|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
HISTOLOGIE
Tesuturile: grupari de celule care au, in general, aceeasi origine, forma, structura si indeplinesc aceeasi functie. Tesuturile adevarate, ale caror celule sunt legate prin plasmodesme, caracterizeaza indeosebi plantele superioare.
I. 1. Tesuturi meristematice (meristeme)
II. 1.1. Meristeme primare (apicale)
In locul presupus a fi ocupat de initiale (conform teoriei clasice a histogenelor: dermatogen, periblem si plerom) se afla de fapt celule relativ vacuolizate, cu continut scazut in ARN si cu activitate de sinteza slaba, care se divid putin sau deloc, formand asa-numitul centru cuiescent (de repaus relativ).
Celulele cele mai active, care se divid intens, formeaza o zona de proliferare subapicala, care se intinde de la centrul cuiescent spre regiunea neteda, pe aproximativ 500-800 um lungime. In aceasta zona celulele se divid activ; ansamblul acestor celule formeaza un meristem primar; ciclul lor mitotic este rapid, derulandu-se in cateva ore (pana la doua cicluri pe zi).
La o anumita departare de apex, in partea inferioara a regiunii netede se pot distinge trei meristeme: protoderm (care va da rizoderma), procambiu (care va da tesuturile conducatoare) si meristem fundamental (care va da scoarta si maduva).
In jurul centrului cuiescent, sub apex se afla o zona de intretinere a scufiei, cu celule ce se divid periclin.
In lungul regiunii netede are loc un proces de diferentiere celulara, astfel incat cele trei meristeme primare (protoderm, procambiu, meristem fundamental) dau zonele anatomice, cu tesuturi definitive, ale radacinii in regiunea sa pilifera.
II. 1.1.2. Apexul caulinar. Studiile statistice, citochimice si experientele de microchirurgie au aratat ca zona apical-axiala (numita si meristem in asteptare) prezinta celule vacuolizate, putin active din punct de vedere mitotic in timpul vietii vegetative a plantei.
Teritoriul organogen propriu-zis se situeaza pe flancurile apexului, care prezinta celule foarte active, tipic meristematice, formand un inel initial subapical (numit si meristem de flanc); plecand de la acesta se formeaza frunzele si cea mai mare parte din structura tulpinii (Pl. X, fig. 2).

Structura tulpinii foliate
rezulta din activitate a unui meristem
subterminal, ca si cea a radacinii. Acest meristem
da o suita de frunze, ale caror baze formeaza tulpina.
Celulele superficiale ale inelului initial, detasate de el in timpul
cresterii fiecarui primordiu foliar, vor prolifera ulterior si
vor produce muguri axilari (ce stau
la originea ramurilor laterale) care, ca si frunzele, au origine
exogena (superficiala).
La baza zonei apical-axiale se afla meristemul medular, care nu participa la formarea frunzelor, ci va da maduva tulpinii, singura de origine propriu-zis caulinara; celelalte parti (epiderma, scoarta, fasciculele conducatoare) deriva din activitatea inelului initial, reprezentand baza foliara; fasciculele conducatoare se vor forma pe seama cordoanelor procambiale.
II. 1.2. Meristeme secundare (laterale). Dupa ce s-a terminat de format structura primara, pe seama meristemelor apicale, cresterea in grosime a multor plante poate fi continuata de meristemele secundare, ale caror celule se divid periclin. Meristemele secundare sunt cambiul si felogenul (Pl. X, fig. 3).
II. 1.2.1. Cambiul. In radacina, provine din procambiu si din dediferentiarea unor tesuturi definitive: periciclul (in dreptul fasciculelor de lemn) si parenchimul razelor medulare (intre fasciculele de lemn si cele de liber), ceea ce explica aspectul sinuos al cambiului initial, cand se trece la structura secundara. In tulpina, la nivelul fiecarui fascicul conducator, intre liber si lemn ramane un rest de procambiu, care va functiona ca un cambiu intrafascicular; pe seama razelor medulare primare, prin dediferentiere, se formeaza cambiul interfascicular. Cele doua feluri de cambiu se racordeaza, formand un inel (cilindru) meristematic continu, care va functiona bifacial si alternativ, dand spre exterior liber secundar si spre interior lemn secundar.
II. 1.2.2. Felogenul Pl. X, fig. 4). Ia nastere totdeauna dupa cambiu si printr-un proces de dediferentiere. De regula, in radacina se formeaza pe seama unor straturi corticale interne si pe seama periciclului, dand mai putin suber spre exterior si mai mult feloderm spre interior. Celulele de felogen se divid mai cu seama periclin si au forma unor prisme dreptungiulare (aplatizate radiar si alungite tangential) pe sectiuni transversale.
|
Care sunt caracterele celulelor meristematice? |
|
Comparati meristemele apicale si laterale dupa localizare, structura si mod de functionare. |
II.2. Tesuturi protectoare
Asigura protectia organelor vegetale impotriva agentilor externi daunatori din aer, apa si sol; in mica masura indeplinesc si rol de sustinere. Obisnuit sunt localizate la exteriorul plantei; exceptie fac endoderma si exoderma (la radacinile aeriene). Pot fi de origine primara (scufia, rizoderma, exoderma, endoderma, epiderma) sau de origine secundara (suberul).
a) Scufia (de cativa milimetri lungime): protejeaza varful radacinii in regiunea apexului meristematic. Are forma unui degetar, mai gros in portiunea axiala (numita columela), unde se afla aparatul statolitic. Celulele de la baza sunt tinere, cu pereti celulozici; cele de la varf si de la periferie sunt definitive, cu peretii usor suberificati, mor curand si se exfoliaza.
b) Rizoderma: este stratul pilifer al radacinii, avand rol atat protector cat si absorbant. Difera de epiderma prin absenta stomatelor si prin celulele sale cu pereti subtiri, necutinizati.
c) Exoderma (numita si cutis, suberoid, suber primar): protejeaza varful radacinii in regiunea aspra, unde rizoderma cu peri absorbanti s-a exfoliat. Cuprinde 1-4 straturi de celule fara meaturi intre ele, cu pereti usor ingrosati si suberificati, deci moarte la maturitate.
d) Endoderma: stratul cel mai intern al scoartei, fiind tipica la radacina, multe tulpini subterane si acvatice, protejand tesuturile vii ale cilindrului central.
Este formata din celule alungite, cu ingrosari de tip special, numite benzile (punctele) Caspary, ce alcatuiesc un cadru suberificat.
Aceasta structura particulara a endodermei se pastreaza la nivelul regiunii aspre, cu exceptia catorva dicotiledonate si a celor mai multe monocotiledonate; in acest caz, depozitele secundare de suberina (la dicotiledonate) si de lignina (mai ales la monocotiledonate) se depun pe peretii laterali si interni (ingrosare in forma de U, V, C, potcoava) si chiar pe toti peretii (in forma de O), exceptand celulele situate in dreptul polilor xilemici, care raman vii si se numesc celule de pasaj.
e) Epiderma (Pl. XI, fig. 1) protejeaza frunzele, piesele florale, semintele si tulpinile cu structura primara. La nivelul epidermei se pot afla stomate, peri (protectori si secretori), glande, spini, papile etc.
# Celulele epidermice propriu-zise sunt strans unite intre ele, variate ca forma, marime si dispozitie. Vazute din fata au contur poligonal sau neregulat, cu peretii laterali drepti sau ondulati la unele gimnosperme si la dicotiledonate, contur dreptunghiular si cu dispozitie in siruri paralele la monocotiledonate (la multe graminee, in acelasi sir alterneaza celule lungi cu celule sau cupluri de celule scurte. Peretele extern al celulelor epidermice este adesea bombat, mult mai gros, cutinizat, cerificat si, uneori, mineralizat (ca la graminee).
# Perii protectori (trihomii) sunt productii epidermice cu rol protector impotriva insolatiei, reflectand razele solare, impotriva pierderii de caldura noaptea, impotriva frigului si umezelii, impotriva animalelor.
Ca structura, prezinta o parte bazala (pedicel, bulb) aflata intre celulele epidermice si o parte terminala (corp), ascutita sau dilatata, dreapta sau arcuata la varf.
Tipuri de peri tectori (Pl. XI, fig. 2, 3, 4):
- unicelulari: papiliformi, aculeiformi sau foarte lungi; adesea ascutita la varf, cistolitica uneori (canepa - Cannabis sativa, urzica vie - Urtica dioica), protectori si secretori in acelasi timp (urzica vie - Urtica dioica), simpla (ranunculacee, umbelifere, rozacee) sau ramificati (in forma de suveica la micsunea - Cheiranthus cheiri sau de stea la traista ciobanului - Capsella bursa-pastoris);
- pluricelulari: simpli, uniseriati (solanacee, scrofulariacee, multe labiate, unele compozee), ramificati in forma de suveica, ac de busola (multe compozee) sau de stea (catina de rau - Berberis vulgaris, salce mirositoare - Eleagnus angustifolia), arborescent-etajati (lumanarica - Verbascum sp.) sau masivi (ferigi, mac - Papaver, maslin - Olea europaea)
|
Evidentiati corelatiile dintre structura si functiile tesuturilor protectoare |
f) Suberul (Pl. XII, fig. 1) (secundar) este format din celule tabulare, paralelipipedice, aplatizate, fara spatii intre ele si care mor repede dupa formarea lor, peretele suberificat impiedicand schimburile cu mediul extern sau cu celulele vecine, deoarece suberina (lipidica) este impermeabila. Aceste celule moarte sunt pline cu aer (uneori si cu tanin), ceea ce explica de ce suberul este atat de usor, fiind numit adesea pluta.
Suberul este prezent la radacini si tulpini in structura secundara (inclusiv rizomi si tuberculi), solzii mugurilor, unele fructe (par - Pyrus sativa); acestuia i se adauga suberul de cicatrizare ce apare la caderea frunzelor si fructelor.
Dupa cum stomatele permit schimburile gazoase la tulpinile tinere cu mediul inconjurator, lenticelele - cai de continuitate in grosimea suberului, joaca acelasi rol pentru tulpina in varsta sau pentru radacina.

Tesuturile situate la exteriorul suberului sunt, fiziologic, izolate de celelalte tesuturi vii si de aceea mor curand. Ele constituie ritidoma (coaja, in sens restrans), care se detaseaza periodic la nivelul unui strat de celule cu peretii putin suberificati sau ramane pe loc daca un asemenea strat nu s-a format; in acest caz, la nivelul ritidomei apar crapaturi, santuri prin care au loc schimburile de gaze.
|
Care sunt diferentele intre rizoderma si epiderma ? De ce este inlocuita epiderma cu suberul, in cazul edificarii structurii secundare ? |
II. 3. Tesuturi absorbante
Au ca rol absorbtia apei cu saruri minerale dizolvate in ea, din sol sau din aer si de a o transmite vaselor conducatoare de lemn, precum si absorbtia solutiilor de substante organice.
Sunt formate din celule vii, parenchimatice, mai rar prozenchimatice, cu pereti foarte subtiri, celulozici, localizate de regula la periferia organelor.
10 . Dintre celulele si tesuturile absorbante pentru apa si substante minerale men-tionam: rizoizii (muschi, protalul ferigilor, unele alge, ciuperci si licheni), hialoderma si hialocitele (muschi de turba - Sphagnum sp.), rizoderma cu peri absorbanti, velamenul, perii absorbanti de pe frunze si tulpini.
a) Velamenul (velamen radicum) este propriu radacinilor aeriene de la epifite tropicale (orhidee, aracee) tropicale carora le confera un aspect argintiu, celulele fiind pline cu aer.
.Celulele sunt izodiametrice sau alungite radiar, moarte la maturitate, fara spatii aerifere intre ele, cu peretii diferit ingrosati (spiralat sau reticulat) cu perforatii in portiunile ramase subtiri. Prin aceste perforatii este absorbita, prin capilaritate, apa de ploaie sau roua; atunci radacina ne apare verde, deoarece scoarta de sub velamen are celule cu cloroplaste. Datorita ingrosarilor peretilor, velamenul poate condensa vaporii de apa absorbiti.
Velamenul persista toata viata radacinii (la majoritatea epifitelor) sau este caduca la radacinile mai in varsta sau la cele ce ajung in sol.
b) Perii absorbanti de pe frunze si tulpini (Pl. XII, fig. 3) sunt caracteristici epifitelor ecuatoriale, plantelor stepice, desertice si mediteraneene.
Sunt, de obicei, solziformi si absorb picaturile de ploaie si de roua pe care le cedeaza tesuturilor inconjuratoare, indeosebi asimilatoare.
La Diplotaxis harra perii absorbanti sunt unicelulari, alcatuiti dintr-o parte bazala vie, umflata si din corpul parului alungit, cu peretii cutinizati si cu lumen umplut cu o masa celulozica stratificata (Pl. XIII, fig. 4).
c) Rizoderma cu peri absorbanti se afla la periferia regiunii pilifere a radacinii de la cele mai multe plante terestre si acvatice (cu radacina fixata in namol). Perii absorbanti lipsesc la plantele acvatice natante, la multi arbori cu micorize si la radacinile aeriene ale multor epifite tropicale.
Perii absorbanti sunt, in mod obisnuit, unicelulari, simpli, cilindrici, cu peretele subtire, format din doua paturi: una interna (calozica sau celulozica) si alta externa (pectica, mucilaginoasa). Perii absorbanti prezinta un sac citoplasmic parietal, nucleu subterminal si o vacuola centrala mare, plina cu suc concentrat. Numarul perilor absorbanti pe unitate de suprafata este de 200-400/mm2, plantele ierboase avand mai multi decat cele lemnoase.
20 Dintre tesuturile absorbante pentru solutii de substante organice mentionam: haustorii de la plantele semiparazite (vasc) si parazite (cuscuta, lupoaie, muma padurii), suspensorul embrionului, stratul absorbant al cotiledonului din samanta gramineelor s.a.
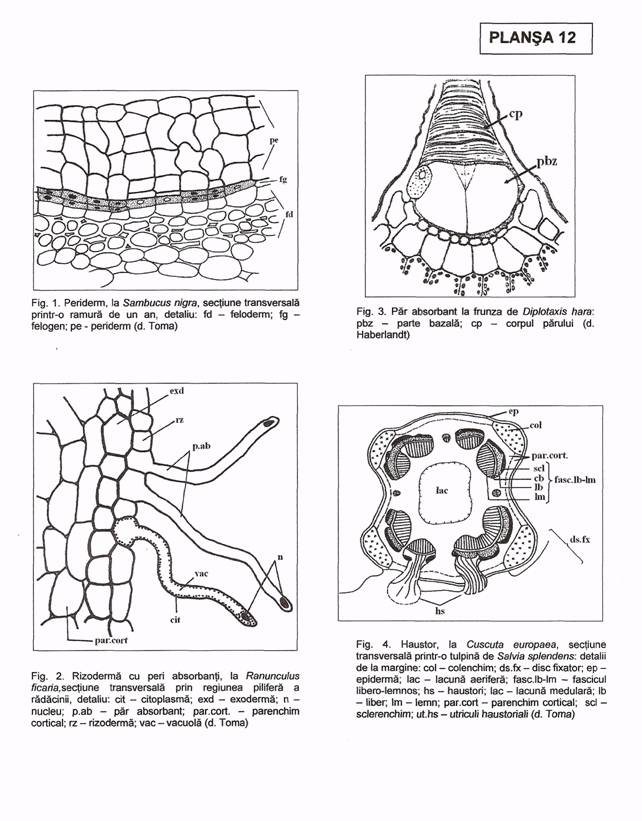
Haustorii (Pl. XII, fig. 4) sunt formatiuni absorbante ce iau din planta gazda seva bruta (la semiparazite) sau seva elaborata (la parazite). Ei se fixeaza pe partile aeriene (Viscum, Loranthus) sau pe parti subterane (Rhinanthus, Melampyrum) ale plantelor gazda.
 n La Viscum haustorul are forma de
pana ascutita, pluricelulare, ce patrunde in scoarta
arborelui, unde da ramuri tangentiale de ordinul I in zona liberului
secundar, iar acestea dau ramuri radiare in lemnul secundar, reprezentand
adevaratii haustori.
n La Viscum haustorul are forma de
pana ascutita, pluricelulare, ce patrunde in scoarta
arborelui, unde da ramuri tangentiale de ordinul I in zona liberului
secundar, iar acestea dau ramuri radiare in lemnul secundar, reprezentand
adevaratii haustori.
n La Cuscuta, haustorii se formeaza pe tulpina plantei gazda, luand nastere din periciclul plantei parazite sub forma unei mici prelungiri pluricelulari. Este alcatuit dintr-un disc fixator, inconjurand tulpina gazda si o prelungire sugatoare ce intra in scoarta si se ramifica in tesutul liberian, mai rar si in maduva. Aceasta prelungire are la varf celule lungi, vii umflate la polul apical, reprezintand adevaratii haustori.
|
Explicati corelatiile dintre structura si functiile tesuturilor absorbante |
II. 4. Tesuturi asimilatoare (clorenchimuri)
Celulele acestor tesuturi au cel mai important rol in metabolismul plantei; ele contin cloroplaste, indeplinind astfel functia de fotosinteza. Sunt prezente in toate organele aeriene, la periferia acestora, cu conditia sa fie iluminate, dar mai ales in frunze, organe specializate pentru functia de fotosinteza (Pl. XIII - fig. 1).
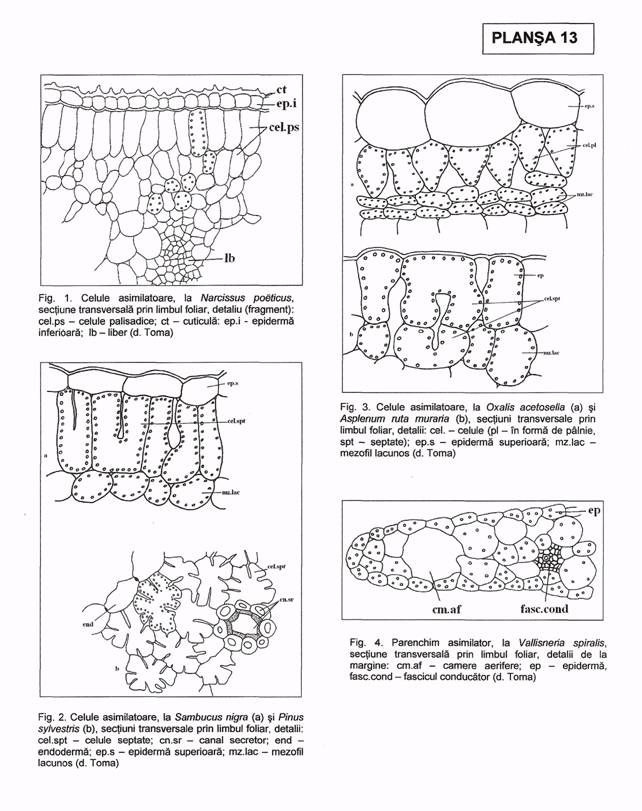
Celulele asimilatoare sunt vii, cu pereti subtiri, celulozici, cu spatii aerifere intre ele, cu cloroplaste adaptate pentru captarea cat mai eficienta a luminii.
Cele mai raspandite si mai adaptate celule pentru a indeplini functia de fotosinteza sunt cele palisadice, de forma cilindrica sau prismatica, perpendiculare pe epiderma, cu multe cloroplaste. Lungimea celulelor palisadice variaza de la specie la specie, in cadrul aceleiasi specii (in functie de internodul la care se insera frunza) ori acelasi organ (in lungul sau). Raportul lungime/latime al celulelor palisadice prezinta valori variind de la 1/1 (stir - Amaranthus retroflexus, papadie - Taraxacum officinalis, ricin - Ricinus communis) pana la 19/1 (paltin de munte - Acer pseudoplatanus).
Tesutul format din celule palisadice poate fi unistratificat sau pluristratificat, in functie de specie, dar si la aceeasi specie ori la acelasi organ, in functie de gradul de iluminare. In mod obisnuit tesutul palisadic se afla la fata superioara a limbului foliar (frunzele se numesc bifaciale heterofaciale sau dorsiventrale), mai rar la ambele fete (frunzele se numesc bifaciale izofaciale, ca la garoafa - Dianthus caryophyllus).
Dintre alte tipuri de celule asimilatoare, mentionam (Pl. XIII, fig. 2, 3): celule septate sau cu brate (pin - Pinus sp., unele ferigi, floarea pastelui - Anemone ranunculoides, soc s.a.), celule ale tecilor clorofiliene, dispuse radiar in jurul fasciculelor conducatoare (rogoz - Juncus sp., sfecla, porumb - Zea mays, grau - Triticum aestivum s.a.), celule asimilatoare izodiametrice (ca in frunzele, sepalele si bracteile multor graminee, liliacee, unele plante acvatice), celule ale parenchimului lacunos, izodiametrice pana la puternic ramificate, uneori alungite paralel cu epiderma si lasand lacune aerifere intre ele (ca in tesutul lacunos al frunzei si in scoarta tulpinilor ierboase).
|
Explicati corelatiile dintre structura si functiile tesuturilor asimilatoare. |
II. 5. Tesuturi si formatiuni aerifere
II. 5.1. Aerenchimul totalitatea spatiilor cu aer din organele plantelor. Spatiile cu aer pot fi (Pl. XIV, fig. 1): meaturi - canalicule foarte stramte, triunghiulare sau romboidale in sectiune transversala, mult mai mici decat celulele intre care se afla (in toate parenchimurile: cortical, liberian, lemnos, medular, palisadic); lacune - spatii izodiametrice, mai mari decat celulele intre care se afla (prezente in tesutul lacunos al frunzelor, in parenchimul cortical al multor radacini si tulpini); canale - spatii lungi si foarte largi, marginite de mai multe celule (prezente in organele axiale ale plantelor acvatice si palustre, putand fi continue sau intrerupte de diafragme); camere - spatii izodiametrice foarte largi (prezente in frunzele plantelor acvatice si palustre).
II. 5. 2. Formatiuni aerifere: stomatele si lenticelele
Stomatele sunt formatiuni alcatuite din doua celule stomatice reniforme, mai rar halteriforme (ca la graminee, ciperacee), ce lasa intre ele o deschidere numita ostiola, celule anexe, camera substomatica si celulele care o delimiteaza, camera suprastomatica (cand exista). Stomatele inlesnesc schimburile de gaze in procesul de respiratie, elimina apa sub forma de vapori (Pl. XIV, fig. 2) in procesul de transpiratie, fac fotosinteza (zaharurile simple ce rezulta avand rol in miscarea de inchidere si deschidere a stomatei).
La frunza, stomatele pot fi localizate: in epiderma superioara (frunze epistomatice: nuferi), in epiderma inferioara (frunze hipostomatice: caz obisnuit) sau in ambele epiderme (frunze amfistomatice: multe leguminoase, crucifere, labiate, cariofilacee, graminee). Numarul stomatelor pe unitate de suprafata prezinta valori medii cuprinse intre 30 si 300/mm2, fiind mai mic la plantele ierboase decat la cele lemnoase; stomatele sunt mai numeroase, dar mai mici, la plantele din locuri uscate si bine iluminate.
Celulele stomatice au citoplasma bogata, cu numeroase cloroplaste. Peretii sunt de mai multe feluri: extern (cutinizat), intern, dorsal (adesea mai subtire) si ventral (adesea mai gros si cutinizat), acesta din urma formand o creasta externa si una interna.
Celulele anexe, uneori diferite de cele epidermice obisnuite, sunt in numar de doua (graminee) sau patru (telegraf, Tradescantia virginica), rareori mai multe, localizate la nivelul celulelor stomatice sau mai sus (Agave, pin).
Dupa relatia dintre stomata si celulele anexe, la frunzele mature de la dicotiledonate s-au stabilit mai multe tipuri de aparat stomatic, dintre care mai raspandite sunt (Pl. XIV, fig. 4):
- tipul anizocitic (crucifer): stomata inconjurata de trei celule anexe inegale, dintre care una este foarte mica;
- tipul anomocitic (ranunculaceu): stomata inconjurata de un anumit numar de celule anexe, care nu difera ca forma si marime de celulele epidermice obisnuite;
- tipul diacitic (cariofilaceu): stomata inconjurata de doua celule anexe, al caror perete comun este perpendicular pe axa longitudinala a stomatei;
- tipul paracitic (rubiaceu): stomata inconjurata de una sau mai multe celule anexe, paralele cu axa longitudinala a stomatei.
La frunza monocotiledonatelor pot fi intalnite mai ales urmatoarele tipuri de aparat stomatic (Pl. XIV, fig. 5):
- aperigen (fara celule anexe de tip special, ca la majoritatea speciilor);
- biperigen (cu doua celule anexe, ca la graminee);
- tetraperigen (cu patru celule anexe, ca la comelinacee);
- hexaperigen (cu sase celule anexe).

II. 5.2.2. Lenticelele sunt formatiuni aerifere aflate pe tulpina si, uneori, pe radacina plantelor cu structura secundara, mai rar pe petiol, axa inflorescentei, fruct. Numarul lor variaza pe unitate de suprafata (cm2) de la 1-5 (salcie) pana la 30-50 (paltin de munte). Ne apar ca mici santuri inconjurate de un guleras de culoare bruna si au rol in schimburile de gaze dintre planta si mediul extern (Pl. XIV, fig. 6).
Tipuri structurale de lenticele (Pl. XV, fig. 1): tipul Sambucus (soc), cu lenticele formate exclusiv din tesut de umplutura (celule cu pereti subtiri dar suberificati); tipul Cerasus (cires) - Prunus (prun), cu lenticele formate din lame subtiri de celule suberoase, separate de tesut de umplutura (celule cu pereti subtiri, nesuberificati).
|
Care sunt tipurile de aparat stomatic dupa relatia dintre stomate si celulele anexe ? |
II.6. Tesuturi de depozitare
Sunt formate din celule vii, lipsite de cloroplaste, cu peretii subtiri sau ingrosati, mai adesea celulozici si cu multe substante de rezerva (Pl. XV, fig. 3); acestea se acumuleaza in citoplasma, amiloplaste, vacuole, mai rar in pereti.
Substantele depozitate pot fi: glucide (glucoza, fructoza, zaharoza, inulina, amidon, hemiceluloza, celuloza, mucilagii), lipide (uleiuri si unturi vegetale), protide (ca substanta fundamentala dizolvata in citoplasma sau ca granule de aleurona), alte substante, fara rol nutritiv (glicozizi, alcaloizi, taninuri, rasini, uleiuri eterice, gume).
Diferitele substante organice de rezerva se pot depune in celule ale parenchimurilor cortical (in radacini, tuberculi), liberian (in radacini tuberizate: morcov - Daucus carota, napi porcesti - Helianthus tuberosum), lemnos (in radacini: ridiche), medular (in radacini tuberizate, bulbi, rizomi, tuberculi, bulbo-tuberculi), in celulele maduvei radacinilor si tulpinilor normale, ale parenchimurilor din fructe si seminte.
Un tesut special de rezerva il constituie albumenul sau endospermul semintelor, in care predomina amidonul (ca la graminee), sau lipidele (ca la cocos, arborele de cacao).
Apa se acumuleaza in vacuolele celulelor din tesutul acvifer, bine dezvoltat indeosebi la xerofite si halofite, constituind o importanta rezerva pentru perioadele de seceta.
Dupa localizare, tesutul acvifer poate fi extern (in frunzele multor epifite tropicale) si intern (indeosebi la plantele suculente, dintre cactacee, liliacee, euforbiacee s.a.)
II. 7. Tesuturi mecanice (de sustinere)
Asigura soliditatea, elasticitatea si flexibilitatea unui organ vegetal, indeosebi aerian. Soliditatea, pozitia erecta a plantelor rezulta in primul rand de celulele turgescente, la care se adauga vasele de lemn. La plantele superioare au aparut, in plus, si tesuturi mecanice speciale, formate din celule numite stereide. Elementele mecanice sunt, adesea, celule lungi, cu peretii ingrosati, in cele mai multe cazuri grupate in cordoane sau fascicule. Dupa modul de ingrosare a peretilor, celulele mecanice apartin la doua tesuturi: colenchim si sclerenchim.
II. 7.1. Colenchimul este tesutul propriu organelor in crestere, indeosebi de la speciile de dicotiledonate, fiind deosebit de elastic si flexibil.
Celulele de colenchim sunt lungi (de contur poligonal in sectiune transversala), vii, avand pereti celulozo-pectici, adesea neuniform ingrosati. Originea colenchimului este mai adesea primara, provenind din meristemul fundamental si din procambiu. Obisnuit, colenchimul este localizat sub epiderma tulpinii, putand fi continuu ori discontinuu sub forma de cordoane (mai adesea in coaste). Celulele definitive sunt lungi pana la 2 mm si largi pana la 20 ìm, ascutite la varf, avand doar perete primar ingrosat si, uneori, cloroplaste. Dupa modul de ingrosare a peretilor celulari este cel putin de doua tipuri: angular si tangential .

a) Colenchimul angular: este mai adesea ameatic, celulele avand peretii terminali orizontali, cu ingrosari centripete numai la unghiuri; este prezent in tulpini, petiol si pedunculul florii ori al inflorescentei (de exemplu la labiate, cucurbitacee).
b) Colenchimul tangential (tabular): este mai adesea ameatic, celulele avand peretii tangentiali (extern si intern) mai ingrosati decat ceilalti; este prezent in tulpina si petiol (ca la multe plante lemnoase).
II. 7.2. Sclerenchimul este format din celule moarte la maturitate, ce contin mai putina apa, au pereti uniform si puternic ingrosati, total sau partial lignificati, nu lasa spatii aerifere intre ele si nu se mai pot alungi dupa completa lor diferentiere.
In stratul extern primar al peretelui microfibrilele de celuloza sunt dispuse longitudinal, iar in cel intern, transversal. In peretele secundar se disting trei straturi: S1 - microfibrile de celuloza dispuse in spirala, dupa un unghi de 45o fata de axa lunga a celulei; S2 (cel mai gros) - microfibrile dispuse tot in spirala, dar in sens opus; S3 - microfibrile orientate diferit.
Originea sclerenchimului poate fi: primara (provenind de regula din procambiu, ca la monocotiledonate si la dicotiledonatele cu structura primara) sau secundara (provenind din cambiu, mai rar din felogen, ca la dicotiledonatele cu structura secundara).
Ca localizare, sclerenchimul se afla atat in pozitie superficiala (sub epiderma), cat si in profunzimea organelor (in zona periciclica, in zona perifloemica, in liberul si in lemnul secundar, in maduva), putand fi continuu (inelar) sau discontinuu (sub forma de cordoane sau fascicule).
Celulele sclerenchimatice pot fi lungi (fibre), alcatuind sclerenchimul fibros, sau scurte (sclereide), alcatuind sclerenchimul scleros.
a) Fibrele de sclerenchim iau nastere din celule meristematice, se schiteaza de timpuriu, sunt alungite paralel cu axa organului si grupate in cordoane. Fibrele pot fi: lemnoase (libriforme si traheidale) si extralemnoase (corticale si liberiene, mai rar medulare).
Fibrele textile sunt fascicule de fibre sclerenchimatice (elementare) care in timpul 'topitului' se degaja de celelalte tesuturi. Lungimea fibrelor variaza mult: 0,5-0,8 mm (ca la arborii de chinina si de camfor), 2-3 mm (ca la tei), 4-40 mm (ca la in), 5-420 mm (ca la ramie); asadar, printre fibrele textile se afla cele mai lungi celule vegetale. Fibrele lemnoase de la dicotiledonate au origine secundara, sunt mai scurte, fusiforme, cu pereti mai ingrosati la esentele tari si mai subtiri la esentele moi. Se cunosc cateva categorii de fibre lemnoase, dintre care cele libriforme au cea mai larga raspandire la dicotiledonate.
b) Sclereidele (Pl. XVI, fig. 1) iau nastere din celule parenchimatice care se modifica secundar, au numeroase punctuatii si sunt localizate printre elementele tesuturilor fundamentale. Ele pot fi scurte, (pietroase sau brahisclereide), usor alungite (macrosclereide), cilindrice sau prismatice, cu capete umflate (osteosclereide) sau ramificate (asterosclereide), fiind localizate in toate zonele anatomice si organele vegetale.
|
Care sunt trasaturile structurale ale tesuturilor mecanice? |
II. 8. Tesuturi conducatoare
Asigura circulatia sevei in planta si sunt tipic reprezentate doar la cormofite, fiind alcatuite din elemente conducatoare prozenchimatice, parenchim longitudinal si fibre, acestea din urma fiind prezente mai cu seama in structura secundara a organelor vegetative axiale si apartinand, de fapt, tesuturilor mecanice. Originea tesuturilor conducatoare poate fi primara, provenind din procambiu sau secundara, provenind din cambiu.
II.8.1. Tesutul lemnos (lemn, hadrom, xilem). Asigura circulatia ascendenta a sevei brute (apa si saruri minerale dizolvate in ea) din sol, care a patruns in planta la nivelul stratului pilifer al radacinii.
Dupa ordinea de formare, lemnul primar se numeste: protoxilem (cu elemente diferentiate inainte de terminarea cresterii in lungime a organelor, avand peretii mai putin ingrosati si lignificati) si metaxilem (cu elemente incapabile de alungire ulterioara, avand peretii mai ingrosati si mai intens lignificati). Lemnul primar cuprinde elemente conducatoare (traheide, trahei) si celule de parenchim lemnos (longitudinal); in plus, lemnul secundar cuprinde si fibre lemnoase.
II. 8.1.1. Elemente conducatoare (Pl. XVI, fig. 2) (vasele) sunt lungi, moarte la maturitate, cu pereti putin sau moderat ingrosati si lignificati. Ele sunt reprezentate prin traheide si tarhei (sau vase in sens restrans).
Traheidele sunt elemente unicelulare, cilindrice sau prismatice, cu capete ascutite sau rotunjite, lungi de la cateva zeci de micrometri pana la 2-3 cm si largi de 1-10 micrometri, dispuse in siruri lungi. Prin ele seva circula mai greu, de aceea se mai numesc si vase imperfecte sau inchise. Peretii pot fi mai putin (la vasele inelate si spiralate) sau mai puternic (la vasele scalariforme si punctate) ingrosati. Sunt prezente la ferigi si gimnosperme, unele angiosperme primitive (de exemplu magnoliacee), la plantele parazite si la cele acvatice; la celelalte angiosperme, traheidele coexista cu traheile, indeosebi in lemnul secundar. Traheidele inelate si spiralate au cea mai mare parte din suprafata peretelui neingrosata si sunt primele elemente conducatoare adevarate aparute atat in filogeneza cat si in ontogeneza. Traheidele scalariforme sunt caracteristice mai ales ferigilor, avand barele scarii mai groase decat spatiile dintre ele si dispuse perpendicular pe benzile longitudinale. Traheidele punctate au cea mai mare parte din suprafata peretelui ingrosata. Au doua feluri de punctuatii areolate: cu torus (la gimnosperme - plante homoxile) si fara torus (la angiospermele dicotiledonate - plante heteroxile).
b) Traheile, numite inca si articule vasculare sau vase in sens restrans, sunt tuburi continue formate din suprapunerea de celule lungi si largi (articule de vas), intre care peretii transversali au disparut. Prin ele seva bruta circula mai usor, de aceea se mai numesc si vase perfecte sau deschise. Lungimea traheilor variaza de la cativa centimetri pana la 1,5 si chiar 3-5 metri (cele mai lungi intalnindu-se la liane), iar largimea, de la 10 la 70 micrometri.
|
Explicati corelatiile intre structura si functiile tesuturilor conducatoare |
II. 8.1.2. Parenchimul lemnos (longitudinal) este format din celule vii, putin alungite, cu punctuatii simple in peretii lor subtiri si celulozici (indeosebi in radacina) sau ingrosati si lignificati (indeosebi in tulpina). Au rol in depozitare, conducere, sustinere si formarea tilelor.
II. 8.1.3. Fibrele lemnoase sau libriforme sunt elemente lungi, moarte la maturitate, cu toti peretii puternic ingrosati si adesea lignificati, prezente in lemnul secundar de la dicotiledonate.
II. 8.2. Tesutul liberian (liber, leptom, floem). Asigura circulatia (in general descendenta) a sevei elaborate, bogata in substante organice sintetizate de catre parenchimul clorofilian.
Dupa ordinea de formare, liberul primar se numeste protofloem si metafloem, dar diferenta intre cele doua categorii de elemente liberiene nu este atat de clara ca la lemn. Liberul primar cuprinde elemente conducatoare (tuburi ciuruite, celule anexe) si, uneori, celule de parenchim liberian (longitudinal); liberul secundar cuprinde totdeauna, pe langa elemente conducatoare, celule de parenchim liberian si, uneori, fibre liberiene.
II. 8.1. Elemente conducatoare sunt lungi, vii, cu pereti adesea subtiri, totdeauna celulozici. Ele sunt reprezentate prin tuburi ciuruite si celule anexe (numai la angiosperme).
a) Tuburile ciuruite sunt formate din articule unicelulare prismatic-alungite, dispuse cap la cap, intre care se formeaza placi ciuruite (deci peretele comun pentru doua articule, orizontal sau oblic, perforat); rareori au campuri ciuruite in peretii laterali comuni la doua tuburi ciuruite.
Articulele tuburilor ciuruite sunt celule vii, de contur poligonal sau circular in sectiune transversala, cu o vacuola mare, citoplasma parietala, fara nucleu la maturitate.
Lungimea medie a tuburilor ciuruite (de la cateva zeci la cateva sute de micrometri) este mai mica decat a vaselor de lemn, rareori depasind 1 mm; largimea este de ordinul micrometrilor, rareori atingand 30 de micrometri.

Placile ciuruite sunt de doua tipuri: simple si multiple (Pl. XVI, fig.3, 4). Placile simple (tip Cucurbita), caracteristice in general plantelor ierboase, sunt acelea in care alterneaza portiuni din perete cu perforatii. Placile multiple (tip Vitis), caracteristice in general plantelor lemnoase, sunt acelea in care alterneaza portiuni din perete cu placi ciuruite simple.
Durata tuburilor ciuruite este scurta, de regula un sezon de vegetatie, dupa care porii placilor ciuruite sunt acoperiti de caloza. La conifere (unde sunt celule ciuruite, cu perforatii in peretii longitudinali), vita de vie, multe dicotiledonate cu frunze sempervirente, acelasi liber functioneaza doua perioade de vegetatie; mai rar (de exemplu la tei) acelasi liber functioneaza pana la 10 ani, intrand in repaus iarna; in acest caz placile de caloza din dreptul perforatiilor se dizolva primavara.
b) Celulele anexe, prezente numai la angiosperme, sunt mai scurte si mai stramte decat articulele tuburilor ciuruite, pastreaza citoplasma bogata in organite, indeosebi mitocondrii si ribozomi; nucleul este mare, iar vacuolele mici.
Se considera ca elementele ciuruite anucleate si celulele anexe nucleate formeaza un complex structural si functional unic, ce constituie unitatea fundamentala a tesutului liberian. Este cunoscut ca circulatia asimilatelor in floem necesita mari cheltuieli de energie. Fiind anucleate si cu organite partial dezorganizate la maturitate, elementele ciuruite nu pot indeplini singure procesul amintit, fara energia necesara; sursa acestei energii o constituie stransa legatura a elementelor cuiruite cu celulele anexe, acestea din urma pastrand multa vreme anumite trasaturi de celule meristematice, avand un aparat mitocondrial bine dezvoltat, care furnizeaza energia necesara pentru transportul sevei elaborate de catre elementele ciuruite.
II. 8.2.2. Parenchimul liberian (longitudinal) este format din celule vii, putin alungite, cu peretii celulozici, bogate in amidon, lipide si protide; lipseste, de regula, la monocotiledonate; are rol in depozitare si conducere.
II. 8.2.3. Fibrele liberiene au peretii foarte ingrosati, lignificati sau nelignificati. Rareori sunt prezente in liberul primar (de exemplu la in, canepa), mai adesea se afla in liberul secundar de la unele dicotiledonate provenind din cambiu; ele formeaza liberul tare: benzi longitudinale ce alterneaza cu liberul moale (tuburi ciuruite, celule anexe, celule de parenchim) (de exemplu la tei - Tilia sp.).
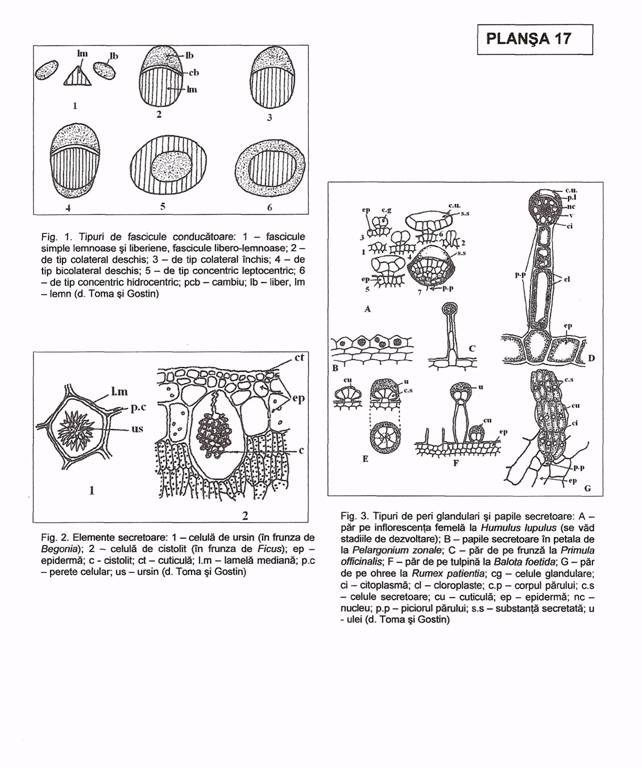
II. 8.3. Fascicule conducatoare In corpul plantelor, vasele conducatoare nu sunt dispersate la intamplare ci grupate in manunchiuri sau fascicule (Pl. XVII, fig. 1).
In radacina, fasciculele sunt simple, radiare, alternand unele de lemn cu altele de liber, toate avand o dezvoltare centripeta a elementelor componente.
In tulpina, frunze, piesele florale, fructe, seminte, fasciculele sunt mixte, libero-lemnoase, mai adesea de tip colateral, mai rar concentric.
Fasciculele de tip colateral. In tulpina, liberul este orientat spre exterior, iar lemnul spre maduva; in frunze, liberul este orientat spre fata inferioara, iar lemnul spre cea superioara; in ambele organe, lemnul are o dezvoltare centrifuga, iar liberul are o dezvoltare centripeta a elementelor componente.
La gimnosperme si dicotiledonate, fasciculele sunt de tip colateral-deschis, intre lemn si liber persistand un rest de procambiu (care se va transforma in cambiu atunci cand se trece la structura secundara); la unele dicotiledonate (dovleac - Cucurbita pepo, cartof - Solanum tuberosum) s.a., fasciculele sunt de tip bicolateral, lemnul fiind incadrat de doua zone de liber, intern si extern. La monocotiledonate, fasciculele sunt de tip colateral-inchis, intre liber si lemn nemaiexistand procambiu. In rizomii multor ferigi, fasciculele sunt de tip concentric-hadrocentric (lemnul fiind inconjurat de liber), iar in cei ai unor monocotiledonate sunt de tip concentric leptocentric (liberul fiind inconjurat de lemn).
II.9. Tesuturi (structuri) secretoare
Au ca functii principale elaborarea si acumularea substantelor straine metabolismului general, numite uneori deseuri sau produse de excretie. Substantele elaborate (uleiuri eterice, rasini, cleiuri, latexuri, taninuri s.a.) raman in celulele care le-au elaborat ori sunt eliminate in cavitati speciale din organele plantelor sau la periferia acestora. Unele din produsele secretate au roluri biolgice speciale: cleiurile - contra transpiratiei exagerate si a insectelor, alcaloizii - contra animalelor ierbivore, uleiurile eterice - pentru atragerea insectelor polenizatoare.
Celulele secretoare sunt vii, mici, cu citoplasma abundenta (foarte bogata in organite in perioada de secretie: REG, leucoplaste, dictiozomi s.a.), fara cloroplaste si cu peretii subtiri, adesea celulozici si saraci in plasmodesme.
Tesuturile sau structurile secretoare sunt reprezentate prin idioblaste sau celule izolate si complexe celulare.
II. 9.1. La idioblaste, produsele secretate se acumuleaza in vacuola centrala sau in spatiul dintre plasmalema si perete (de exemplu celulele cristalifere, celule ce secreta uleiuri, mucilagii, taninuri s.a.).
II. 9.2. Complexele celulare sunt localizate la exterior, pe suprafata organelor (papile, glande, peri) sau la interiorul organelor (buzunare, canale, laticifere).
a) Papilele secretoare sunt celule epidermice conice, care secreta uleiuri eterice si dau aspectul catifelat al petalelor; mai rar se afla pe tulpina, frunze, muguri (muscata - Pelargonium zonale, levantica - Lavandula officinalis, plop - Populus sp., castan porcesc); in cazul petalelor, toate celulele epidermice sunt papiliforme, formand un tesut secretor.
b) Glandele salifere sunt prezente la halofite si ne apar formate din 2, 4, 8 sau 9 celule care secreta saruri, prin gutatie activa; sarea se depune sub forma de eflorescente saline de calciu (Limonium, Tamarix, Atriplex, Saxifraga).
c) Perii secretori (glandulari) (Pl. XVII, fig. 3) sunt productii epidermice, mai adesea pluricelulare, ce secreta uleiuri volatile si sunt prezente mai ales pe frunze si tulpini, rareori pe bracteile inflorescentei femele de hamei sau pe catafilele mugurilor de la castanul porcesc.
In alcatuirea unui par secretor intra: baza sau tatana, adesea unicelulara si aflata intre celulele epidermice; pedicelul sau piciorul, unicelular sau pluricelular (uniseriat sau ramificat), rareori absent; glanda, capul sau partea secretoare, unicelulara ori pluricelulara, cu 2, 4 sau 8 celule dispuse in acelasi plan sau etajat, toate avand o cuticula comuna. Produsul secretat se acumuleaza intre peretele celulei secretoare si cuticula; aceasta se bombeaza si va plesni, eliberand astfel uleiul eteric in atmosfera.
d) Glandele secretoare sunt peri secretori de tip special, ce elimina enzime proteolitice, nectar, apa.
m Glandele digestive sunt caracteristice plantelor carnivore. Secreta mucilagii si enzime proteolitice, cu care sunt prinse si digerate unele animale mici (Pl. XVIII, fig. 1 A).
m Glandele nectarigene (Pl. XVIII, fig. 1 B) secreta nectar (solutie apoasa de zaharuri simple, uleiuri eterice, gume) si pot fi de doua feluri: florale (in diferite pozitii) si extraflorale (pe stipele, la baza petiolului si a limbului foliar).
m Hidatodele (Pl. XVIII, fig. 2) elimina apa lichida (cu saruri minerale) prin procesul de gutatie, la nivelul stomatelor acvifere ale frunzelor. Pot fi active (una sau cateva celule epidermice in forma de peri, care nu au legatura cu nervurile frunzelor) si pasive (doua celule stomatice moarte, permanent deschise, camera substomatica acvifera, parenchim incolor numit epitem, aflat in contact cu traheidele nervurilor).
e) Buzunarele sau pungile secretoare (Pl. XVIII, fig. 3 B) sunt spatii intercelulare izodiametrice, captusite de unul (la buzunarele schizogene) sau cateva (la buzunarele lisigene) straturi de celule epiteliale, secretoare de uleiuri eterice si rasini, balsamuri, gume, mucilagii). Sunt prezente in tulpina, frunza si fructul multor plante din familiile rutacee (citrice), mirtacee (eucalipt), hipericacee (sunatoare - Hypericum perforatum), cactacee, compozee s.a.

f) Canalele secretoare (Pl. XVIII fig. 3) sunt spatii intercelulare lungi, formate pe cale schizogena, inconjurate de un strat de celule epiteliale; la conifere sunt inconjurate si de un strat de celule mecanice. Sunt prezente in toate partile plantei, fiind intalnite la numeroase specii si numindu-se: oleifere (compozee, umbelifere), mucilagigene (malvacee, urticacee), rezinifere (conifere).
g) Laticiferele (Pl. XVIII fig. 3) sunt structuri secretoare plurinucleate, sisteme de celule cu peretii dintre ele disparuti sau celule plurinucleate care cresc nedefinit (laticifere nearticulate, simple - urticacee, canabacee sau ramificate - unele euforbiacee). In ambele tipuri, latexul se acumuleaza in unica vacuola centrala, mai rar in citoplasma. Laticiferele sunt prezente la peste 12.500 de specii, dintre care peste 1.800 au cauciuc in latex. Ca localizare, se afla in diferite organe si tesuturi, predominand in scoarta si liber. In laticiferele adevarate (nearticulate) latexul este vacuolar, iar in cele false (articulate) este citoplasmic.
|
Explicati corelatiile intre structura si functiile tesuturilor secretoare. |
Copyright © 2024 - Toate drepturile rezervate