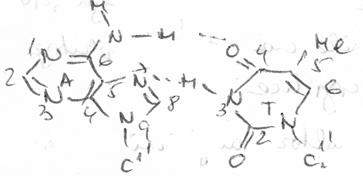|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Proprietatile fizice ale nucleozidelor si nucleotidelor
1. Solubilitate
Datorita caracterului (polar) ionic acizii nucleici sunt solubili in apa pana la o concentratie de 1% (m/v) depinzand de masa lor moleculara. Din solutiile apoase acizii sunt precipitati de alcooli (suspensii vascoase) de unde pot fi "colectati" prin agitare sau cu ajutorul unei baghete de sticla.
I.1.2. Ionizarea
Comportamentul acido-bazic al acizilor nucleici este
o caracteristica importanta deoarece determina incarcarea
(sarcina) pe diversi atomi, tautomerismul si deci, capacitatea de a
genera legaturi de hidrogen, legaturi ce determina
interactiunile dintre baze. Datorita existentei, incepand chiar
de la nivelul bazei azotate, ribozei sau restului de acid fosforic, a mai
multor centre apte de a accepta sau ceda protoni, acizii nucleici si
componentele constitutive ale acestora prezinta mai multe valori ale
exponentului de aciditate, pKa. pKa pentru diversele
centre de protonare variaza in functie de structura: nucleozid,
nucleotid, iar pentru acestea, de tipul particular molecular. Astfel (tabel),
in cazul nucleozidelor, valoarea cea mai scazuta (3 )
corespunde protonarii atomului N-7 al guanidinei, iar valoarea cea mai
ridicata (9,93) pentru atomul N-3 al timidinei. In cazul in care baza este
inglobata in nucleotid, valorile pKa cresc, de exemplu in cazul adeninei
se inregistreaza urmatoarele valori (mai mari pentru nucleozidele
Adenozina (N-1) 3,52
3'-Nucleotid 3,70
5'-Nucleotid 3,88
Din valorile pKa, rezulta ca la valorile fiziologice ale pH (5 < pH < 9) bazele aotate nu sunt ionizate.
In privinta pentozelor, ribozele pot ioniza la pH > 12, in timp ce, deoxiriboza ionizeaza la valori si mai mari (pH > 15), astfel ca la valorile fiziologice ale pH nici pentozele nu sunt ionizate.
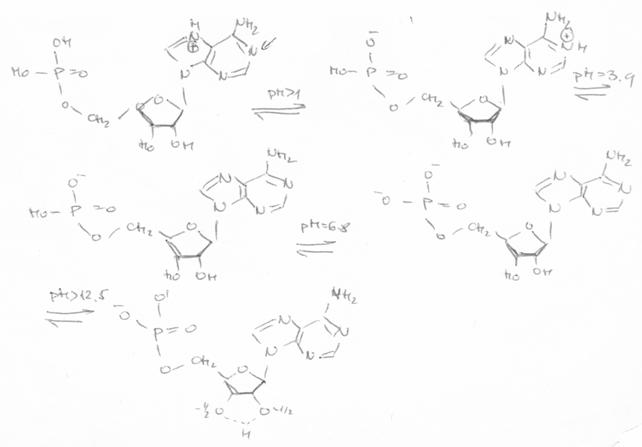
In sfarsit, in privinta gruparilor fosforice, acestea pierd un proton la pH > 1, iar cel de-al doilea proton este cedat la pH 7. Tinand cont de cele prezentate, se pot imagina urmatoarele structuri pentru AMP in functie de pH.
|
|
|
Structuri ionice ale acidului 5'-adenozin-monofosforic, functie de pH |
Dintre cele trei baze aminice, A, C si G, protonarea se realizeaza preferential la atomii de azot din ciclu si mai putin la gruparile -NH2 exociclice, deoarece legarea la aceasta grupare interfera cu procesul de delocalizare al electronilor neparticipanti; lungimea legaturilor C - NH2 este de aproximativ 1,34Å, ceea ce corespunde unui procent de aprox. 40% caracter de dubla legatura, in timp ce, in gruparile C=O din C, G, T si U au aproximativ 85 - 90% caracter de dubla legatura (v. tautomeria). Protonarea atomilor de azot din ciclu este favorizata si de pozitia gruparilor fosforice, care maresc bazicitatea atomilor de azot din ciclu; aceleasi grupari maresc, in schimb caracterul acid al gruparilor -NH2.
3. Tautomeria
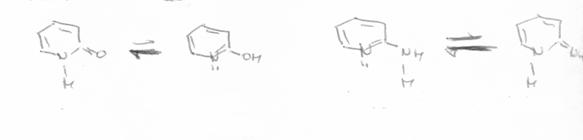
Tautomerii sunt izomeri care difera prin delocalizarea (pozitia) atomilor de hidrogen. Atat in cazul purinelor, cat si al pirimidinelor, sunt posibile structurile tautomerice de tipul:
ceto - enolica;
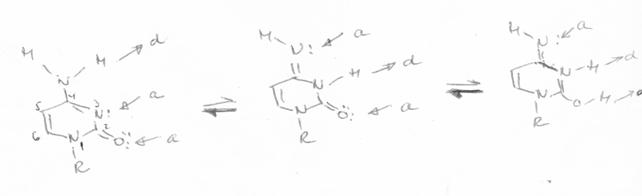
amino - iminica.
|
|
|
|
Tautomerie ceto - enolica (lactam - lactim) |
Tautomerie amino -iminica |
Toate determinarile spectroscopice (IR, UV, RMN) au demonstrat ca, la pH fiziologic, nucleotidele sunt, in proportie de > 99,99% in formele tautomere amino si ceto (lactam) motiv pentru care, in continuare ne vom referi la aceasta varianta structurala.
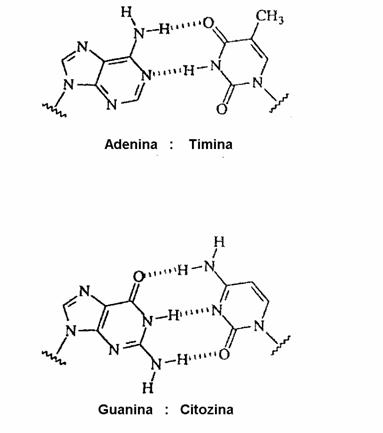
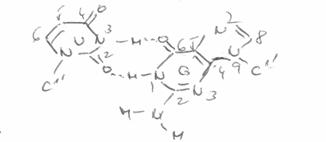
4. Legaturi de hidrogen
In modelul Watson Crick al ADN se presupune existenta unor legaturi de hidrogen intre bazele "complementare" A:T si respectiv G:C. Initial, considerandu-se ca bazele sunt in forma enolica, fiecare pereche realizeaza cate doua legaturi de hidrogen, ulterior, demonstrandu-se existenta formelor cetonice, pentru perechea G:C, s-a admis existenta a trei legaturi.
Gruparile - NH2 functioneaza ca donoare de atomi de hidrogen (d), in timp ce, gruparile cetonice (C = O), si atomii de azot din cicluri sunt mai puternic acceptoare(i) comparativ cu atomii de oxigen din gruparile fosfat sau hidroxil.
Legaturile donor - acceptor (d - a) au un caracter ionic pronuntat (0,2 eV) iar energia de legatura este de aprox. 8 - 12 kJ/mol.
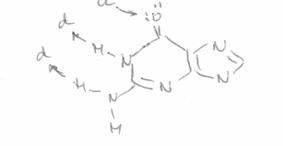
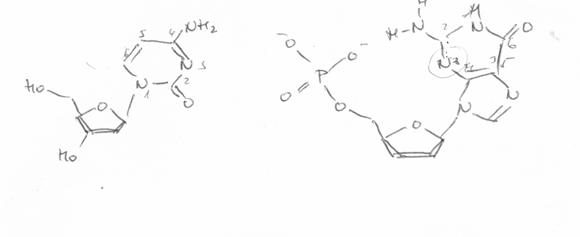
Citozina (in structura lactam) participa la formarea legaturilor de hidrogen prin atomii O2, N3, N4, fiind a.a.d in timp ce, izomerul minor (lactim) are o structura a.d.a, iar cel enol iminic o structura d.d.a. Structura formei tautomere a dG (N2N1O6) este d.d.a, fiind astfel total complementara cu citozina (lactam). Forma majora a dT este a.d.a, iar pentru dA este a.d, ceea ce permite formarea a numai doua legaturi cu A:T.
|
|
||
|
amino - ceto |
ceto - imina |
enol - imina |
|
Forme tautomere ale citidinei |
||
|
|
|
Structura "donor - acceptoare" a guaninei "normale": d.d.a |
Structura perechilor A : T si G : C a fost studiata in diverse molecule de acizi nucleici, ca si in oligonucleotide; determinarile s-au efectuat prin cristalografie cu raze X sau prin spectroscopie RMN.
Distanta dintre atomii de azot si oxigen, care
participa la formarea legaturilor de hidrogen este
de aprox. 2,80 - 2,95 Å, ceea ce face ca distanta dintre atomii de carbon
din pozitiile
Ca rezultat al acestei geometrii izomorfe, cele patru combinatii posibile A: T, T : A, C : G, G : C se pot forma cu aceeasi probabilitate intr-un duplex ADN.
|
|
|
Complementaritate. Legaturi de hidrogen |
Desi cele doua moduri de imperechere A:T, G:C (Watson - Crick) sunt cel mai frecvent intalnite in structura ADN, totusi, au fost propuse si alte variante (unele confirmate experimental) - perechile Hoogstean sau perechile "wobble" Crick.
Varianta Hoogstean nu este izomorfa cu perechile Watson - Crick deoarece unghiul dintre legaturile glicozidice este de 80 iar distanta dintre atomii de carbon anomerici este de 8,6 Å.
|
|
|
Pereche A : T "Hoogstean" |
Exista si variante, atat Watson - Crick, cat si Hoogstean "inversate", una dintre bazele din pereche fiind rotita cu 180 fata de modelul "normal"
Crick a propus si existenta unor baze perechi "wobble"
pentru a explica degenerarea codului genetic. Se
considera ca exista o baza in pozitia
|
|
|
Pereche U : G ("wobble"; non-Watson-Crick) |
Alte perechi analizate sunt I : A sau I : U.
Prezenta variantelor "non Watson Crick" este importanta deoarece este permisa formarea unor "triplexuri" de acizi nucleici. Prezenta legaturilor de hidrogen "anormale" a fost evidentiata prin difractie de raze X intr-o serie de oligodeoxiribonucleotide; se presupune ca aceste legaturi sunt prezente si in triplexuri de tip (poli (dA) 2 poli (dT) sau (poli (rG) 2 poli (rT) cel de-al doilea lant pirimidinic fiind situat la santul major al primelor doua catene, legandu-se de purina prin legaturi "Hoogstean".
1.1.2.3. Forma nucleotidelor
Nucleotidele au o forma destul de compacta, datorita existentai a numeroase interactiuni intre atomi; se pare ca nu exista diferente majore intre forma moleculei libere (monomer) si cea a moleculei incluse intr-un oligomer sau chiar o elice de acid nucleic De fapt, se presupune ca forma elicoidala a catenelor de ADN este determinata de conformatia moleculei libere. Forma unei molecule de nucleotitd poate fi definita prin lungimea legaturilor si prin unghiurile dintre acestea. Intr-o secventa oligonucleotidica se noteaza cu a b g d e,ζ, unghiurile dintre legaturile ce formeaza "coloana" fosfodiesterica, cu θ0 - θ4 unghiurile din ciclul pentozei si cu χ valoarea unghiului legaturii glicozidice.
|
|
|
Unghiuri de torsiune |
Datorita existentei multor unghiuri de torsiune, se incearca simplificarea prezentarii moleculei facandu-se referire la patru parametri:
plierea ("cutarea") zaharidului (sugar puchering) (pucher - a increti, a zbarci, a face cute);
conformatia sin - anti a legaturii glicozidice;
orientarea legaturii C4' C5';
forma legaturii fosfodiesterice.
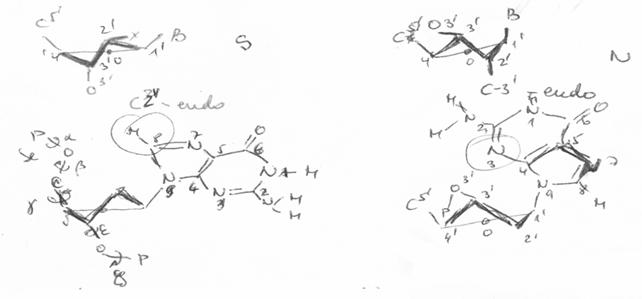
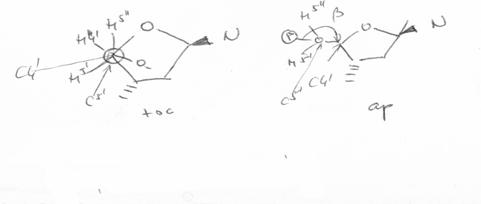
1.1.2.3.1. Plierea zaharidului
Ciclul furanozic nu este plan. El este incretit (twisted out) fata de acesta pentru a se minimaliza interactiunile dintre divertii atomi sau grupari ce nu fac parte din ciclu.
Plierea se poate caracteriza (defini) prin identificarea
pozitiei atomilor de carbon
|
|
|
|
C2'-endo / antiG (B-ADN) |
C3'-endo / sinG (Z-ADN) |
|
Plierea glucidului |
|
Se mai foloseste si o alta nomenclatura N (nord) si S (sud) corespunzatoare pozitiei atomului C3' fata de plan.
In solutie, intre cele doua forme S si N exista un echilibru, trecerea dintr-o forma in alta fiind facilitata de valoarea redusa a barierei energetice ( 20 kJ/mol). Echilibrul este influentat de o serie de factori:
preferinta substituentilor electronegativi din pozitiile C2' si C3' pentru orientari axiale;
orientarea bazei (pozitia sin favorizeaza pozitia C2'- endo);
formarea unei legaturi de hidrogen intracatenare intre atomul de hidrogen legat de O2' (dintr-o catena ARN) cu atomul de oxigen O4' din nucleotidul urmator favorizeaza plierea C3' endo.
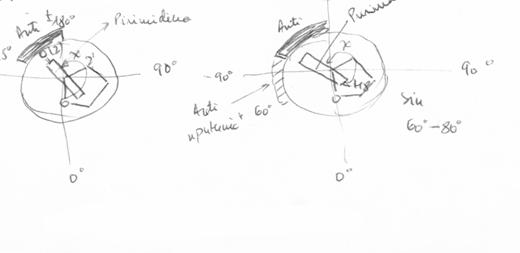
1.1.2.3.2. Conformatii sin - anti
Planul bazei azotate este aproape perpendicular pe cel al zaharidului, formand - aproximativ - bisectoarea unghiului O4' - C1' - C2', ceea ce permite bazei sa ocupe doua pozitii principale. Izomerul (conformerul) anti are atomul de hidrogen H6 (Py) sau H8 (Pu) deasupra ciclului glucidic, in timp ce izomerul (conformerul) sin are atomul (mai mare) O2 (Py) sau N3 (Pu) in aceasta pozitie. De obicei, pirimidinele contin un procent redus din conformatia anti, spre deosebire de purine, care contin domenii anti extinse (ex. formicina - tip de 8- azopurina).
|
|
|
Conformatii "sin - anti" |
Exemple:
|
|
|
|
anti-dC |
sin-dG |
Exista insa si exceptii, astfel prin
determinari
1.1.2.3.3. Orientarea legaturilor C4' - C5' / C3' - O3' (pozitia gruparii fosforice in mononucleotide)
Conformatia legaturii exociclice C4' - C5' determina pozitia gruparii fosforice 5' in raport cu ciclul zaharidului. Exista conformatii favorizate: sinclinal (sc), antiperiplanar (ap). In cazul nucleozidelor pirimidinele prefera varianta + sc, in timp ce purinele variantele + sc si ap cu concentratii aproximativ egale.
|
|
|
Orientarea legaturilor C4' - C5' / C3' - O3' |
In nucleotide, prezenta restului fosforic reduce gradele de libertate, astfel ca conformatia dominanta este + sc; cu exceptia Z-ADN care contine un procent ridicat de ap.
Conformatii similare se intalnesc si pentru legaturile C3'- O3'
Diesterii fosforici au o conformatie tetraedrica si antiperiplanara pentru legatura C5'- O5', in timp ce legatura C3' - O3' poate fi antiperiplanara sau anticlinala. S-a dezvoltat conceptul legaturii "virtuale" potrivit caruia lanturile P5' - O5' - C5' - C4 si P3' - O3' - C3' - C4' pot fi considerate ca determinand plane rigide, simplificare care permite calcule teoretice pentru structuri polimerice complexe.
Studii mai amanuntite, realizate cu raze X, asupra ARN
sau oligomeri ADN au demonstrat ca H4'- C4'- C5'- O
O situatie inversa se observa la legatura P - O5'.
Alte interactiuni de nelegatura impun ca ambele unghiuri a si z sa aiba valori apropiate de + 3000 (- 600) in structurile elicoidale, in timp ce in dinucleotide, valorile sunt de aproximativ + 600.
Modificari ale acestor valori se intalnesc atat in structurile poli nucleotidice ne-helicoidale si in cele de tip Z.
Copyright © 2024 - Toate drepturile rezervate