou. Lacarii mari- Acrocephalus arundinaceus, prefera stuful mai inalt, de peste 2,50 m, pe cand celelalte specii de lacari isi ocupa teritorii adesea in stufarii de talie mica.
Acesti lacari in migratia de primavara se aduna in grupuri de 4- 12 indivizi, in cordoanele de stuf. In aceasta perioada nu apare comportamentul luptei pentru teritoriu. Ei cauta hrana, pentru a-si reface rezervele adipoase necesare inceperii perioadei de reproducere. Miscarile pentru cautarea hranei sunt realizate pe distante scurte, 20- 30 m si mai ales in apropierea suprafetelor compacte de stuf. Salturile de pe un fir de stuf pe altul sunt scurte si la o inaltime destul de mica de sol sau de apa. Comportarea lor este retinuta. Miscarile sunt facute cu precautie. Lacarii nu cauta, inca sa-si faca simtita prezenta iar trilurile lor sunt scurte si de o intensitate scazuta.
Delimitarea teritoriului
La aproximativ 5-10 zile de la sosire, in aprilie, isi instaleaza teritoriul de cuibarit pana la sfarsitul lunii aprilie, inceputul lunii mai. Mai intai ajung masculii si apoi femelele (Cramp S., 1992). In general acestea apar la 9- 11 zile in teritoriile de cuibarit, conform observatiilor noastre. La jumatatea lunii mai, in sudul Moldovei si la sfarsitul lunii mai, in centru si in nord-est, din constatarile noastre, teritoriile de cuibarit sunt delimitate.
In general la populatiile de lacar mare se constata o fidelitate ridicata pentru locurile de cuibarit atat la adulti cat si la juvenili (Hasselquist D., 1998). In schimb masculii care nu inregistreaza succes intr-un anumit teritoriu nu se mai intorc in acel loc in anul urmator. (Bensch si Hasselquist, 1991 citat de Hasselquist D, 1998)
Stabilirea teritoriului nu incepe, din observatiile noastre, prin evaluarea greutatii corporale (Ion C., 2004) ,inainte de refacerea stratului de grasime ce le asigura energia necesara zborului si inceperea perioadei de reproducere, deoarece dupa ce isi delimiteaza teritoriul, lacarul mare nu-l mai paraseste pentru o perioada ce variaza intre 10 si 20 de zile.
In perioada delimitarii teritoriului, activitatea este maxima pe tot parcursul zilei si al noptii. Rare sunt momentele de pauza si acestea apar la amiaza in zilele foarte calduroase si noaptea, mai ales in cazul unor furtuni, dupa cum am constatat. In momentul in care erau prinsi in plase ornitologice mai multi lacari mari si tinuti in saculete de panza, seara, acestia se agitau foarte mult pana la ora 20.30-21.00, cand se insereaza. Noi am considerat ca lumina este un factor declansator al activitatilor la lacarul mare. Noaptea, activitatea lor nu inceteaza, dar se limiteaza la miscari scurte si cantece de multe ori sporadice. Exista si cazuri cand lacarii sunt foarte agitati chiar si in timpul noptii. Aceasta se intampla spre sfarsitul perioadei de delimitare a teritoriului in cazul lacarilor tineri care nu-si gasesc teritorii favorabile reproducerii si hranirii.
Noi am constatat ca teritoriile pe care le stabilesc lacarii, se refera numai la acele spatii in care are loc acuplarea si cuibaritul, dar nu si hranirea.
Teritorialitatea la lacari poate fi vazuta ca o forma de agresiune si competitie, cand intre congeneri exista o concurenta sexuala, dar si una pentru resurse de mediu, ce include posibilitatea de a se hrani mai usor si a gasi un adapost propice.
In timpul stabilirii teritoriului, lacarul mare zboara si canta cel mai adesea din interiorul teritoriului de cuibarit spre granitele acestuia, dupa care se reintoarece in apropirea centrului. Centrul teritoriului reprezinta in cele mai multe cazuri locul unde isi va instala cuibul viitoarea pereche. Noi am observat ca lacarul canta uneori perioade lungi de timp in aceasta perioada, peste 10 minute, aproape fara incetare. Pentru a canta, masculii se sprijina pe varful firelor de stuf. Cantecele lor sunt formate dintr-o juxtapunere de motive care se repeta si alterneaza intr-o succesiune rapida. Cantecul are rol nu numai in atentionarea congenerilor cu privire la delimitarea teritoriului dar si la atragerea femelei in vederea reproducerii. Cantecul reprezinta in acest caz un mijloc de marcare acustica, fiind un indiciu al fortei. Prin intermediul cantecului rivalii sunt informati daca este sau nu cazul sa se angajeze intr-o confruntare teritoriala. Acustica etalata de lacarul mare in perioada delimitarii teritoriului are o mare importanta, dovada fiind faptul ca in cazul unor vanturi puternice, 4- 5 pe scara Beaufort, uneori insotite de ploaie, ei nu inceteaza sa cante. Dimpotriva amplitudinea sunetelor creste, si chiar incearca sa se urce pe varful firelor de stuf, insa de cele mai multe ori fara succes, intrucat taria vantului ii obliga sa coboare mai jos. Aceasta este o dovada, in plus, a devotamentului pe care il manifesta fata de spatiul ocupat.
Lacarul mare isi marcheaza teritoriul atat acustic cat si vizual. Marcarea optica a teritoriului are loc prin afisarea unei posturi care da impresia de corp mare, mai mare decat cel pe care il are in realitate: penajul infoiat, coada si aripile desfacute.
De multe ori se opresc la marginea teritoriului si canta. In tot acest timp sunt orientati cu capul spre interiorul teritoriului ca si cum ar vrea sa vada daca nu apar intrusi. Uneori reprizele de cantec sunt intrerupte deoarece lacarul mare isi curata penajul. Probabil ca acest gest reprezinta un mijloc de distragere a atentiei celor ce vor sa-i incalce teritoriul.
In unele cazuri s-a vazut ca masculul celibatar nu-si delimiteaza neaparat un singur teritoriu la inceput, ci cauta mai multe locuri pentru a canta si a-si delimita o arie. Cu alte cuvinte ei cauta sa obtina mai multe oportunitati de a-si gasi o femela in vederea reproducerii ceea ce sporeste cu mult sansele de reusita.
Cand doi lacari canta in doua teritorii invecinate, fiecare dintre ei, sta, de obicei in centrul propriei arii. In cazul in care unul din ei manifesta mai mult curaj, se apropie foarte mult de granita propriului teritoriu. Situatia aceasta se poate inversa insa de mai multe ori pe parcursul unei zile. Lacarii se departeaza unul de celalalt, fiecare cantand din propriul teritoriu, fara a face pauze lungi. Pozitia lacarului mare in tot acest timp arata astfel: capul orientat in sus, penele de pe cap putin zburlite, tectricele de pe gusa sunt ridicate, de un alb imaculat, pieptul putin proiectat in fata, iar aripile sunt usor desfacute, aduse in fata, coada usor desfacuta in evantai. In opinia unor specialisti (McFarland D., 1999), o astfel de postura de amenintare, reprezinta un avantaj in luptele dintre congeneri. Nu de putine ori am vazut ca acestia se apropie unul de celalalt la distante foarte mici, de numai 2- 3 m. In tot acest timp cantecul lor este etalat cu o frecventa si intensitate ce se accentueaza pe masura apropierii, celor doi, unul de celalalt. Pozitia celor doi este una de amenintare, cu penele de contur zbarlite, aripile ridicate, coada desfacuta in evantai. Timbrul vocal al cantecelor poate suferi modificari in astfel de situatii. Combatantii emit franturi de cantec compuse dintr-un numar mic de motive vocale. Uneori are loc un contact fizic intre cei doi, de scurt timp, ciocnindu-se usor cu pieptul. Dupa aceea fiecare lacar se reintoarce in interiorul teritoriului sau. In alte situatii, masculul mai puternic ramane la granita teritoriului pe varful firelor de stuf, iar cel de-al doilea mascul se retrage in interiorul suprafetei de stuf.
Lacarii din teritorii contigue, realizeaza in zilele calduroase zboruri introspective la baza stufului dupa care se reintorc in varful firelor de stuf si canta triluri lungi de cateva secunde cu pauze foarte scurte si cu o intensitate ridicata. In momentul in care unul dintre lacari se ridica pe varful firelor de stuf si celalalt din teritoriu vecin se va urca la capatul tulpinilor de de stuf si isi va expune partitura vocala. Cantecul lor nu este emis in acelasi timp, ci alternant, cand unul, cand celalalt, in numeroase cazuri se confirma observatiile efectuate de noi. La lacul Gradinii Botanice din Iasi am auzit, in mai multe zile la rand (25-29 Mai- 2004) chiar si trei lacari cantand in trei teritorii contigue, cantand pe rand, fiecare cam intre 1-2 minute. Foarte rar, lacarii canta in acelasi timp, in teritorii invecinate.
Intre teritoriile stabilite de lacari nu exista hiatusuri. Spatiile ocupate de lacarii mari, in sezonul de cuibarit sunt contigue. Cu toate acestea in stufarii intinse de sute de ha (Larga Jijia, Beresti, Iezar, Balta Lata, Vladesti, Carja) lacarii mari formeaza adevarate colonii cu teritorii bine delimitate, ce nu se intrepatrund. Intre aceste colonii exista spatii ocupate de alte specii de pasari (rate, chire, pitigoi de stuf, presure de stuf, ciconiiforme acvatice s.a.) sau de intinderi de apa, fara stuf. Insa in cadrul suprafetelor umede exista si zone de stufaris mai putin populate de lacarii mari. Uneori celelate specii de paseriforme de stuf formeaza aglomerari relativ mari (5-10 indivizi) care isi apara in comun teritoriile.
Teritoriile au o suprafata de de la 100 m2 pana la 250 m2, acolo unde intinderile de stuf sunt mici in zonele umede ale Moldovei (spre exemplu Gradina Botanica Iasi). Pe suprafata stufariilor de la Larga Jijia sau Iezar- Dorohoi, teritoriile de cuibarit au suprafete mult mai mari intre 1000 m2 si 5000 m2.
In cazul benzilor de stuf inguste, de 20- 40 m latime, cum este cazul stufariilor de la Gradina Botanica- Iasi, sau cele ce marginesc digurile de separe ale lacurilor la Vladesti, Carja, ori Larga Jijia, teritoriul lor poate fi foarte mic de numai 50- 100 m2, in cazul in care abundenta lacarilor este foarte mare. Un numar mare de lacari apare primavarile cu vegetatie bogata si abundenta ridicata de insecte. In sezoanele de cuibarit 2003 si 2004 la inceputul primaverii a plouat foarte mult si vegetatia s-a dezvoltat foarte repede. Dupa aparitia vegetatiei au aparut si insectele in numar mare mai ales in momentul cand temperatura aerului a crescut si soarele a stralucit indelung.
Delimitarea teritoriilor la lacari este in stransa dependenta cu conditiile meteorologice din mediul in care traiesc. Teritoriile mici, in cazul lacarilor, din observatiile noastre apar in primaverile foarte caldurose, destul de ploioase, cand in luna mai (anii 2003, 2004), temperaturile, au depasit 20 0C. In anii cu temperaturi scazute (primavara 2001), sunt cazuri, in care, in unele zone restranse ca intindere (exemplu Gradina Botanica, 2001), lacarii apar pe la sfasitul lunii mai si cuibaresc pe teritorii foarte vaste, de aproape 2500 m2. In opinia noastra temperaturile crescute, lipsa precipitatiilor sunt factori nefavorabili pentru cuibaritul lacarului mare. De asemnea, temperaturile foarte scazute , intarzie delimitarea teritoriilor, inceperea formarii perechilor. In anul 2001 la Gradina Botanica la sfarsitul sezonului de cuibarit am observat foarte putine exemplare juvenile (la 7 perechi clocitoare, 10 indivizi juvenili), ceea ce inseamna ca rata succesului, in acest caz a fost destul de scazuta. Drycz, 1995, afrima ca subspecia lacarului mare prezenta in estul Europei (ssp. arundinaceus), tolereaza un spectru ingust al conditiilor de mediu si de areal, ceea ce vine in completarea afrimatiilor noastre.
Ariile de cuibarit sunt rar stabilite spre interiorul stufariilor, intrucat nu ofera avantaje pentru gasirea locurilor de hranire. Primii sositi, conform datelor noastre, ocupa intotdeauna stufariile care au deschidere atat inspre suprafata de apa cat si spre maluri. Indivizii ce sosesc mai tarziu incearca sa recupereze terenul pierdut prin cantece foarte puternice si miscari ample, de atragerea a atentiei: survolari cu remigele primare puternic rasfirate, coada desfacuta in evantai si usor ridicata, penele zburlite. Teritoriile pe care le ocupa acestia se situeaza in interiorul stufariilor, sau in locuri mai umbroase, unde oportunitatile de hranire sunt slabe, intrucat numarul de insecte este foarte mic.
Daca la marginea intinderilor cu stuf, teritoriile au o forma neregulata spre interior si mai ales intre teritoriile invecinate a doi lacari aceste teritorii urmeaza o linie imaginara aproape dreapta. Cu toate acestea teritoriile nu trebuie imaginate ca niste zone cu granite fizice fixe. Suprafata lor se poate schimba in timp si depinde de comportamentul teritorial (notiune mentionata de Cociu M., 1999) manifestat de lacar.
De altfel trei factori sunt cei ce favorizeaza teritorialitatea: calitatea resurselor, distributia lor in spatiu si timp precum si competitia pentru resurse (McFarland D., 1999). Zonele umede ca: Gradina Botanica, Beresti, sarace in resurse ofera conditiile unei competitii intraspecifice accentuate pentru resursele de hrana, de accea teritoriile sunt mici, iar uneori granitele sufera modificari.
Dupa stabilirea teritoriilor, lacarii mari survoleaza pe deasura teritoriului, ori la suprafata apei cercetand terenul. In cazul aparitiei unui intrus, acesta este imediat alungat si urmarit in afara teritoriului uneori pe distante destul de lungi, de 50-100 m. Noi am observat ca in tot acest timp ocupantul teritoriului ia o pozitie de amenintare. Penele de pe cap si uneori si cele de pe spate sunt usor ridicate. Intrusul are penajul putin ridicat, dar renunta repede la pozitia de lupta. Rareori am observat contacte fizice intre "proprietar" si intrus. De obicei stapanul teritoriului alunga ceilalti lacari fara nici un fel de dificultati. Patrunderea congenerilor in teritoriile stabilite nu sunt rare si sunt realizate, cel mai adesea fara etalarea cantecului specific din partea intrusilor.
Noi am observat ca exista cazuri cand ocupantul unui teritoriu la aparitia unui concurent in spatiul sau, executa zboruri verticale, emite sunete puternice de alarma, coada si aripile sunt usor coborate, si de la aproximativ 3- 4 m se repede asupra celuilalt lacar. Intrusul de obicei sta undeva in partea de jos a stufariilor, cercetand terenul. In momentul atacului de cele mai multe ori nu riposteaza cu ciocul sau cu picioarele, ci este nevoit sa plece.
Datele din literatura (Cramp S., 1992) arata ca primii care sosesc in teritoriile de cuibarit sunt indivizii mai in varsta de doi, trei ani, care in multe cazuri sunt poligami. Exemplarele tinere, vin mai tarziu, si ocupa habitate mai putin favorabile hranirii si construirii cuiburilor. Ei sunt monogami sau nu-si gasesc pereche in vederea reproducerii. Cu alte cuvinte, lacarii in varsta au o rata a succesului reproductiv mai ridicata decat cei tineri. Noi am observat ca rareori lacarii care au ocupat prima data o arie din stufarii sunt alungati. Pe parcursul anului 2001 am constat trei cazuri, in luna mai, la Larga Jijia, in care exemplare noi venite de lacari au ocupat teritoriile altor indivizi mai vechi. Luptele rituale au fost de scurta durata. Indivizii noi veniti aveau un zbor tremurat, rapid, penele de pe coroana capului zbarlite si se asezau pe varful firelor de stuf etalandu-si cantecul. Din cauza penajului mai putin uzat si a zburlirii penelor pareau mai mari decat ocupantii acelor teritorii, corpul era aproape sferic. Cel alungat fie, isi lua zborul intr-o alta directie, fie se retragea in interiorul vegetatiei. Disputele teritoriale in acest caz sunt destul de pasnice. Semnalul pe care il da cel invins consta din aspectul penajului. El are penajull netezit, coada nedesfacuta, pare mai mic ca lungime si volum.
In opinia noastra suprematia aratata de un lacar sau altul se rezuma la etalarea unor cantece complexe si a unei posturi intimidante.
Formarea cuplului
Schimbarile sezoniere climatice in privinta temperaturii si a precipitatiilor exercita influente puternice in ceea ce priveste sezonul reproductiv. Printre pasari, disponibilitatile de hrana pentru pui par sa fie cauzele determinante in declansarea sezonului reproductiv. (Lack, 1968 citat de McFarland D., 1999).
Literatura de specialitate mentioneaza ca lacarii sunt monogami, dar simultan sau succesiv pot fi poligami (Cramp S., 1992). Au fost semnalate si cazuri de promiscuitate. Noi nu am constat, in zonele umede studiate ale Moldovei cazuri de poligamism ori promiscuitate.
In cadrul comportamentului de formare a cuplului intreaga desfasurare a activitatii sexuale este o succesiune strans orchestrata de comportamente ce difera radical in privinta formei si a functiei ramanand in acelasi timp canalizata catre actul unic al fertilizarii. (Wilson E., 2003). Pot fi distinse cel putin cinci asemenea clase de actiuni: parada sexula, curtarea, legatura sexuala, comportamentul copulator si manifestarile post- copulatorii.
Noi am observat ca parada sexuala, in cazul lacarului mare, se refera la semnalizarea prezentei de catre mascul prin cantec si postura. Curtarea pentru atragerea femelei are loc prin diversele forme de comportament. Masculul aduce insecte sau material vegetal in cuib. Cantecele emise sunt deosebit de elaborioase, alternand motive care semnifica chemarea cu cele care dau semnalul de alarma. Masculii isi etaleaza penajul mult infoiat si culoarea rosie a cavitatii bucale. Legatura sexuala se formeaza in momentul cand femela nu mai este deranjata de mascul si ii accepta acestuia prezenta. Comportamentul copulator si cel post- copulator se intrepatrund cu constructia cuibului si furnizarea de hrana la cuib.
Formarea cuplului are loc, in general, in zonele umede din Moldova la sfarsitul lunii mai, inceputul lunii iunie.
Sosirea femelelor este urmata la scurt timp de formarea perechilor si de acuplare (luna mai). Femelele cand ajung in teritoriile de cuibarit nu-si aleg imediat partenerul. Noi am observat ca ele se aduna mai multe la un loc in suprafete compacte de stuf, unde gasesc refugiu si de unde pleaca spre marginile stufariilor pentru a-si cauta hrana. Abia la 5-6 zile dupa venire, cand incepe sa li se formeze placa incubatoare si dupa ce isi refac fortele pornesc in cautarea unui partener.
Lacarii, in ceea ce priveste inceperea sezonului de cuibarit respecta principiul lui Bohning G. (citat de Valcu M., 2003) conform caruia migratorii de distanta lunga isi incep sezonul reproductiv mai tarziu si au o rata reproductiva mai scazuta ceea ce sugereaza ca acestia au o rata de mortalitate mai scazuta comparativ cu migratorii de distanta scurta si cu populatiile rezidente. Astfel specii ca Sturnus vulgaris sau Passer domesticus la sfarsitul lunii mai au deja o a doua ponta. Uneori populatiile rezidente din zonele umede ale Moldovei au si pana la trei ponte intr-un an. La lacarii din zonele umede ale Moldovei nu am observat cazuri in care femelele sa aiba o a doua ponta.
Femela este cea care isi alege partenerul. O mare importanta o are calitatea habitatului (Cramp S., 1992), in ceea ce priveste marimea sa, disponibilitatile de hrana pe care le ofera. Noi am observat ca masculul de lacar mare incepe deja constructia unui cuib, pe care femela il cerceteaza. Pana la venirea femelei, masculul aduna material de cuib, din interiorul stufului si la intervale de 5- 10 minute se urca in varful firelor de stuf cercetand terenul din imprejurimi.
Ulterior, definitivarea cuibului va fi continuata de ambii parteneri. Femela care patrunde in interiorul unui teritoriu nu se intereseaza in mod direct de proprietarul acestuia, ci de locul instalarii cuibului, mai intai.
In incursiunile noastre in teren am constat ca masculul incearca sa atraga femela mai ales prin etalarea unor cantece "de chemare", cu triluri lungi, complexe cu amplitudine si intensitate crescute. In cazul aparitiei unei femele, masculul realizeaza zboruri circulare in jurul acesteia, cu penajul infoiat. Femela in tot acest timp, conform observatiilor noastre de la sfarsitul lunii mai din Gradina Botanica- Iasi, poate emite cantece usor sacadate, de alarma, in interiorul suprafetei de stuf, fara a putea fi observata vizual. Masculul ramane la aproximativ 10-20 m de ea si nu coboara de pe varful firelor de stuf.
In momentul apropierii femelei, masculul pare a-si schimba tonalitatea cantecului, fiind mult mai tremurat, mai putin amplu. Pozitia capului se mentine intr-un unghi de 450 fata de sol, picioarele putin curbate, pentru a se putea mentine pe varful firelor de stuf. Penele de contur sunt zburlite, semn al excitarii de care este acesta cuprins. La intervale de 10- 20 de minute, masculul patrunde in interiorul stufului, cu salturi scurte pe firele de stuf si continua sa cante, dar nu cu aceeasi intensitate ca la inceput, cercetand femela. Dupa aproximativ 5 minute se reintoarce in aproape acelasi loc si continua sa cante cu tarie.
Studiul privind atenuarea efectelor sonore a aratat ca folosirea varfului firelor de stuf ca punct de sprijin asigura o penetrare sonora optima (Jilka si Leisler, 1974, citat de Cramp S., 1992).
Apoi am constat ca zborurile masculilor sunt scurte sub forma literei Z, se ridica pe varful firelor de stuf, explorand imprejurimile. La revenirea deasupra stufului cantecul masculului ajunge din nou la o intensitate foarte ridicata.
In mai multe cazuri, la o zi sau doua, cei doi potentiali parteneri se apropie unul de celalalt la distante mici pentru scurt timp, dupa care se departeaza. Uneori are loc un contact usor intre cei doi prin ciocurile lor.
Contactul ciocurilor la pasarilor are functii multiple centrate pe stabilirea si mentinerea legaturilor (Wickler, 1972, citat de Wilson E, 2003). In cazul descris anterior contactul dintre ciocuri semnifica acceptarea de catre femela a masculului. Astfel, pe langa comunicarea vizuala, apare si comunicarea tactila cu un rol deosebit in imperechere. La Gradina Botanica- Iasi si Larga Jijia in stufarii am observat ca masculul executa salturi inainte si inapoi pe firele de stuf, pe cand femela realizeaza mici salturi in partea de jos a stufului. Dupa aceasta masculul se retrage putin catre marginea teritoriului, catre mal, iar partenera vine putin catre el dupa care se intoarce la prima sa pozitie. Ambii parteneri executa miscari prin care se apropie unul de celalalt si au posturi ce sunt directionate unul spre celalalt.
In mai multe situatii am observat ca intre cei doi parteneri are loc o adevarata "joaca" rituala. Ei se apropie unul de celalalt, se privesc pentru cateva clipe, dupa care se fugaresc unul pe celalalt. In momentul apropierii cei doi se lovesc, uneori usor unul de celalalt cu piepturile. Masculul are coada ridicata, aripile desfacute, penajul zburlit, pe cand la femela penajul arata mai putin spectaculos. Coada ei este desfacuta in evantai si coborata, aripile aduse in fata si usor curbate, penajul mai mult neted, doar cateva pene de contur de la gat si de pe coroana capului sunt desfacute, ciocul este putin deschis intr-o pozitie de amenintare.
In cadrul comportamentului de curtare, lacarul mare executa zboruri rituale in timpul caruia sunt expuse cu maxim de avantaje modele speciale ale penajului. Astfel in multe cazuri, masculul expune in directia femelei albul de pe spatele aripii si al cozii. Culoarea alba alterneza cu ruginiul tartitei in zborul remarcabill pe care-l executa. Apoi inainte de a se ajeza pe varful stufului ridica aripile aproape vertical si tremura din varful lor. Toate aceste posturi si miscari sunt menite sa atraga femela in vederea acuplarii.
Daca in aceasta perioada in interiorul teritoriului apare un congener, ambii parteneri pot participa la alungarea lui. Uneori intrusul este ciupit de dupa gat sau de cap.
In primele faze de formare a cuplului cei doi parteneri manifesta foarte multa retinere chiar uneori agresivitate mai ales din partea femelei. De altfel, Cociu M., 1999, mentioneaza ca: "excitatia sexuala si tendinta de apropiere, pe de o parte, teama de partener, agresivitatea si tendinta de evitare, pe de alta, interactioneaza frecvent si in grade variabile, generand un surplus de motivatie ce se canalizeaza, de regula, prin comportamente specifice starilor conflictuale."
La Iezar si Larga Jijia am observat ca uneori femela isi alunga partenerul din teritoriu cand acesta se apropie prea mult, emitand sunete de alarma cu o durata de 2-3 secunde. Ea adopta o pozitie de amenintare fata de mascul la inceputul perioadei de formare a cuplului.
Masculul recurge si la alte strategii pentru a-si atrage femela in vederea acuplarii. Ii aduce acesteia insecte in cioc. Chiar si asa femela nu accepta de la inceput prezenta masculului. (fig. 9) Masculul in acest caz nu paraseste teritoriul mai mult de 2- 3 minute. Apropierea de femela se realizeaza din mai multe directii. Emite sunte de chemare, etaleaza un tremurat din aripi, coada este usor desfacuta. Femela, emite sunete scurte ascutite, de alarma, si isi cerceteaza foarte atent partenerul.
Masculul, l-am vazut in mai multe cazuri carand material de cuib in cioc, din afara teritoriului de cuibarit. Un semn al acceptarii il constituie preluarea firelor de material vegetal de catre femela direct din ciocul masculului asa cum de altfel se mentioneaza si in literatura de specialitate (Cramp S., 1992).
Dupa formarea perechii constructia cuibului este continuata de ambii parteneri. Femela este insa cea care va definitiva constructia. Masculul o insoteste in zborurile ei si o ajuta la cararea materialului pentru cuib. Totusi miscarile ei sunt pe distante scurte, pe cand masculul executa zboruri mai lungi, la distante de peste 100 de metri.
Adeseori, femela zboara in urma masculului si il urmareste pe acesta in interiorul teritoriului, pentru a-i cunoaste granitele. In acest timp cantecul masculului aproape ca inceteaza, semn al faptului ca formarea cuplului s-a definitivat.
In cazul formarii cuplului, masculul are penajul usor infoiat, pe cand femela are penele netezite. El emite in tot acest timp sunete de chemare.
Femela apare in apropierea cuibului si-l indeparteaza usor cu ciocul pe acesta continuandu-si constructia cuibului. Femela etaleaza un comportament mai mult agresiv fata de mascul, nu-i accepta in totalitate prezenta si apropierea. Emite scurte sunete de alarma cand acesta se apropie. Uneori executa zboruri in care il alunga pe mascul de la cuib. Desi agresiva, penajul femelei ramane neted. (fig.10)
Un semn al formarii cuplului il constituie inhibarea agresivitatii partenerului, linistirea lui. Femela incepe usor sa accepte prezenta masculului in preajma sa si chiar sa-i accepte ajutorul, nu il mai respinge. Coada este stransa, aripile desfacute, usor coborate, capul tinut drept inainte.
Masculul in tot acest timp, am obsevat ca hraneste femela ocupata cu constructia cuibului in interiorul stufariilor. Cantecul masculului este auzit doar dupa ce acesta isi hraneste partenera, insa cu o mai slaba intensitate. Postura masculului ramane neschimbata: aripile si coada usor ridicate, desfacute usor in evantai, penele de pe cap si gat usor zburlite. Miscarile sale sunt directionate mai ales in interiorul stufariilor si mai rar pe varful firelor de vegetatie in perioada constructiei cuibului.
Femela cere de mancare scotand franturi scurte de sunete, iar conform datelor unor etologi straini (Kluyver, 1955 citat de Cramp S., 1992) sunetele sunt insotite de batai puternice din aripi si indoiri usoare ale acestora timp de un minut sau doua.
In acelasi teritoriu de cuibarit poate apare si un al treilea individ, asa cum am observat in stufariile la lacul Gradinii Botanice. Daca stapanul canta, fie in centru teritoriului sau la marginea acestuia, intrusul se misca si el in jurul teritoriului incercand sa atraga atentia femelei care face obiectul disputei rituale dintre cei doi. Desi se incearca alungarea intrusului, acesta nu va fi alungat imediat. Stapanul renunta sa-si mai alunge congenerul si isi reia comportamentul prin care incearca sa atraga atentia femelei. Interesant ca la numai cateva ore incepe o concurenta intre cei doi masculi. Acestia canta alternant sau chiar in acelasi timp, cu penajul infoiat, coada desfacuta in evantai. Zborul in care cei doi se cerceteaza unul pe celalalt continua timp de mai multe minute. Stapanul teritoriului canta fara incetare (uneori 15-20 minute, cu pauze foarte scurte) indreptandu-si corpul in cele patru puncte cardinale si cercetand granitele propriului teritoriu. La ambii masculi se observa foarte clar rosul cavitatii bucale, culoare care se constituie intr-un atractant sexual. Femela in tot acest timp zboara in interiorul stufului emitand sunete scurte, fragmentate, neapropiindu-se mai mult de 15 m de cei doi masculi. In cele din urma, in cazul observatiilor noastre, stapanul teritoriului castiga dreptul de a se acupla cu femela. Acesta pare a avea o statura mai impozanta si cantecul de o amplitudine foarte ridicata. In cazul descris mai sus acuplarea femelei poate avea loc totusi cu individul care pare a fi pierdut lupta rituala pentru imperechere. Astfel de situatii, mentioeaza si unii ornitologi de specialitate (Cramp S., 1992, Mc. Farland D., 1999). De altfel,asa cum se va descrie in capitolele urmatoare, masculul va participa la ingrijirea cuibului o perioada de timp, purtand rolul probabil al unei doici.
Femela, in cazul in care imperecherea esueaza, il paraseste pe acesta si se indreapta in alta directie. Astfel am observat mai multe cazuri in care unul din masculi, la lacul Gradinii Botanice, dupa ce si-a delimitat teritoriul si format perechea a ramas singur ca urmare a esuarii imperecherii.
Date concrecte despre acuplare nu se cunosc. (Cramp S., 1992). In opinia noastra ea are loc in interiorul stufariilor si dureaza nu mai mult de 2-3 minute. Ambii parteneri in aceasta perioada sunt observati cu coada ridicata si desfacuta in evantai, aripile desfacute, penajul la mascul infoiat, la femela neted. Sunt si cazuri citate in literatura de specialitate cand femela se poate incrucisa cu mai multi masculi, desi isi pastreaza acelasi partener (Cramp S., 1992), pentru a asigura succesul reproductiv.
Perioada formarii cuplului dureaza, dupa observatiile noastre, la lacarul mare, intre 7 si10 zile.
Depunerea oualor si ponta
Cuiburile, in cazul stufariilor intinse, sunt stabilte mai ales catre malurile apelor, deoarece aici gasesc resurse trofice suficiente pentru intretinerea lor si a puilor. S-a observat ca cel mai adesea cuiburile de lacar trebuie sa aiba o deschidere si catre apa, ceea ce reprezinta o cale suplimentara de scapare in caz de pericol. La aparitia unui pradator, de obicei, se refugiaza catre apa realizand zboruri planate catre alte cordoane de stufaris.
Constructia cuiburilor dureaza in medie 5- 6 zile, conform observatiilor noastre.
Cuiburile sunt amplasate la 0.3-2 m de suprafata de apa. Ele sunt constituite din materiale vegetale impletite in jurul firelor de stuf. (Alcatuirea cuiburilor si dimensiunile sale au fost descrise in Referatul II, Ion C., 2004).
In cazul distrugerii cuibului, lacarii mari, in cele mai multe cazuri, nu-si mai refac cuibul. Astfel, am observat la Larga Jijia, Iezar si lacul Gradinii Botanice, dupa o furtuna, cuiburile acestora au fost distruse chiar dupa depunerea oualor. Cei doi parteneri au parasit teritoriul de cuibarit si nu s-au mai intors.
La Larga Jijia pe parcursul mai multor ani am observat mai multe cuiburi parasite ca rezultat, fie al vremii nefavorabile, vant puternic (iunie 2002), fie al esuarii acuplarii (iunie , 2001, 2002,).
In timpul depunerii oualor daca viata la cuib este deranjata de apropierea omului, lacarii nu-si parasesc cuibul si ouale. Dimpotriva recurg la diverse mijloace acustice si optice pentru a-i distrage atentia, asa cum vom prezenta in capitolele urmatoare.
Depunera oualor are loc pe parcursul a 3-6 zile conform observatiilor noastre. O femela de lacar mare depune in medie intre 2-5 oua (Fig. 60) Femela de lacar mare depune un ou pe zi. Ouale sunt asezate de obicei cu capul mai bont spre exterior.
In timpul depunerii oualor, femela nu-si paraseste cuibul decat pentru a bea apa. Masculul, dupa observatiile noastre, este cel care-i aduce de mancare. Hrana este compusa in special din coleoptere ori odonate de dimensiuni mari, care poate furniza partenerei suficiente rezerve energetice necesare depunerii oualor.
In momentul depunerii tuturor oualor, femela si masculul parasesc cuibul chiar pentru cateva ore. Ele realizeaza zboruri planate deasupra apei, in jurul cuibuilui, spre maluri, survoleaza de mai multe ori teritoriul, dupa care cauta hrana. Postura masculului este caracteristica, coada usor cobarata, stransa, aripile coborate, penajul neted si de foarte multe ori a fost observat stand in apropierea cuibului si pazindu-l.
Ambii parteneri au fost observati ulterior depunerii oualor cautandu-si hrana. Masculii realizeaza zboruri mai ales in pajistile si padurile de la marginea suprafetei acvatice. Prezenta femelei, este insa mult mai discreta ca a partenerului.
Incubatia
Dupa observatiile noastre incubatia oualor dureaza intre 10- 14 zile. Femela este cea care cloceste. Rare sunt cazurile cand cloceste masculul. Acest lucru se intampla doar in cazul masculilor monogami. (Cramp S. 1992)
In timp ce femela cuibareste, masculul furnizeaza hrana la cuib. El nu se departeaza la mai mult de 50-100 m de cuib. Observatiile noastre au reliefat ca sunt cazuri in care femela paraseste cuibul pentru mai multe zeci de minute. In acest timp, nu este inlocuita de mascul. Ea se hraneste si isi curata penajul. Ambii parteneri sunt vazuti zburand in tandem la marginea stufarisului. Pe parcursul unei zile femela isi paraseste de mai multe ori cuibul.
Dupa Kluyver citat de Cramp S., 1992 pe parcursul unei zile o femela sta pe cuib in medie 13 minute si absenteaza 31/2 minute.
La inceputul perioadei de cuibarit plecarile femelei de la cuib sunt mai rare. Cu cat se apropie momentul eclozarii, femela paraseste mai des ouale in cautarea hranei.
Noi am observat ca intotdeauna femela, cand se apropie de cuib, manifesta foarte mullta prudenta. Inconjoara de mai multe ori cuibul pe la baza stufului, dupa care se urca treptat pe firele de stuf, se urca pe marginea cuibului de mai multe ori, priveste in jos spre interiorul cuibului. Dupa aproximativ 4- 5 minute de cercetari se urca si se aseaza in cuib.
In aceasta perioada masculul canta in jurul cuibului si nu inceteaza sa-si arate prezenta. Prin aceasta el isi mentine granitele teritoriului intacte si are grija de femela si cuib in cazul aparitiei unor pradatori. Cand se departeaza de cuib, in mod normal, masculul nu scoate sunete.
Observatiile facute la Iezar- Dorohoi, au aratat ca femela poate fi inlocuita de mascul, in timpul zilei, cu toate ca el nu poseda placa incubatoare. Cand inlocuirea la cuib are loc, masculul emite scurte fraze de cantec, la care femela ii raspunde cu sunete scurte, fragmentate de o tonalitate joasa. Femela, cand revine la cuib, isi loveste usor cu ciocul masculul si uneori se aseaza deasupra acestuia, dupa care masculul paraseste, prin salturi scurte, cuibul. Acelasi ritual are loc si cand masculul isi inlocuieste partenera. In momentul cand masculul sta pe cuib femela pleaca in cautarea hranei.
In 2004 la lacul Gradinii Botanice din Iasi, am observat aparitia in preajma cuibului a unui al treilea individ. Este probabil unul dintre masculii cu care ocupantul teritoriului isi disputase cu o luna in urma teritoriul. Acesta juca rolul unei doici ceea ce insemana ca la lacarul mare apare si comportamentul de intrajutorare la cuib. Atat el cat si partenerul femelei de pe cuib furnizeaza hrana. Cand este vorba de hrana de dimensiuni mai mari, unul din ei o preia undeva la jumatatea traseului si o duce la cuib.
In zonele umede investigate am observat ca asa numitul ajutor sau doica prinde hrana de dimensiuni mari la distante destul de mari de mal, iar unul din cei doi parteneri o preia la mal si o duce la cuib.
In perioada de cuibarit, hrana este procurata atat din interiorul stufului cat si de la suprafata apei si din zonele de la marginea lacului, intrucat aici gasesc prazi de dimensiuni mai mari.
Puii la cuib
Comportamentul adultilor se schimba de o maniera radicala in aceasta perioada. Teritoriile nu mai sunt aparate si masculii inceteaza sa mai cante, daca nu au facut-o deja in perioada incubatiei.
Lacarul mare, este prin excelenta o specie altriciala in care puii se nasc slab dezvoltati si au nevoie de ingrijire parentala complexa si sustinuta. Grija parintilor fata de progenituri este in fapt o forma de altruism in care parintii ingrijesc mai intai puii si apoi pe ei insisi.
Cei doi parinti stau in primele zile la cuib si se inlocuiesc. Mai ales, pe perioada noptii isi acopera cu aripile puii. Comportamentul adultilor este mai discret. Ei realizeaza zboruri destul de joase, aproape de sol. Miscarile sunt scurte, rapide si mai putin increzatoare.
Rata de eclozare este, in cele mai multe cazuri observate de noi aproape de 100%. In 2003, 2004 am monitorizat 14 cuiburi, 2 de la Gardina Botanica (numai pe parcusrsul anului 2004) si 12 de la Iezar Botosani si am observat ca din toate ouale gasite de noi au iesit pui. In 2001, 2002, din 7 cuiburi tinute sub observatie am vazut ca din 21 de oua au iesit 15 pui. Cel mai adesea, insa nu toti puii supravietuiesc. La maturitate ajung unul sau doi pui, in mai mult de 50% din cazuri. Cei mai multi pui mor din cauza inanitiei. Fie ca cei doi parinti nu pot satisface nevoile de hrana ale puilor, fie ca resursele trofice in cazul unor furtuni si ploi repetate se imputineaza, asa cum a fost cazul in anii 2002, 2004., cand in perioada hranirii puilor au fost ploi de lunga durata.
Un alt motiv ce duce la o mortalitate ridicata a puilor este constituit de aparitia pradatorilor. Noi am observat ca serpii de apa (Natrix tesselata) sau cei de casa (Natrix natrix) urca pe firele de stuf si manaca fie ouale, fie puii abia eclozati.
De asmenea eretele de stuf (Circus aeruginosus) este unul din principalii pradatori ai puilor de lacar mare, avand in vedere si numarul lor relativ ridicat observat in zonele umede ale Moldovei (3 exemplare-10 ha).
Ulii, Accipiter gentilis si Accipiter nissus mai rar, actioneaza mai ales in stufariile rare, la inceputul sezonului de cuibarit (Hansson B., 2000).
Noi am observat atacuri ale uliilor in stuf la Beresti si Balta Lata. In acest caz lacarii incearca prin zboruri demonstrative, cu dese schimbari de directie, menite sa induca in eroare pradatorul, sa atraga pradatorii intr-o urmarire. Lacarul simuleaza in acest caz o pasare tanara, incapabila sa zboare, vulnerabila.
Puii sau ouale de lacar, din cuiburile instalate in interiorul stufariilor compacte pot fi mancate de Botaurus stellaris- buhaiul de balta si Rallus aquaticus- carstelul de balta. (Hansson B., 2000).
Noi am observat prezenta acestor specii in special in stufariile din nord, nord- estul Moldovei (Iezar, Balta Lata, Larga Jijia), si cele din sudul Moldovei (Carja si Vladesti). In aceste zone numarul cuiburilor de lacari a fost foarte ridicat de pana la 100 perechi clocitoare/10 ha (Ion C. ,2004), ceea ce presupune un risc al pradatorismului foarte ridicat.
In incursiunile noastre din teren am gasit si pui de lacar mare, cu capul mancat. In acest caz presupunem ca ei au fost ucisi de catre bizami, aflati intr-un numar mare in zonele umede cu mult stuf, de la Larga Jijia, Beresti, Carja sau Vladesti.
Un alt motiv al mortalitatii foarte ridicate a puilor il reprezinta intemperiile. Am observat la Iezar, Larga Jijia ori la lacul Gradinii Botanice ca dupa furtuni, in cazul anilor 2003, 2004 si mai putin 2003 oua cazute din cuib. De asemenea, am gasit cuiburi distruse, sau pui cazuti in apa.
Am constatat ca puiul care eclozeaza primul are cele mai multe sanse de supravietuire. El de obicei creste mai repede si are tendinta de a-si impinge fratii catre marginile cuibului. Culoarea care stimuleaza parintii sa ofere hrana puilor este reprezentata de rosul cavitatii bucale. Deschiderea larga a ciocurilor reprezinta un sociosemnal prin care puii sunt hraniti. In primele zile puii stau cu gurile deschise aproape tot timpul. Intredeschiderea gurii este insotita de semnale acustice constituite din piuituri intense. Daca la inceput masculul este cel care furnizeaza cea mai mare parte din hrana, pe masura ce puii inainteaza in varsta, miscarile pe care le realizeaza cei doi parinti sunt din ce in ce mai dese. Parintii isi hranesc progeniturile cu o frecventa care este adaptata in functie de varsta si talia puilor. In unele cazuri masculul aduce hrana, iar femela o preia si o plaseaza mai departe puilor sau si-o pastreaza pentru ea. Femela poate inghite hrana, dupa care la intervale de timp o regurguiteaza, semidigerata in gura puilor.
Adultii cand aduc hrana la cuib, nu se indreapta direct catre pui, ci mai intai executa zboruri planate pe deasupra cuibului si in apropierea acestuia, efectueaza salturi scurte si numai daca nu observa nici o schimbare, in mediul inconjurator, se indreapta la cuib.
La apropierea unui potential pericol, cum ar fi o pasare de prada sau o femela de cuc, adultul da drumul hranei din cioc si emite sunete de alarma. Puii devin tacuti si nemiscati. Dupa ce primejdia trece, se aud ambii parinti emitand sunete scurte, tremurate, de eliberare.
Puii au nevoie de o perioada scurta de timp pentru a invata sa zboare singuri si sa se hraneasca, aproximativ 2- 3 saptamani
In primele zile 2-3, conform si datelor din literatura (Tobler, 1924, citat de Cramp S., 1992), fecalele din cuib sunt inghitite de catre parinti. Mai tarziu, atat femela, cat si masculul cara excrementele in afara cuibului.
Penajul la pui creste foarte repede. La 2, 3 zile de la eclozare, deja corpul puilor este acoperit cu puf de culoare cenusie. La 5- 6 zile, dupa cum am constat puii au corpul acoperit cu pene de contur de culoare brunie. La peste 10 zile, apar si penele de zbor, remige si rectrice.
Daca penele de contur apar la toti puii, observati, la aceleasi intervale de timp, nu acelasi lucru putem spune despre penele de zbor. Ele apar in functie, mai ales credem noi, de hrana furnizata. La cuiburile observate pe timp frumos si la care parintii executau zboruri de hranire foarte dese, penele puilor aveau un ritm de crestere foarte rapid. La alte cuiburi la care puii au eclozat mai tarziu si care au fost suprinsi de ploi torentiale si vanturi cresterea lor si implicit a penajului a fost cu mult intarziata. Puii ce aparut mai tarziu aveau peanjul complet format abia dupa 30 de zile.
Emanciparea puilor
Puii devin zburatori dupa 10-21 de zile de la eclozare, conform observatiilor noastre.
La inceput, dupa ce parasesc cuibul, puii sunt inca incapabili sa zboare. La mai putin de o ora de la eclozare, ei se ascund in apropierea cuibului, Dupa circa 4- 5 zile pot sa-si ia zborul pe distante scurte. Dupa circa 10 zile de independenta paraseste teritoriul de cuibarit. De altfel, in momentul cand puii devin zburatori notiunea de teritoriu isi pierde sensul in cazul lacarului mare.
Puii isi semnalizeaza prezenta prin sunete scurte care se aud de la cativa metri.
Pe timpul cat isi castiga independenta ei invata sa-si caute hrana si sa o prinda. La inceput captureaza insecte sau alte nevertebrate de dimensiuni mici. Miscarile lor se limiteaza in spatiul stufului din jurul cuibului. Atunci cand executa salturi de pe o tulpina de stuf pe alta manifesta destula nesiguranta. Treptat castiga incredere si am observat ca-si urmeaza indeaproape parintii. Zboara mai intai in interiorul stufului compact la inaltimi destul de mici, dupa care isi incearca abilitatile pana in salciile sau tufisurile de la marginea apelor.
La lacul Gradinii Botanice am observat ca in perioada cand puii invata sa zboare si trebuie sa se departeze de cuib, unul din parinti sta de santinela pe unul din firele de stuf, emitand sunete scurte. Celalalt parinte cauta hrana in pajistile inconjuratoare, dupa care revine cu o insecta in cioc, da un sunet scurt, la care atat puii cat si partenerul ii raspund.
Parintii au grija de pui pana in momentul emanciparii acestora (moment, cand incep sa zboare). Acesta este si stadiul in care progeniturile devin independente. Cu toate acestea, noi am observat ca, sunt destul de rare cazuri in care cuibul, ponta si puii au parte de ingrijirea parintilor. Masculul paraseste cuibul intr-un stadiu anterior, cand la pui inca nu s-a format in intregime penajul de zbor.
Emanciparea completa a puilor are loc dupa 30-40 de zile de la eclozare. In anii in care abundenta de insecte in zonele umede investigate a fost mai mica, emanciparea puilor a durat si o luna si jumatate, astfel incat au devenit zburatori abia la inceputul lunii august.
Migratia de toamna
Cand puii devin buni zburatori, (la o varsta de aproximativ 30 de zile) ei pierd legatura cu parintii. Urmarind capturile, in plasele ornitologice (Ion C., 2004), care le-am avut la sfarsitul lunii iulie, inceputul lunii august, am observat ca se prindeau mai ales juvenili. Parintii parasesc mult mai repede teritoriile de cuibarit dupa ce si-au refacut rezervele energetice. Unii dintre pui vor ramane pana la sfarsitul lunii august, inceputul lui septembrie in teritoriile de cuibarit din Moldova, in care au eclozat. Indivizii ce apar la sfarsitul toamnei, septembrie, octombrie in zonele umede ale Moldovei sunt indivizi ce au eclozat in cartirele de cuibarit din nord, nord-estul Europei. Deplasarile la juvenili devin mult mai complexe, datorita dispersiei lor in habitatele cu stufaris si dezvoltarii sistemului de orientare dupa opinia ornitologului rus Mukhin A., 2003. Miscarile deosebit de active din timpul zilei, in opinia noastra sunt executate in principal pentru procurarea hranei, in perioada migratiei de toamna.
Adultii ierneaza la sud de Sahara in sud-estul Africii, evitant padurile tropicale (1997- The EBCC Atlas of European Breeding Birds). Primii lacari mari sunt deja semnalati in aceste cartiere de iernare la mijlocul lunii septembrie. Lacarii respecta principiul "leap- frog" (citat de Valcu M., 2003), conform caruia cu cat zona de cuibarit a unei populatii este mai in nord cu atat cartierul de iernat se afla mai in sud.
Juvenilii in aceasta perioada consuma o mare cantiate de hrana pentru a-si asigura energia necesara migratiei. Procesul de acumulare a energiei ca depozit lipidic, se produce treptat si este similar pentru toate cele trei specii de migratori de distanta lunga: Acrocephalus arundinaceus, A. scirpaceus, A. scheonobaenus.
Emanciparea puilor se termina, practic, in momentul formarii complete a remigelor si rectricelor. Plecarea juvenililor este decalata fata de cea adultilor cu 3- 4 saptamani. Din zonele umede ale Moldovei puii eclozati pleaca cel tarziu la sfarsitul lunii august.
In migratia de toamna, raportul numeric adulti/juvenili este puternic deplasat in favorea juvenililor, acest raport echilibrandu-se primavara din cauza mortalitatii mari a pasarilor aflate in primul an de viata.
In migratie aceste pasari nu fac popasuri decat in acele locuri in care gasesc resurse trofice suficiente pentru a-si satisface nevoile energetice. Am observat ca in perioada de toamna, pe timp nefavorabil, ploaie, vant, lacarii stau ascunsi. Au existat perioade de cateva zile, toamna, in care sa nu s-a observat nici un exemplar de lacar mare in zonele umede ale Moldovei. Lacarii mari pleaca cu putin inainte de declansarea intemperiilor, fie se adapostesc in interiorul stufariilor compacte. De altfel, am vazut ca inainte de inceperea ploilor, miscarile lor sunt foarte alerte si uneori chiar emit sunete de alarma. Daca vantul bate din nord ei nu se opresc din calatoria lor spre sud. Vantul reprezinta un factor ajutator pentru zborul lor catre sud (Schaub M., 2001). Cu toate acestea vanturile foarte puternice le pot frange aripile, iar ploaia din cauza dezvoltarii reduse a glandei uropigiene le poate afecta zborul, prin udarea penajului.
In noptile cu luna plina din luna Septembrie s-au observat grupuri mici de 5- 6 indivizi de lacari zburand pe deasupra stufariilor. Daca ploaia si vantul se opresc, lacarii mari isi continua zborul catre sud.
Cu toate acestea am observat ca in cazul unor conditii meteorologice nefavorabile, lacarii mari prezinta o toleranta mai mare la conditiile de mediu si la habitatele pe care le ocupa, fata de specii ca Acrocephalus scirpaceus- lacarul de stuf sau Acrocephalus schoenobaenus- lacarul mic.
In mai multe situatii am observat ca lacarii mari, in perioada august- septembrie, "fugarindu-se" prin stuf. Aceasta urmarire intre indivizii de lacar mare reprezinta un lucru mai putin obisnuit dupa terminarea perioadei de reproducere. Joaca lor este de fapt o incercare de apropiere intre indivizi pentru a forma stoluri mici, de 5 pana la 10 indivizi in vederea migratiei.
Comportamentul de hranire la Acrocephalus arundinaceus, Acrocephalus scirpaceus si Acrocephalus schoenobaenus.
Datele din literatura (Cramp S., 1992) arata ca lacarul mare- Arocephalus arundinaceus consuma artropode, in special insecte, putini paianjani, melci, mici vertebrate, uneori fructe si seminte, in afara sezonului de reproducere.
Din observatiile noastre reiese ca tehnicile de prindere a prazii la specia Acrocephalus arundinaceus sunt asemanatoare cu cele ale speciei Acrocephalus scirpaceus, dar miscarile de cautare a hranei sunt mai putin agile. Lacarul mare tinde sa se hraneasca mai aproape de sol chiar la baza vegetatiei. Prinderea insectelor in copaci sau arbusti, ca de exemplu in salcii- Salix sau stejari- Quercus este o practica obisnuita.
Distantele pe care le parcurge in stufarii, in cautarea hranei, in perioada cuibaritului sunt adesea destul de lungi, de cateva sute de metrii. Pe timp frumos, lacarul mare realizeaza incursiuni in terenurile agricole si pajistile inconjuratoare suprafetelor acvatice.
Stabilirea teritoriilor lacarilor este conditionata si de sursele de hrana disponibile. Hrana este mai mult sau mai putin distribuita uniform atat in interiorul stufariilor cat si in afara acestora, astfel incat lacarii ocupa teritorii exclusive conform pricipiului lui Torn. (Wilson E., 2003). Teritoriile sunt contigue si pozitionate astfel incat accesul spre pajistile din jurul lacurilor si spre apa sa fie mai usor. Cel mai adesea in interiorul teritoriului nu va cuibari nici o alta specie de pasare.
Lacarii mari au fost observati la elesteele Vladeni in luna Iunie, in timp ce carau la cuib insecte. Intr-o prima faza ei culeg insecte pentru nevoile proprii, de dimensiuni relativ mici. Ulterior obisnuiesc sa prinda mai ales insecte de talie mare, in special carabide, pe care le cara la cuib. In momentul deplasarii pe sol, lacarii se ajutau de aripi. Mersul lor este greoi si lovesc usor cu aripile solul incercand sa se propulseze inainte. Daca nu reuseste sa prinda insecta din prima miscare, atunci, executa un al doilea atac de pe un suport inalt, un fir de iarba, un tufis. Lacarul in mometul atacului decisiv, se arcuieste, isi intinde corpul, aripile sunt usor desfacute si aduse pe langa corp, gatul intins, capul pozitionat in linie dreapta, in prelungirea corpului, penajul neted. Lacarul mare, apoi se arunca asupra insectei strapungand-o cu ciocul.
De obicei cand vaneaza, lacarul mare, se dovedeste a fi deosebit de perseverent. Isi urmareste prada fie din iarba, fie de pe sol, dupa care o ataca in mai multe randuri pana o prinde. De multe ori vaneaza si nervetebrate aflate in zbor, dar niciodata nu se foloseste de picioare, ci doar de cioc. Cand urmareste din aer o insecta aflata la sol, are comportamentul asemanator cu al unei chire: aripile falfaie pe langa corp, coada se misca in plan vertical, mentinand echilibrul, iar capul si ciocul sunt indreptate in jos catre sol, intr-un unghi de 900.
In perioada de reproducere lacarii mari zboara in tandem pentru a aduce artropode la cuib. In perioada de migratie, perechile nu se mai mentin si fiecare partener isi cauta necesarul de hrana separat. Miscarile de cautare a hranei in cursul diminetii sunt repetate la intervale de 10- 15 minute, dupa care incepand cu amiaza deplasarile sunt din ce in ce mai rare. Hrana nu este consumata niciodata in afara stufariilor: Insectele sunt prinse, duse in preajma cuibului si faramitate. Prefera mai ales insectele cu o mobilitate ridicata asa cum sunt odonatele, mai ales din genul Orthretum sp. si Coenagrion sp..
De ingrijirea puilor si aducerea hranei la cuib, lucru valabil nu numai pentru lacarul mare ci si pentru speciile Acrocephalus scirpaceus si Acrocephalus scheonobaenus, noi am observat ca se ocupa pe langa perechea clocitoare, si unul, mai rar, doi indivizi cu rolul de doica. Ei ajuta la cresterea progeniturilor prin furnizarea unui supliment de hrana..
Situatia se schimba in migratie cand lacarii realizeaza o adevarata echipa pentru procurarea hranei, formand grupuri de 4- 5 indivizi.
Constatarile noastre vin sa le intregeasca pe cele ale altor specialisti (Chernetsov N., 1999) care afirma ca speciile de Acrocephalus cu cerinte ecologice distincte in perioada de reproducere, utilizeaza aceleasi habitate in timpul migratiei. Totusi ei nu intra intr-o competitie evidenta in perioada cand isi refac rezervele energetice.
Am constatat de asemenea, ca disponibilitatile de hrana favorizeaza speciile de lacari in zonele umede ale Moldovei (Ion C., 2004). Astfel, rezultatele filetarilor entomologice, pe parcursul anului 2004, au fost asociate cu cele ale capturilor si observatiilor ornitologice. In luna iulie (vezi tabel 9), in sezonul de reproducere al lacarilor, in zona umeda Iezar, unde au fost efectuate cercetari amanuntite, cele mai abundente artropode sunt dipterele, araneele si himenopterele. Prezenta acestor categorii de artropode este in stransa concordanta cu abundenta ridicata a speciilor Acrocephalus arundinaceus si Acrocephalus schoenobaenus si mai scazuta a speciei Acrocephalus scirpaceus. Odonatele, hrana preferata a lacarului mare nu se regaseste, din nefericire, intr-un numar direct proportional cu abundenta lor in filetarile entomologice, de aceea nu sunt bine reprezentate in interpretarile noastre.
Tabel 9: Abundenta si dominanta medie a artropodelor in luna iulie- lacul Iezar (Dorhoi)
|
No |
Taxon |
Abundance |
Dominance (%) |
|
Araneae | |||
|
Collembola | |||
|
Thysanoptera | |||
|
Heteroptera | |||
|
Homoptera | |||
Seria Auchenorrhyncha | |||
Seria Stenorrhyncha | |||
|
Hymenoptera | |||
Coleoptera | |||
|
Diptera | |||
|
Subordinul Nematocera | |||
|
Subordinul Brachycera | |||
TOTAL | |||
In migratie, in luna septembrie abundenta cea mai mare in plasele ornitologice instalate la Iezar a avut-o specia Acrocephalus schoenobaenus. In filetarile entomologice, realizate in aceasta etapa, cel mai mare numar de insecte apartine ordinelor Homoptera si Diptera (vezi Tabel 10) . Disponibilitatile de hrana sunt diferite fata de prima perioada, ceea ce a avut ca efect o repartitie distincta a exemplarelor speciilor de lacari in zona respectiva.
Tabel 10: Abundenta si dominata medie pe zi a artropodelor in luna septembire- lacul Iezar
|
No |
Taxon |
Abundenta |
Dominanta (%) |
|
Araneae | |||
|
Odonata | |||
|
Subordinul Zygoptera | |||
|
Subordinul Anisoptera | |||
|
Heteroptera | |||
|
Homoptera | |||
Seria Auchenorrhyncha | |||
Seria Stenorrhyncha | |||
|
Hymenoptera | |||
Coleoptera | |||
|
Neuroptera | |||
|
Diptera | |||
|
Subordinul Nematocera | |||
|
Subordinul Brachycera | |||
TOTAL | |||
Comparand perioada reproductiva cu cea de migratie s-a constat ca dupa reproducere, adultii de lacari se hranesc intensiv, ajuta la ingrijirea puilor si dupa ce juvenilii incep sa zboare, indivizii care au acumulat suficiente rezerve energetice zboara catre sud. Puii raman un timp pe loc, castiga in greutate iar la sfarsitul lunii august si in cursul lunii septembrie isi incep migratia. Astfel se explica si numarul mare de adulti vara si un numar ridicat de juvenili toamna.
Am constat ca in perioada de reproducere sau de migratie competitia interspecifica intre lacari este scazuta. In perioadele ce urmeaza momentelor cu o nebulozitate ridicata situatia se schimba, concurenta intre speciile de lacari si exemplare devine evidenta. In mai multe situatii am observat cum lacarul mare respinge prezenta lacarului de stuf sau a lacarului mic. Postura sa semana mai ales cu a unei pasari de prada: capul cobarat in jos, coada desfacuta usor in evantai si adusa inainte, cu aripile ridicate si aduse in fata. Uneori lacarul mare fugareste din teritoriul sau alte specii de paseriforme caracteristice stufariilor, lacari sau pitigoi de stuf, si chiar ii ciupeste de cap.
In cursul inelarilor realizate s-a observat ca cel mai mare numar de capturi in plasele ornitologice se inregistreaza dimineata intr-un interval de 2- 3 ore de la rasaritul soarelui. (vezi Fig. 57, 58) In aceasta perioada disponibilitatile de hrana sunt reprezentate de aranee, coleoptere, himenoptere si diptere. La lacul Iezar, cel mai mare numar de lacari pentru aceasta perioada apartine speciilor Acrocephalus scirpaceus si Acrocephalus schoenobaenus, regasite si in perioada de seara cu efective ridicate. Acrocephalus arundinaceus si Acrocephalus schoenobaenus utilizeaza cel mai frecvent resursele de hrana in primele ore ale zilei si seara. Lacarul de stuf, Acrocephalus scirpaceus, este o specie mult mai activa si isi cauta hrana pe parcursul intregii zile, cand comunitatile de artropode isi schimba componenta. La amiaza, cele mai abundente in filetari sunt homopterele, iar seara dipterele. Acrocephalus scirpaceus utilizeaza o hrana mai diversificata si mai bogata, intrucat necesarul de energie calorica la acesta este mult mai ridicat. Distantele pe care se deplaseaza si ruta de migratie este mult mai lunga comparativ cu cealalta specie cu care se aseamana in ceea ce priveste talia, greutatea si lungimea aripii.
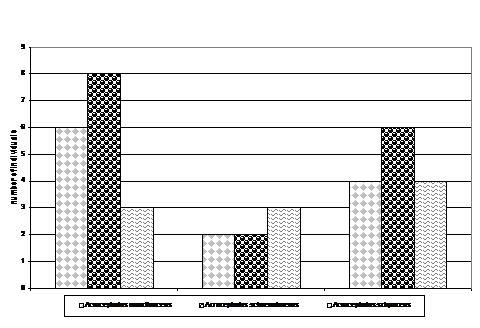
Fig. 57: Situatia medie a capturarii lacarilor in cursul unei zile
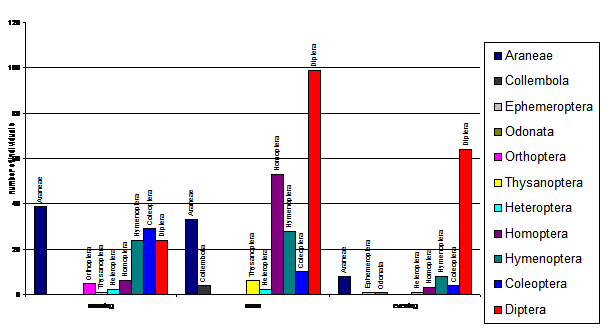
Fig. 58: Situatia numarului mediu de capturi a artropodelor pe parcursul unui ciclu diurn
Datele din literatura de specialitate (Chernetsov N., 1999, a, b, 2000) arata ca analiza continutului stomacal este in concordanta cu rezultatele noastre privind disponibilitatile de hrana.
Comportamentul de aparare
Lacarul isi apara teritoriul, cuibul si puii, prin miscari alerte, emitand triluri complexe de avertizare si amenintare. Comportamentul de aparare este evident mai ales in cazul delimitarii teritoriului si in momentul eclozarii puilor.
Agresiunea pe care o manifesta lacarul mare in cadrul comportamentului de aparare este de patru tipuri: 1) agresiunea teritoriala, in care foloseste cel mai expresiv comportament de semnalizare aflat la dispozitia sa pentru a respinge intrusii; 2) agresiunea sexuala, in care mai ales femela poate respinge la inceput masculul, pentru ca mai apoi sa il accepte; 3) agresiunea de dominare, intrucat intre ambele sexe se stabileste o relatie de parteneriat, in care fiecare incearca sa-si aduca intr-un fel sau altul aportul la succesul reproductiv si 4) agresiunea impotriva pradatorilor. Masculul de lacar poate executa manevre pur defensive care se pot transforma intr-un atac total asupra pradatorului, asa cum este cazul relatiei dintre lacar si cuc.
Spre exemplu, la auzul cantecului de pe un minicasetofon portabil instalat de noi, ocupantul teritoriului se urca pe firele de stuf si canta cu putere. Lacarul mare se apropie apoi de sursa sonora, face salturi in jurul acesteia, o priveste cu atentie, dupa care incearca sa o ciupeasca. Vazand ca prezumtivul intrus nu inceteaza cantecul se urca inapoi pe firele de stuf si isi etaleaza cantecul propriu. De data aceasta insa numai tine capul ridicat in sus, ci si-l indreapta, indiferent de pozitie in directia casetofonului portabil, de unde se aude cantecul.
Comportamentul lacarului mare la auzul unei inregistrari cu cantecul tipic speciei sale, este diferit de cel al lacarului de stuf, pus in aceeasi situatie. Lacarul mare manifesta foarte multa indrazneala si cerceteaza sursa sonora. Lacarul de stuf la auzul cantecului teritorial inregistrat, survoleaza putin terenul, dupa care paraseste zona respectiva.
Zburlirea penelor, ca si in opinia altor specialisti (Catchpole si colab., 1986, citat de Cramp S., 1992) reprezinta o reactie tipic agonistica (termen propus de Scott si Fredericson, 1951, citati de Wilson E., 2003). In momentul etalarii unei astfel de posturi, inseamna ca respectivul individ ia o pozitie de lupta, de aparare. Cel care se supune sau renunta la lupta, are penele netede si pare slab, dupa cum mentioneaza si Kluyver 1955, citat de Cramp S., 1992.
Unele mentuini din literatura (Springer, 1960, Catchpole si Leisler, 1986, Jackson si Stone, 1983, Springer, 1960, Tardeu, 1978, Panov, 1973, citati de Cramp S., 1992) sugereaza ca in cazul speciei Acrocephalus arundinaceus exista si cazuri de teritorialitate interspecifica.
Noi am observat ca in cazul speciilor Acrocephalus schoenobaenus si Acrocephalus arundinaceus nu poate fi vorba de asa ceva. Lacarul mic- A. schoenobaenus este o specie introvertita si mult mai ascunsa care-si cauta locuri de cuibarit departe de teritoriile lacarilor mari. Este capabila de zborui mai lungi pentru hranire si activitati comportamentale mai complexe. Uneori isi instaleaza cuiburile in interiorul stufariilor, la distante destul de mari de locurile de hranire, fie la marginea stufariilor dar in teritorii foarte restranse. De altfel lacarul mic- Acrocephalus schoenobaenus credem ca poseda o intelegenta mai mare decat celelalte specii de lacari. In momentul eliberarii din plase rareori se intorc catre acestea. In schimb lacarii mari sau lacarii de stuf sunt, in multe cazuri, recapturati in plasele ornitologice, la cateva minute dupa eliberare sau pe parcursul aceleeasi zile.
La speciile A. arundinaceus si A. scirpaceus am observat uneori lupte teritoriale, in care lacarul mare (A. arundinaceus) il ataca pe lacarul de stuf (A. scirpaceus), ciupindu-l de dupa cap, sau executa zboruri rapide in urma acestuia. In tot acest timp lacarul mare emite sunete monosilabice. Urmaritul, incearca sa evite atacul prin schimbari bruste de directie. De cele mai multe ori lacarul de stuf trebuie sa renunte la pretentiile teritoriale. Lacarii de stuf, cel mai adesea, formeaza grupuri familiale in teritorii neocupate de lacarul mare. Aceste zone sunt aparate in grup si au mai multe sanse de a fi protejate. Astfel am observat cate doi, trei lacari de stuf, (la Larga Jijia), urmarind un lacar mare si incercand sa-l alunge, din teritoriile lor.
In incursiunile noastre in teren am observat ca lacarul mare intra deseori intr-o competitie interspecifica cu chira cu obraz alb (Chlidonias hybridus). Aceasta isi construieste cuiburi plutitoare pe ochiuri de apa inconjurate de stuf. In stufariile ce marginesc locurile de cuibarit ale chirelor se instaleaza lacarii mari. Numai ca acestia nu pot zbura catre apa, ci sunt obligati sa zboare catre mal, sau in interiorul stufariilor. Chirele cu obraz alb daca observa un lacar mare in teritoriul detinut de ele imediat il ataca si incearca sa-l alunge.
A alta specie cu care stabileste relatii interspecifice lacarul mare, este privighetoarea de zavoi (Luscinia luscinia). In observatiile pe care le-am realizat la Vladesti- Oancea, (iunie, 2001) am constatat ca desi perechea era formata, la lacarul mare un mascul de privighetoare de zavoi a ocupat teritoriul, fara avea loc o lupta fizica, ci doar un duel al repertoriilor vocale intre masculi. Lacarii mari s-au vazut nevoiti sa abandoneze teritoriul lor. Dupa aproximativ 48 de ore, masculul de privighetoare de zavoi a parasit teritoriul, intrucat in respectivul spatiu nu a aparut nici o partenera. La mai putin de ora au fost vazuti lacarii mari intorcandu-se in teritoriul lor si cercetandu-l cu atentie. Totusi postura lor era mai degraba una de supunere: capul usor plecat, coada stransa, penajul neted, miscari agitate.
Langa cuiburile de lacari mari am gasit la Iezar cuiburi de starc pitic- Ixobrychus minutus cu care se pare ca convetuieste in interiorul stufariilor, intr-o aparenta comuniune.
La lacul Gradinii Botanice, am observat ca in teritoriul ocupat de lacari mari, daca apare un starc pitic cu o insecta in cioc, acestia se reped catre acesta sa-l indeparteze. Starcul atacat coboara in interiorul stufului. Lacarii dupa reusita actiunii lor, se ating usor cu piepturile, ca intr-un gest de multumire, dupa care femela zboara catre cuib, iar masculul se va aseza exact in locul in care a stat starcul in momentul patrunderii sale.
La indivizii tineri de lacar mare, chiar si la cei de lacar de stuf sau cei de lacar mic, nu am observat actiuni de competie interspecifica. Dimpotriva la juvenilii de lacar mare si cei de stuf, am observat ca in perioadele de toamna impart aceleasi locuri de hranire si de popas. Acelasi lucru se intampla si la lacarul mare si lacarul mic. Nu am observat in schimb apropieri intre lacarul mic si cel de stuf.
La Larga Jijia si Iezar s-a observat ca in habitatul ocupat de lacarii mari la distante nu foarte mari de acestia se afla pitigoii de stuf, atat adulti cat si pui, precum si presure de stuf. Intre aceste specii nu am constat actiuni de competitie interspecifica, acustice sau optice.
In zonele ce marginesc lacurile, cu pajisti, lacarii pot fi observati in compania codobaturilor: de munte (Motacilla cinerea), codobatura alba (Motacilla alba), codobatura galbena (Motacilla flava), care au regim de hrana tot insectivor. Cu toate acestea codobaturile nu intra in concurenta pentru hrana cu lacarii. Lacarii prind insectele zburatoare mai ales, pe cand codobaturile prind pe cele de pe sol.
Un comportament deosebit de violent il manifesta lacarul mare mai ales cand aude cantecul de cuc (Cuculus canorus) sau cand il observa pe acesta. Lacarul mare il poate chiar ataca sau urmari, emitand sunete de alarma. Cand observa apropierea cucului, cantecul lacarului mare devine tremurat, trancanit. Lacarul mare se apropie de marginea teritoriului, dupa care isi urmareste intrusul pana cand acesta iese din spatiul ocupat. Unele date din literatura (Molnar, 1944, citat de Cramp S., 1992) sugereaza, chiar ca intre lacarii mari si exemplarele de cuc pot exista lupte fizice, dovada fiind penele de cuc gasite in apa, in apropierea cuiburilor de lacar.
Daca lacarul mare are oua la cuib sau pui, la vederea cucului, el se retrage mai intai in stuf, dupa care revine pe varful firelor de stuf si isi etaleaza cantecul cu tarie, realizand o alternanta intre sunete tracanite de alarma si cantece teritoriale.
In cazul auzului cantecului de cuc, lacarul mare riposteaza cu "fraze scurte", dupa care se retrage spre cuib cercetandu-l. Pentru un timp nu se va auzi decat cantecul de cuc. Dupa aproximativ 5 minute lacarul mare isi reia pozitia in varful firelor de stuf, cantand si cercetand vizual zona.
Comportamentul lacarilor fata de om
Prezenta omului in teritoriile lacarilor mari trezeste multa curiozitate dar si teama. In mai multe situatii am asteptat zeci de minute, sau ore pentru a fotografia lacarii mari la cuib, deoarece acestia manifesta multa circumspectie la vederea omului. Femela executa, in prezenta noastra, salturi scurte pe tulpinile de stuf, la mica distanta de luciul de apa, dupa care urca in varful stufului, sau executa salt, dupa salt, in semicerc in jurul cuibului. Uneori realizau zboruri in spatele nostru intr-un alt petec de stuf, cercetandu-ne. (Fig. 63) Ulterior se reintorceau la locul unde era cuibul. Pasarea nu s-a apropiat mai mult de 20 cm de cuib, niciodata. La plecarea noastra ne-a urmarit cativa metri, dupa care s-a reintors la cuib. Masculul a fost mai circumspect, in sensul ca rar se apropia de noi sau de cuib. Zborurile sale erau ample, executate mai ales in treimea superioara a stufului emitand sunete de alarma care aveau darul sa ne alunge.
In cazul apropierii omului de cuib, niciodata lacarii nu se departeaza prea mult de cuib. Masculul este cel care da semnale de avertizare. Agitatia celor doi parteneri se accentueaza pe masura ce prezenta omului persista in teritoriului lor. Penele lacarilor de pe cap se zburlesc ca si cele de pe gat. Aripile sunt coborate usor, coada desfacuta in evantai luand o postura de lupta. Femela in tot acest timp emite si ea sunete, insa razlete si reduse ca tonalitate. (Fig. 58)

Fig. 58: Postura masculului la apropierea omului de cuib: penele de pe cap si gat sunt zburlite- foto original
In momentul capturarii unuia dintre parteneri cu ajutorul plaselor ornitologice la cuib, celalalt isi paraseste teritoriul si nu se mai aude cantand. In momentul eliberarii ambii parteneri emit sunete scurte. Lacarul necapturat emite sunete de chemare, iar cel eliberat sunete de eliberare.
Lacarii, la aparitia unui intrus de talie mare, asa cum este omul, recurg, in multe cazuri, la pozitii si atitudini de diversiune care au rolul de a induce in eroare. In nenumarate cazuri, la aparitia noastra in teritoriile de cuibarit ale lacarilor mari, unul din ei, de obicei, realizeaza zboruri intr-o alta directie decat se afla cuibul. Se opreste la distante apreciabile de cuib, 30-40 de m si isi etaleaza cantecul. Postura (fig.23) sa este una de atac ca si cum ar apara cuibul, cu penajul infoiat, aripile si coada desfacute. Alteori insa se lasa in josul stufului si scoate sunete tremurate. Corpul pare mult slabit. Penele sunt netezite. Salturile sunt foarte lente. Lacarul adopta o pozitie ca si cum ar fi ranit si se indreapta in zboruri nesigure intr-o alta directie decat cea in care se afla cuibul.
In literatura de specialitate (Ganvik, 1918, Dementiev si Gladkov, 1954, Dyrcz, 177, Francasso, 1978, Petretti, 1979, Beier, 1981 citati de Cramp S., 1992) se afirma ca lacarii mari pot ataca omul in cazul in care acesta patrunde in teritoriul lor. Noi am observat ca intensitatea atacului pe care-l pot exercita lacarii mari asupra omului se afla in corelatie cu stadiul in care se afla cuibaritul. Daca lacarii au oua, atunci, ei se vor departa foarte mult de cuib si vor urmari cu atentie actiunile noastre. Daca puii au doar cateva zile si corpul acoperit cu puf, femela va fi cea mai nelinistita si incearca sa se apropie de om si sa-l ciupeasca. Daca puii au penajul complet format atunci comportamentul parintilor devine extrem de violent fata de om. Ei ciupesc in special de cap, dar si de maini. Apropierea celor doi parinti de om se face de cele mai multe ori din doua directii. Am putea afirma ca lacarii aplica o stratagema in care te iau prin invaluire ca sa atace. Sunetele au o intensitate ridicata, de alarma, penele de pe cap si de pe gat sunt zburlite, coada desfacuta in evantai, remigele primare rasfirate ca la o pasare de prada.
Totusi cele mai multe intruziuni pe care le-am realizat in teritoriile lacarilor mari, la inceputul perioadei de cuibarit pentru a le cerceta cuiburile au fost urmate de plecarea acestora. Cu toate acestea in literatura (Granvik, 1918, citat de Cramp S., 1992) se mentioneaza ca femelele de lacar mare nu sunt foarte sperioase si daca i se fac vizite repetate va sta nemiscata. Aproape ca iti va permite sa o atingi.
In concluzie, etalarea comportamentului de agresiune devine evident la sfarsitul perioadei de reproducere, cand puii sunt mari si se apropie de momentul emanciparii.
Aspecte privind etalarea cantecului la Acrocephalus arundinaceus
Specialistii ornitologi apreciaza ca lacarii, masculii si femelele sunt monomomorfici si coloritul penajului este de un colorit mohorat, fara nici un fel de ornament (Cramp S., 1992). Singurele ornamente care atrag femela in vederea imperecherii (Catchpole, 1985, 1986 citat de Hasselquist D., 1998) sunt cantecul, in special si postura afisata de mascul.
Cantecul la lacarul mare este desoebit de complex, format din unitati (Ion C., 2003) ce se succed si alterneaza in diferite moduri, reprezentand una din cele mai importante caracteristici ale comportamentului sau. Cantecele la lacari sunt emise numai de masculi. Marea calitate a cantecului la lacarul mare consta in capacitatea acestuia de a modula volumul, tonalitatea, structura armonica si succesiunea unitatilor vocale, ceea ce creeaza un mare numar de semnale catre congeneri.
Hasselquist D., 1998, afirma ca marimea repertoriului vocal este corelata pozitiv cu succesul anual de imperechere al masculului. De asemenea, femelele aleg masculii cu un repertoriu vocal complex, intrucat s-a constatat ca masculii cu un astfel de repertoriu dau nastere la indivizi viabil genetic.
Noi am observat ca manifestarea acustica a lacarului mare este asemanatoare cu cea de la lacarul de stuf. De obicei, se agata pe un fir de stuf, penele sunt putin zburlite si canta. Intodeauna cand isi manifesta comportamnetul teritorial sau pe cel de reproducere masculul canta de pe o pozitie observabila, prin care isi face simtita prezenta.
In opinia unor specialisti marimea repertoriului vocal este corelata cu numarul de pui nou eclozati (Hasselquist D., 1998) dar si cu abilitatea de aprovizionare a puilor cu hrana (Greig- Smith, 1982, Hill, 1991 citati de Hasselquist D., 1998). Noi am observat ca sunt emise cantece cu intensitate proportional cu numarul de pui. Daca au pui multi atunci cantecele sunt mai scurte si formate din mai putine motive. Daca disponibilitatile de hrana sunt sarace atunci cantecele sunt ample si de lunga durata. In cazul unor resurse trofice bogate cantecele sunt inlocuite de zboruri de hranire dese.
Imediat dupa venirea din migratie, cantecele sunt sporadice. In schimb, dupa ce isi refac rezervele energetice cantecele contenesc pe parcusul intregii zile pana la perioada formarii cuplurilor. In timp ce canta adopta si o postura caracteristica: isi curata penajul cu ciocul; penajul este usor zburlit; coada este desfacuta in evantai; capul este indreptat spre interiorul lacului sau al stufariilor in care incearca sa-si stabileasca un teritoriu. Uneori fac mici pauze pentru a-si curata penajul capului sau al gatului cu ajutorul ghearelor de la unul din picioare. Exista si fluctuatii ale emisiei vocale ale lacarilor mari pe parcursul unei zile. Astfel in cazul unor temperaturi de peste 28 0C; 30 0C, taria sunetelor scade. De asemenea daca apar ploi torentiale, sau vanturi foarte puternice, cantecele scad in intensitate.
Cantecele sunt auzite, cel mai bine pe parcursul diminetii. Lacarii, cand incep sa cante in stufariile, formeaza un adevarat cor in zilele de primavara. Ecoul vegetatiei din jur contribuie la rasunetul emisiilor sonore. Diminetile foarte devreme cantecul lacarilor este bine auzit, intrucat in aceasta perioada din zi nu se hranesc. Temperaturile sunt scazute si este prea frig ca insectele sa zboare, si mult prea putina lumina pentru a le face usor observabile. In zonele umede studiate de noi, in zilele de primavara, dimineata, in general, nu bate, vantul, si astfel cantecul nu poate fi distorsionat.
La primele ore ale diminetii soarele tinde sa incalzeasca aerul de sus in jos. Din aceasta cauza aerul rece de la nivelul solului incepe sa se ridice, pentru a se egaliza presiunile intre cele doua paturi de aer. Undele sonore emise de lacari in loc sa se ridice sunt reflectate in jos de granita dintre cele doua straturi de aer. (Attenborough D., 1998).
Folosirea unui suport inalt cum este stuful pentru etalarea cantecelor teritoriale asigura in opinia unor specialisti (Jilka si Leisler, 1974, citat de Cramp S., 1992) o "penetrare sonora optima".
Lacarii emit nu numai cantece ci si sunete- semnal, care sunt scurte de obicei, simple si pot fi etalate atat de masculi cat si de femele. Semnalele pe care le produc lacarii sunt de obicei de aparare sau de avertizare a puilor, ori a partenerului.
La aparitia unui intrus cantecele devin sacadate, semn al faptului ca si-a observat concurentul. Dupa cateva secunde revine la un repertoriul complet si daca, nici in cazul unui duel vocal, intrusul nu paraseste teritoriu, stapanul pleaca in urmarirea lui.
In perioada cuibaritului, desi literatura de specialitate (Panov 1973, citat de Cramp S., 1992) mentioneaza ca lacarul mare canta in timpul zborului, noi nu am observat decat foarte rar acest lucru. Cand realizeaza zboruri in perioada reproducerii pentru a atrage femelele, emite sunete lungi ca durata, dar formate dintr-un numar mic de motive. Aceste semnale le-am incadrat, in categoria emisiilor sonore de chemare a partenerei. Aripile sunt usor coborate in zbor, cu varfurile putin ridicate, ceea ce ii permite sa planeze. Coada este tinuta dreapta si usor desfacuta, iar penele de pe gat sunt zburlite.
In zilele de primavara lacarii canta tot timpul cat lumina soarelui straluceste pana la aproximativ ora 21. Pe la 5- 6 dimineata ei isi reiau cantecul.
Perioada in care lacarii mari isi etaleaza cantecul nu se afla in corelatie cu activitatea cea mai intensa. Ei sunt activi in perioada de reproducere ca si lacarii mici in prima parte a zilei si la lasarea serii spre deosebire de lacarii de stuf care sunt mai activi la amiaza (Ion C., 2004). In migratie, atat lacarul mare, cat si cel mic sunt mai activi dimineata intre 6-14. Lacarul de stuf este mai activ dupa-masa (14-21). Datele noastre au fost confirmate partial si de alti ornitologi (Valcu M., 2003, Schaub si Jenni 2000).
La orele amiezii, cand este foarte cald, cantecele masculilor nu se mai aud de pe varful firelor de stuf, ci mai ales din interiorul stufariilor. Acelasi lucru se intampla si in cazul vanturilor foarte puternice (scara 4, 5 Beaufort) cand tulpinele de stuf se indoaie foarte mult. Noaptea exista anumite perioade de timp, cand nu se mai aud. Totusi acolo unde exista stufarii intinse, noaptea se aud razlet cantece de lacar mare. Spre dimineata spre ora 4, se aud din ce in ce mai multi masculi cantatori. Daca vremea este inourata lacarii mari continua sa cante. In cazul aparitiei unei ploi, insa catecul lor diminueaza foarte mult.
In perioada de stabilire a teritoriului, in cazul aparitiei unor intemperii, ei inceteaza sa mai cante sau se aud doar franturi de cantec.
Dupa ce are loc formarea cuplului, cantecele masculilor nu se mai aud cu aceeasi intensitate. Numarul masculilor ce isi etaleaza cantecul descreste. Noaptea nu se mai aud cantece, ci mai ales dimineata si seara.
In timpul construirii cuibului si apoi a incubatiei, sfarsitul lunii iunie, frecventa cantecelor descreste foarte mult. Interesant ca in aceasta perioada ploaia actioneaza ca un stimulent pentru lacarii mari in sensul ca inainte de venirea acesteia sunt foarte agitati, dovada fiind si un numar mare de capturi in plasele ornitologice. Dupa ploaie incepe sa cante destul de pregnant.
In cazul apropierii unui intrus de teritoriul lacarilor in perioada cand acestia aveau puii, ni s-a intamplat sa auzim de mai multe ori sunte asemanatoare cu cele ale unei pisici care toarce sau foarte asemanatoare cu sunetele de alarma ale unui grelusel (genul Locustella). Noi am presupus ca suntele emise de lacarul mare in acest caz fac parte din categoria sunetelor de amenitare.
Capacitatea de imitare a cantecelor altor specii este deosebita. Noi am observat spre exemplu, ca la apropierea unei femele de cuc de teritoriul ocupat, ambii parteneri scot sunete foarte ascutite, cu tonalitate inalta, asemanatoare cu ale unui sorecar sau acvile. Literatura bibliografica mentioneaza capacitatea de imitatie deosebita a lacarilor (Brehm, 1964). Ei pot etala cantecul a altor 19 specii de pasari.
De altfel, prezenta unui intrus in teritoriul de cuibarit al lacarilor mari in aceasta perioada functioneaza ca un factor declansator al cantecului. Ei incep sa emita sunete scurte.
Repertoriii complete la inceputul lunii iulie se aud doar dimineta intre orele 5 si 9.
In a doua jumatate a lunii iulie si la sfarsitul acesteia se mai aud doar indivizii care nu si-au gasit pereche. Exista si alta categorie de lacari care in experimentul nostru s-a apropiat de cantecul emis de pe un minicasetofon. Poate fi vorba de femele la care reproducerea a esuat sau pur si simplu de indivizi curiosi.
Lacarii mari sunt mult mai tacuti spre sfarsitul perioadei de cuibarit si dupa ce aceasta se definitiveaza, decat lacarii de stuf. Lacarii de stuf emit cantece ori sunete slabe pana la sfarsitul lunii august
Cel mai tarziu am auzit un lacar mare cantand in zona umeda Vladeni, pe 05.10.2002. Era vorba de un individ singuratic. El probabil era in cautarea unor companioni cu care sa porneasca in migratie.
Concluzii
Studiul nostru asupra ornitofaunei de stuf din unele zone umede ale Moldovei a relevat prezenta a 9 specii de paseriforme apartinand la 4 familii sistematice.
Zonele umede cercetate sunt: Balta Lata si Iezar (judetul Botosani), Larga Jijia (judetul Iasi), Carja (judetul Vaslui), Vladesti (judetul Galati), Beresti (judetul Bacau). Observatii detaliate au fost efectuate si in Delta Dunarii, in zona lacului Furtuna (Maliuc), ceea ce ne-a permis compararea populatiilor cuibaritoare din Moldova cu cele din delta.
Habiatele investigate ale passeriformelor de stuf sunt constituite din luciu de apa invadat partial de stuf, inconjurat de pajisti, culturi agricole, tufisuri.
Speciile de paseriforme de stuf au o distributie aglomerata in habitatele analizate, fapt determinat de restrangerea suprafetelor acvatice, datorita secetei din ultimii ani si micsorarea suprafetelor cordoanelor de stuf, datorita taierii stufului in vederea folosirii sale ca hrana pentru pestii fitofagi din iazurile piscicole.
Dinamica sezoniera a paseriformelor de stuf in zonele umede din Moldova inregistreaza variatii numerice ample. In aspectul prevernal, corespunzator migratiei de primavara, se inregistreaza un maxim, pentru ca apoi numarul lor sa scada in celelalte aspecte ecologice.
Analiza sinecologica a paserifomelor acvatice a relevat urmatoarele aspecte:
Acrocephalus arundinaceus si Acrocephalus scirpaceus prezinta efectivele cele mai mari;
Din punct de vedere al dominantei Acrocephalus arundinaceus si Acrocephalus scirpceus sunt considerate specii eudominate in zonele umede ale Moldovei, Acrocephalus schoenobaenus este specie dominanta,
Din punct de vedere al indecelui de constanta toate speciile analizate
Din analiza indicelui de semnificatie ecologica reiese ca rolul cel mai important in biocenoza paseriformelor de stuf este indeplinit de lacarul mare- Acrocephalus arundinaceus;
Cea mai mare afinitate cenotica este inregistrata intre speciile: Acrocephalus scirpaceus si Emberiza schoeniclus;
- Limitele intervalului de confidenta a variabilelor metrice si ponderale, pentru parametrii calculati de noi, se incadreaza in cele oferite de literatura de specialitate, cu mentiunea ca limita inferioara este, cel mai adesea, mai mica.
- In zonele investigate se pot observa unele diferentieri ale populatiilor pe baza anumitor caractere metrice, dar fara a putea afirma ca in cele patru zone se afla patru populatii locale distincte morfologic, cu exceptia speciei Acrocephalus arundinaceus- lacarul mare.
Cantecele speciilor Acrocephalus arundinaceus si Acrocephalus scirpaceus sunt asemanatoare intre ele si se diferentiaza de cel al speciei Acrocephalus schoenobaenus.
Cantecul la Acrocephalus arundinaceus are sase unitati distincte;
Cantecului la Acrocephalus scirpaceus in timpul perioadei de reproducere a evidentiat cinci tipuri de unitati.
La Acrocephalus schoenobaenus cantecul are sapte tipuri de unitati distincte.
Studiul caracteristicilor sunetelor permite identificarea mai facila a speciilor de lacari genul Acrocephalus, a reportoriiilor diferite a congenerilor aceleasi specii, si va permite elucidarea unor aspecte acustice comportamentale in sezonul de reproducere.
Comportamentul lacarilor este deosebit de complex, bazat pe indrazneala si retinere.
Lacarii fac parte din categoria migratorilor de distanta lunga, care vin tarziu in cartierele de cuibarit, la inceputul sau jumatatea lunii aprilie.
In migratia de primavara lacarii se aduna in grupuri de 4- 12 indivizi in cordoanele de stuf de pe lacuri, balti, zone umede. Ei efectueaza zboruri dese la marginea stufului pentru cautarea hranei. Comportarea lor este retinuta. Trilurile lor sunt scurte si de o intensitate scazuta.
La aproximativ 5-10 zile de la sosire, in aprilie, isi instaleaza teritoriul.
Teritorialitatea la lacari poate fi vazuta ca o forma de agresiune si competitie, cand intre congeneri exista o concurenta sexuala, dar si una pentru resurse de mediu, ce include posibilitatea de a se hrani mai usor si a gasi un adapost propice.
Spatiile ocupate de lacarii mari, in sezonul de cuibarit sunt contigue, ce variaza in medie intre 1000- 5000 m2, in medie. In cazul cordoanelor de stuf inguste, teritoriile sunt mai mici, intre 50- 100 m2.
Factorii ce favorizeaza teritorialitatea sunt: calitatea resurselor, distributia sa in spatiu si timp precum si competitia pentru resurse.
Primii care ocupa un teritoriu sunt lacarii mai in varsta. Indivizii tineri ajung mai tarziu si ocupa teritorii mai putin favorabile reproducerii si hranirii.
Lacarii ce ocupa un teritoriu manifesta multa agresiune fata de intrusi.
Suprematia aratata de un lacar sau altul, in perioada stabilirii teritoriilor, se rezuma la etalarea unor cantece complexe si a unei posturi intimidante.
Disponibilitatile de hrana sunt cauzele determinante in declansarea perioadei de reproducere.
Femele ajung in cartirele de vara cand deja teritoriile sunt formate.
Indivizii masculi urmariti de noi sunt monogami.
Parada sexuala, in cazul lacarului mare, se refera la afisarea prezentei de catre mascul prin cantec si postura.
Lacarul mare incepe constructia unui cuib, pe care femela il va cerceta, inainte de formarea cuplului. Definitivarea cuibului va fi continuata de ambii parteneri.
Masculul incearca sa atraga femela mai ales prin etalarea unor cantece "de chemare", cu triluri lungi, complexe cu amplitudine si intensitate crescute. Penele de contur ale masculului, in tot acest timp sunt infoiate, iar coada este desfacuta in evantai. Pentru a atrage femela in vederea acuplarii, masculul ii aduce, in cioc, femelei insecte.
Contactul ciocurilor masculului si a femelei reprezinta semnalul de formare definitiva a cuplului. Un alt semn al acceptarii masculului de catre femela il constituie preluarea firelor de material vegetal pentru cuib de catre femele din ciocul masculului.
In primele zile de formare a cuplului, cei doi parteneri manifesta foarte multa retinere, uneori agresivitate.
Perioada formarii cuplului dureaza, in opinia noastra, la lacarul mare 7-10 zile, si are loc in decursul lunii mai.
Cuiburile sunt construite din material vegetal impletit in jurul tulpinilor de stuf, iar constructia lui dureaza in medie 5 pana la 7 zile.
Depunera oualor are loc pe parcursul a 3-6 zile conform observatiilor noastre. O femela de lacar mare depune in medie intre 2-5 oua, in general un ou/zi
Incubatia oualor dureaza intre 10- 14 zile, la sfarsitul lunii mai sau in luna iunie. Femela este cea care cloceste. Rare sunt cazuri cand cloceste masculul.
Lacarii, in momentul cand au pui la cuib, nu mai canta iar notiunea de teritorialitate se pierde.
Rata eclozarii puilor este foarte mare, de aproape 100%.
Mortalitatea puilor depaseste 50 %. Cauzele mortii puilor sunt multiple: inanitia, pradatorismul, intemperiile.
Grija parintilor fata de pui, in fapt este o forma de altruism, in care parintii ingrijesc mai intai puii si apoi pe ei insisi.
Puii devin zburatori la 10-21 de zile de la eclozarea conform observatiilor noastre, cand li se formeaza penele de contur si cele de zbor.
Parintii au grija de pui pana in momentul emanciparii acestora
Emanciparea completa a puilor are loc la 30-40 de zile.
Lacarii adulti parasesc primii cartierele de cuibarit. Juvenilii din zonele umede ale Moldovei isi incep calatoria catre sud, din cartierele de vara, la sfarsitul lunii august, inceputul lunii septembrie.
In migratie lacarii se aduna in grupuri de 5, 6 indivizi.
Speciile A. arundinaceus, A. scirpaceus, A. scheonobaenus, cu cerinte ecologice distintcte folosesc acelasi habitate pentru hrana.
In timpul perioadei de reproducere, concurenta pentru resursele trofice apare doar intre congenerii aceleasi specii de lacar.
In timpul migratiei, concurenta intre speciile de lacari si intre congeneri este moderata.
Dinamica sezoniera si diurna a artropodelor din zonele umede are un rol foarte important in ceea ce priveste hrana lacarilor si declansarea competitiei pentru resursele existente.
Conditiile de mediu au un rol deosebit in comportamentul de hranire a lacarilor. Lacarii manifesta o activitate intensa de hranire dupa ploi, vant sau cand exista temperaturi joase. La temperaturi ridicate zborurile de hranire sunt mai rare intrucat, abundenta artropodelor este foarte mare.
Agresiunea pe care o manifesta lacarul mare in cadrul comportamentului de aparare este de patru tipuri: agresiunea teritoriala, in care foloseste cel mai expresiv comportament de semnalizare aflat la dispozitia sa pentru a respinge intrusii; agresiunea sexuala, in care mai ales femela poate respinge la inceput masculul, pentru ca mai apoi sa il accepte; agresiunea de dominare, intrucat intre ambele sexe se stabileste o relatie de parteneriat, in care fiecare incearca sa-si aduca intr-un fel sau altul aportul la succesul reproductiv; agresiunea impotriva pradatorilor.
Prezenta omului in teritoriile lacarilor mari trezeste multa curiozitate dar si teama.
Etalarea comportamentului de agresiune pentru aparare fata de om devine evident la sfarsitul perioadei de reproducere.
Cantecul la lacarul mare este desoebit de complex, format din unitati (Ion C., 2003) ce se succed si alterneaza in diferite moduri, reprezentand una din cele mai importante caracteristici ale comportamentului. Cantecele la lacari sunt emise numai de masculi. Marea calitate a cantecului la lacarul mare consta in capacitatea acestuia de a modula volumul, tonalitatea, structura armonica si succesiunea unitatilor vocale, ceea ce creeaza un mare numar de semnale catre congeneri.
Cantecele lacarilor sunt auzite, in perioada de reproducere, cel mai bine pe parcursul diminetii.
Comportamentul reproducator al pasarilor consideram ca trebuie studiat mai amanuntit, prin infiintarea unor puncte de observatii permanente. Prin reliefarea detaliilor legate de etologia pasarilor vom reusi sa intelegem mai bine lumea lor senzoriala.
Zonele umede ale Moldovei reprezinta sanctuare ale biodiversitatii florei si faunei din Romania si trebuie protejate pentru a se putea realiza o valorificare durabila a resurselor naturale existente.
Aceasta lucrare ofera o imagine globala asupra ornitofaunei stuficole din unele zone umede ale Moldovei si prefigureaza o serie de directii de cercetare ulterioare care sa contribuie la cunoasterea minutioasa a paserifomelor de stuf si sa fundamenteze o baza de cunostinte necesara pentru protectia si refacerea zonelor umede.
Bibliografie:
Copyright © 2024 - Toate drepturile rezervate