|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
Informatii generale
Virusurile zoonotice sunt virusuri transmisibile de la animale (artropode, vertebrate) la om. Arbovirusuri (= arthropod borne) este o denumire colectiva pentru acele virusuri care (1) se replica in unele artropode (2) in unele vertebrate si care (3) sunt transmise de catre artropode intre gazdele lor vertebrate, odata cu saliva in timpul hranirii cu sange. Unele sunt transmise prin intermediul unui artropod vector hematofag infectat. Altele pot fi transmise prin inhalare, contactul conjunctivelor cu excretii infectate sau prin contactul direct cu animalele infectate.
Transmisia biologica (ciclica) cu replicarea virusului in artropod este criteriul de baza. Un virus ce adera la piesele bucale ale unui artropod si este transmis mecanic fara o replicatie prealabila in glandele salivare nu este un arbovirus. Arbovirusurile sunt definite exclusiv de caractere biologice si ecologice.
Sistemul taxonomic
Aproximativ 600 de virusuri au fost desemnate ca arbovirusuri. Ele se gasesc in diferitele familii virale: Reoviridae (circa 150 de specii), Bunyaviridae (circa 200 de specii), Flaviviridae (circa 50 de specii) si Togaviridae (circa 30 de specii). Tabelul 13 si Figura 3, prezinta familiile si genurile de virusuri ce cuprind si arbovirusuri.
Unele dintre virusuri numite "specii" reprezinta in fapt numai tulpinile usor diferite ale altor "specii", cu siguranta ca mai ramane un mare numar de arbovirusuri ce nu au fost detectate. Este dificil de estimat cate arbovirusuri pot exista - posibil cateva mii. De aceea arbovirusurile vor continua sa reprezinte o provocare importanta pentru virusologi si pentru parazitologi.
Multe dintre specii sunt slab cunoscute. Unele dintre ele produc stari febrile la om (si animale), desi pana in prezent (in conditii naturale) nu au fost corelate cu simptome de imbolnavire (ceea ce nu inseamna ca sunt apatogene).
Conceptul de specie, gen si familie in virologie este diferit de cel referitor la alte discipline biologice. Un sistem filogenetic real al virusurilor nu poate exista din mai multe motive.
Virusurile sunt diferite de organismele vii, apoi virusuri similare pot avea origini independente si diferite, desi evolutia lor depinde de factorii bine cunoscuti: mutatii, selectie, izolare etc.
Majoritatea familiilor virale si unele genuri nu sunt monofiletice desi reunesc membri cu un grad ridicat de similaritate (genomic) si in unele cazuri cu origine similara dar independenta. S-a sugerat ca cel putin in cazul arbovirusurilor, acestea deriva de la gazdele lor artropode. Aceasta supozitie este suportata de faptul ca arbovirusurile nu produc boli sau nu reprezinta o amenintare pentru gazdele lor artropode (exista totusi exceptii, printre Togaviridae) dar adesea prezinta pericole mai mult sau mai putin serioase pentru gazdele lor vertebrate.
Distributia celor mai importante familii - Arbovirusurile sunt raspandite pe toate continentele, chiar si la latitudini ridicate unde tantarii pot dezvolta populatii considerabile. In plus, pasarile care ajung in zonele nordice indepartate pot actiona ca gazde pentru virusurile ce au ca vectori capusele. Numarul arbovirusurilor endemice pentru o anumita regiune geografica sporeste catre ecuator, zonele tropicale si cele subtropicale gazduind cel mai mare numar de arbovirusuri.
|
FAMILIA |
INVELIS |
SIMETRIE |
GENOM |
IMAGINE |
MARIME |
|
Togaviridae |
da |
icosaedrica |
Un singur lant de ARN |
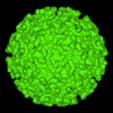
Alfavirus modelul suprafetei, generat de computer |
|
|
Bunyaviridae |
da |
helicala |
Un singur lant de ARN segmentat |
|
|
|
Reoviridae |
da |
icosaedrica |
catena dubla de ARN segmentat |
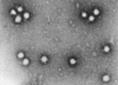
Reovirus de tipul 3, invelisul intern si extern al capsidei. |
|
In Europa au fost izolate aproximativ 30 de arbovirusuri, printre care si cele trei flebovirusuri transmise de catre flebotomi, ele cauzeaza febrele Papataci raspandite in sudul Europei. Desi, la nivel european virusul encefalitei de capusa ramane cel mai important arbovirus, alte cateva virusuri pot provoca imbolnaviri ale sistemului nervos central.
Europa Centrala gazduieste (in unele perioade si in unii ani) 11 arbovirusuri.
Sase dinte ele au ca vector capusele: Tribec, Lipovnik, Eyach, Uukuniemi, Bhanya si encefalita de capusa (TBE); toate sunt transmise de catre ixodide, in principal Ixodes ricinus.
Cinci virusuri sunt transmise de catre tantari : Calovo, Tahyna, Lednice, West Nile, Sindbis; Calovo este transmis de catre Anophelinae, celelalte de Culicinae.
Triburile, Lipovnik, Eyach, Uukuniemi, TBE, Calovo, Tahyna si (cel putin partial) Lednice sunt probabil endemice pentru Europa Centrala unde si hiberneaza. West Nile si Sindbis (partial virusul Lednice) sunt cel mai probabil introduse regulat de catre pasarile migratoare. De asemenea, virusurile Tahyna si Calovo sunt endemice (in sensul ca pot sa apara in zona in tot cursul anului).
Se pune intrebarea daca Europa Centrala gazduieste virusuri autohtone ce au ca vectori tantarii sub aspect strict biogeografic. Din cele 11 virusuri 9 pot produce boli la om, cel mai important dintre ele fiind virusul encefalitei de capusa responsabil de cazuri severe in fiecare an, asociat sporadic chiar cu decese. Dupa producerea unui vaccin si dupa aplicarea lui la scara mare la populatia umana incepand din anii 1970, importanta TBE in Europa Centrala a scazut mult.
Tabel 13. Familiile de virusuri ce contin arbovirusuri.
Familia |
Caracterizare |
Genurile reprezentative |
Principalele gazde Artropode |
|
"African swine fever-like viruses" (familie fara denumire) |
ADN dublu catenar, virusuri sferice, cu invelis 175-215 nm |
"African swine fever-like viruses" (gen fara denumire) |
Ceratopogonidae |
|
Reoviridae |
ARN dublu catenar, virusuri icosaedrice, fara invelis, 60-80 nm |
Orbivirus Coltivirus |
Ceratopogonidae Phlebotominae Culicidae Ixodidae Ixodidae |
|
Rhabdoviridae |
ARN monocatenar negativ, forma de bacili, cu invelis , 100-430 x 45-100 nm |
Vesiculovirus Ephemerovirus |
Culicidae Phlebotominae (Tabanidae) Culicidae Phlebotominae |
|
Orthomyxoviridae |
ARN monocatenar negativ, virusuri sferice, cu invelis, 80-120 nm |
"Thogoto-like viruses" (gen fara denumire) |
Ixodidae |
|
Bunyaviridae |
ARN monocatenar negativ,virusuri sferice, cu invelis, 80-120 nm |
Bunyavirus Nairovirus Phlebovirus |
Culicidae Ixodidae Argasidae Phlebotominae Culicidae Ixodidae |
|
Flaviviridae |
ARN monocatenar pozitiv, virusuri sferice, cu invelis, 40-50 nm |
Flavivirus |
Ixodidae Culicidae |
|
Togaviridae |
ARN monocatenar pozitiv, virusuri sferice, cu invelis, 70 nm |
Alphavirus |
Culicidae |
Celelalte virusuri produc stari febrile cu simptome clinice. Datorita complexitatii sistemelor ecologice in care circula arbovirusurile preconditiile pentru mentinerea ciclului viral sunt de regula restranse numai in areale limitate. Aceste zone sunt numite focare pentru care virusurile sunt endemice. (Termenii "endemic" si "endemisme" au intelesuri diferite in biologie si medicina. In biologie "endemic" inseamna "restrans la un anumit biotop sau regiune, zona muntoasa, insula etc.". In medicina, "endemic" este mai mult sau mai putin sinonim cu "autohton"). De aceea, amplificarea unui focar este, de regula, dificila. Formarea de noi focare presupune conditii ecologice potrivite. Stabilirea si raspandirea noilor focare presupune introducerea virusului intr-o biocenoza favorabila de catre pasarile migratoare viremice sau de catre mamiferele mari (in particular omul) ce pot actiona ca gazde amplificatoare. Exemple ale modului in care migratiile speciei Homo sapiens au favorizat dezvoltarea a numeroase focare noi pentru infectiile arbovirale in zonele tropicale si subtropicale, sunt febra galbena si denga.
In secolele XVIII si XIX virusul febrei galbene a fost introdus in cateva tari din Sudul Europei (Spania, Portugalia, Italia, Franta) unde s-a stabilit pentru scurte perioade.
In secolul XX virusul Denga I a fost introdus in Grecia unde a produs o mare epidemie in anul 1928. Virusurile Bluetongue au fost introduse si au produs epidemii la animalele domestice din Sudul Europei. Aceasta raspandire antropogenica a infectiilor arbovirale joaca un rol important in toate ciclurile arbovirusurilor in care omul este principala gazda vertebrata.
Urmatoarele familii de artropode cuprind vectori pentru arbovirusuri: Ixodidae, Argasidae, Culicidae, Phlebotominae si Ceratopogonidae. De asemenea, arbovirusurile au fost izolate din alte familii de artropode (capuse, plosnite, purici, simulide, tabanide); totusi nu este clar daca unele dintre ele pot actiona ca gazde naturale si ca vectori biologici.
Structura de baza a ciclului unui arbovirus este prezentata in Figura 4. Pot fi diferentiate doua cai:
De regula, virusul este transmis de catre artropodele hematofage (vectorul si gazda artropod) la un vertebrat la care se produce viremia. In aceasta perioada, ce dureaza de obicei cateva zile, virusul poate fi preluat de catre un alt artropod hematofag. Vertebratul poate sau nu sa se imbolnaveasca la scurt timp, in acelasi timp sau dupa viremie. Astfel, omul poate fi integrat in ciclul arbovirusului si poate dezvolta boala. In unele cicluri arbovirale omul este singura gazda vertebrata ce mentine ciclul virusului (denga, febra galbena urbana, Chikungunya), in altele omul este gazda finala a ciclului viral, chiar daca nu dezvolta boala.
Exista si un alt tip de ciclu in care virusul poate "ramane" in gazda sa artropoda fiind transmis vertical - transovarian - catre generatia urmatoare si apoi transtadial catre stadiile larvare, pupale si la adulti. S-a semnalat si transmisia sexuala ("veneriana") de la mascul la femela si vice versa in timpul copulatiei.
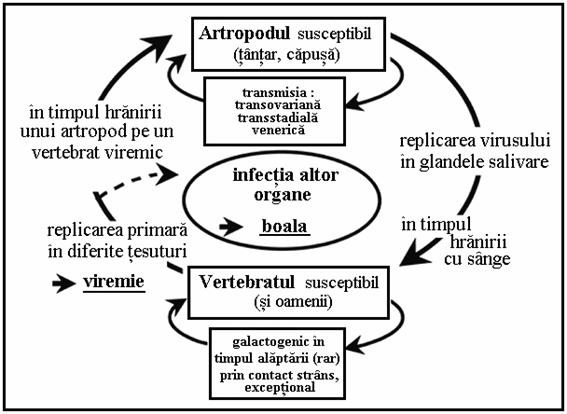
Figura 4. Ciclurile arbovirusurilor.
Transmisia transovariana a fost stabilita comparativ numai pentru un numar mic de arbovirusuri, dar se presupune ca ea este frecventa pentru majoritatea arbovirusurilor, indiferent de tipul vectorului: capuse, tantari, flebotomi etc.
Un mod mai neobisnuit de infectie a fost descris la capuse: capusele neinfectate ce se hranesc simultan cu capuse infectate pe aceiasi gazda si se pot infecta (in absenta unei viremii adevarate a gazdei) prin preluarea virusului transmis de catre capusele infectate, cel mai probabil dupa replicarea in unele celule ale pielii. Este posibil ca acest mecanism, ce nu este inca bine inteles, sa fie de mare semnificatie pentru arbovirusuri.
Virusurile ingerate infecteaza mai intai celulele epiteliale ce captusesc intestinul mediu; apoi sunt eliberate treptat in hematocel de unde pot infecta glandele salivare si ovarele. Susceptibilitatea epiteliului din intestinul mediu este determinantul primar pentru competenta vectoriala a unui artropod.
Infectia glandelor salivare se poate produce dupa amplificarea secundara in alte celule sau fara o amplificare secundara.
Perioada dintre ingestia virusului si secretia salivara se numeste perioada de incubatie extrinseca PIE. Acest interval nu trebuie sa depaseasca durata de viata a unui artropod ce actioneaza ca vector potential. La unele specii de alfavirusuri tesuturile intestinului mediu sunt puternic afectate de catre infectie incat sunt "perforate", fapt ce permite virusului sa intre in hematocel si apoi sa infecteze rapid glandele salivare. Aceasta duce la o perioada de incubatie extrinseca neobisnuit de scurta.
Temperatura are o mare influenta asupra duratei PIE care, de regula, este de cateva zile; cresterea temperaturii poate duce la cresterea ratelor de transmisie. Coinfectiile cu alti patogeni pot, de asemenea influenta astfel: microfilariile ce trec prin epiteliul intestinului mediu cauzeaza leziuni care faciliteaza transferul virusului catre hematocel. Transferul virusului de la tantar la vertebrat se produce, de regula, in timp ce acesta tatoneaza tesuturile cu ajutorul pieselor bucale in cautarea capilarelor sanguine. Cel mai adesea prima replicatie are loc la nivelul tesuturilor extravasculare.
Alti factori ce influenteaza capacitatea unei anumite specii de artropod pentru ciclul virusului sunt: cantitatea de virus din glandele salivare, numarul de generatii, densitatea si dinamica populatiilor, frecventa si durata hranirii cu sange, distributia orizontala si verticala, ritmurile circadiene si spectrul gazdelor. Dintre toti acesti factori, numarul indivizilor capabili de realizarea transmisiei intr-o anumita biocenoza pare sa fie parametrul cheie.
Gazdele vertebrate ale arbovirusurilor cuprind numeroase mamifere pe de o parte si pasarile pe de alta; in putine cazuri reptilele si amfibienii sunt (sau pot fi) importanti.
Capacitatea unei anumite specii vertebrate de a fi o gazda importanta pentru mentinerea ciclului depinde de mai multi factori, cel mai important fiind concentratia virusului in timpul viremiei precum si lungimea perioadei de viremie, precerinte pentru infectarea cu succes a unui mare numar de vectori.
Alti parametri sunt: densitatea si dinamica populatiilor, ratele de reproducere si speranta de viata, gradul de imunizare al populatiei, activitatea pe orizontala si pe verticala, ritmul circadian si spectrul de paraziti hematofagi precum si numarul de vertebrate gazda susceptibile sa devina viremice.
Luand in considerare toti acesti factori rezulta ca avem de fapt de a face cu un sistem ecologic foarte complex.
La arbovirusurile ce apar in zonele temperate o importanta problema ecologica legata de ciclul virusului se datoreaza existentei iernii si deci a hibernarii. Aceasta perioada poate fi depasita prin cateva mecanisme: transmisia verticala in cazul gazdelor artropode supravietuirea se face in stadiile hibernante (larve sau adulti) ale artropodelor; infectiile cronice ale gazdelor vertebrate cu viremie persistenta sau recrudescenta sau prin introducere regulata de catre pasarile migratoare (cu o viremie prelungita) ce vin din regiunile tropicale.
Hibernarea virusurilor transmise de catre capuse este de regula realizata mai usor datorita persistentei virusului in larve si nimfe si prin transmisia transtadiala. La unele virusuri transmise de catre tantari (Anopheles sp., Culiseta sp., Culex sp.) hibernarea este de asemenea posibila in adulti.
Cu toate ca transmisia verticala transovarina si sexuala sunt fenomene biologice remarcabile si mecanisme importante, se pare ca mentinerea virusului in unele biocenoze se poate realiza si prin moduri ce implica participarea mamiferelor in circulatia virusurilor.
O cale naturala este transmisia galactogena realizata in timpul alaptarii descendentilor sau la om (in cazul animalelor al carui lapte este consumat). Astfel de cazuri s-au semnalat pentru encefalita de capusa transmisa la oameni prin consumul laptelui infectat. In lapte virusul este gasit numai in timpul perioadei de viremie la animalul care alapteaza.
Contactul cu animalele afectate (vii sau moarte) poate duce, de asemenea, la infectie, faptul a fost demonstrat la vertebratele ce au fost vanate sau omorate (transmisia virusului febrei Rift Valley la om).
De asemenea exista virusuri ce nu au fost niciodata izolate de la un artropod si care aparent sunt transmise intre gazdele lor vertebrate numai prin contactul strans dintre ele (Modoc, Rio Bravo). Aceste virusuri nu sunt arbovirusuri prin definitie; ele au fost totusi incluse in aceasta categorie datorita relatiilor antigenice foarte stranse cu alte virusuri care prezinta toate caracterele unui arbovirus real.
Din punctul de vedere al rolului vertebratelor in circulatia arbovirusurilor, acestea pot fi impartite in trei grupe:
- specii care nu sunt susceptibile si in care nu se produce replicatia virusurilor;
- specii ce sunt susceptibile dar care dezvolta o viremie redusa (si de scurta durata) nefiind insa suficienta pentru infectarea cu succes a artropodelor. Aceste specii nu sunt semnificative pentru mentinerea ciclului viral;
- specii ce sunt susceptibile si care dezvolta o viremie suficient de ridicata si infecteaza pe termen lung artropodele. Acestea sunt speciile ce mentin ciclul virusului.
In afara de intensitatea si de durata viremiei, speciile din ultimele doua grupe pot, dar nu este obligatoriu, sa dezvolte o boala. Acele vertebrate care sunt esentiale pentru mentinerea ciclului pot sa nu dezvolte boala; in unele cazuri ele nu prezinta nici un simptom. Aceasta pare sa fie in acord cu consideratiile generale asupra parazitilor; de asemenea, trebuie tinut cont de faptul ca un vertebrat infectat cu un virus poate dezvolta imunitate, fapt ce previne reinfectia. De aceea, o gazda vertebrata poate contribui la ciclul viral direct in timpul stadiului sau viremic numai pentru o scurta perioada de timp. Dupa aceea (in afara cazurilor rare de infectii cronice cu viremie persistenta sau intermitenta), vertebratul poate sa nu fie o sursa de infectie. Totusi, el poate contribui indirect la circulatia virusului, actionand ca o gazda pentru artropodele care mentin circulatia (capusele care ating populatii ridicate daca gazdele sunt abundente). Speciile de vertebrate ce dezvolta imbolnaviri severe cu grad ridicat de mortalitate pot actiona ca gazde de mentinere, in special daca au densitati populationale ridicate si cu un procent suficient de indivizi non-imuni. Denga si febra galbena (in zonele urbane) sunt bune exemple ale circulatiei dintre Culicidae (Aedes aegypti) pe de o parte, si om, de cealalta parte.
Dintre cele 600 arbovirusuri descrise pana in prezent, aproximativ 150 s-au dovedit patogene pentru om. In unele infectii au fost identificati numai anticorpii, fara alte simptome care sa indice ca omul este susceptibil.
Sindroamele cauzate de catre infectiile arbovirale pot fi impartite in:
stari febrile de regula cu dureri de cap, eruptii, cu sau fara artralgie si poliartrita (Denga, West Nile, Chikungunya, Mayaro, Ockelbo, O'nyong-nyong, Ross River, Semliki, Sindbis si altele);
febre hemoragice (Crimea-Congo, Omsk, boala padurii Kyasanur, Denga, Febra galbena);
deranjamente neurologice cu meningita si encefalite (boala padurii Kyasanur, Powassan, encefalita de capusa, encefalita equina de est, encefalita equina Venezueleana, encefalita equina de vest si altele).
Infectiile cu arbovirusuri au un impact puternic asupra sanatatii umane peste tot in lume. In fiecare an peste 100.000 de oameni (posibil milioane) sunt infectati de catre arbovirusuri si cateva mii se imbolnavesc. In fiecare an, sunt raportate epidemii mai mici sau mai mari datorate virusurilor (denga, febra galbena, Chikungunya, encefalita Japoneza, Rift Valley etc.) - cu un numar variabil de decese.
De asemenea, sunt serioase pierderile economice datorate infectiei cu arbovirusuri a efectivelor de animale domestice. Unele infectii arbovirale au avut un rol considerabil pentru istoria umana. Febra galbena (care se pare ca a fost introdusa in America in timpul transporturilor de sclavi din Africa) a dus la infrangerea armatei franceze in Caraibe la inceputul secolului XIX astfel ca Napoleon a fost fortat sa cedeze Louisiana la U.S.A., fapt ce a dus la sfarsitul influentei Frantei in America de Nord. In anii 1880, febra galbena a impiedicat constructia canalului Panama.
Vigilenta constanta si supravegherea sunt componentele importante in reducerea impactului virusurilor pentru sanatatea publica.
Nu exista medicamente specifice pentru tratamentul infectiilor arbovirale, este posibil numai tratamentul simptomatic.
Exista vaccinuri numai pentru urmatoarele arbovirusuri aflate in circulatie: virusul encefalitei de capusa, cel al encefalitei Japoneze si pentru virusul febrei galbene. De asemenea, au fost produse cateva vaccinuri "ne-comerciale" folosite pentru protectia lucratorilor din laboratoare si din domeniul veterinar. Vaccinul pentru Denga a fost realizat si se afla in stadiul testelor de teren.
Bibliografie
Arnell, J. H. - 1973. Mosquito studies (Diptera, Culicidae) XXXII. A revision of the genus Haemagogus. Contributions of the American Entomological Institute, 10(2): 1-174.
Belkin, J. N. - 1962. The mosquitoes of the South Pacific. Berkeley, University of California Press. Vol. I, 608 pp; Vol. II, Plates 1-412.
Brian, G.W., ed. - 1981. Handbook series in zoonoses, Section B. Viral zoonoses. Vol.1, 510 pp.
Brown, A.W.A. - 1977. Yellow fever, dengue and dengue haemorrhagic fever. pp. 271-317. In: Howe, G.M. ed. A world geography of human diseases, London, Academic Press, 621 pp.
Calisher, C. H. & Thompson, W.H. eds. - 1983. California serogroup viruses. New York, Liss.
Christophers, S. R. - 1960. Aedes aegypti, the yellow fever mosquito. Cambridge, University Press, 739 pp.
Halstead, S. B. - 1980. Dengue haemorrhagic fever - a public health problem and a field for research. Bulletin of the World Health Organization, 58: 1-21.
Harwood, R . F. & James, M .T. - 1979. Entomology in human and animal health. New York, MacMillan, 548 pp.
Karabatsos, N. ed. - 1985. International catalogue of arboviruses, including certain other viruses of vertebrates. 3rd Ed. Baltimore, American Society of Tropical Medicine and Hygiene, 1147 pp.
Khin, M. M. & Thraw, K. A. - 1983. Transovarial transmission of dengue 2 viruses by Aedes aegypti in nature. American Journal of Tropical Medicine and Hygiene, 32: 590-594.
Knight, K. L. & Stone, A. - 1977. A catalogue of the mosquitoes of the world (Diptera: Culicidae). (Thomas Say Foundation Vol. Vl). College Park M.D. Entomological Society of America, 611 pp.
Knudsen, A. B. - 1977. The silent jungle transmission cycle of dengue virus and its tenable relationship to endemic dengue in Malaysia. The Malayan Nature Journal, 31: 41-47.
Leake, C. J. - 1988. Strategies for vector-borne disease control in rice production systems in developing countries: arboviruses other than Japanese encephalitis. In: Vector-bome disease control in humans through rice agroecosystem management. International Rice Research Publication, in collaboration with the WHO/FAO/UNEP Panel of Experts on Environmental Managemen for Vector Control, pp.161 173.
Macdonald, W. W. - 1976. Mosquito genetics in relation to filarial infections. pp. 1-24 In: Taylor A.E.R. & Muller, R. ed. Genetic Aspects of host-parasite relationships. (Symposium of the British Society for Parasitology, Vol. 14). Oxford, Blackwells, pp. 1-24.
Monath, T. P. - 1979. Arthropod-borne encephalitides in the Americas. Bulletin of the World Health Organization, 57: 513-533.
Monath, T. P. - 1980. St. Louis encephalitis. Washington D.C., American Public Health Publication,.
PAHO. Dengue outbreak in Boa Vista, Roraima Territory, Brazil. Epidemiological Bulletin, 3 (6): 5-7 (1982).
Rosen, L. et al. - 1983. Transovarial transmission of dengue viruses by mosquitoes: Aedes albopictus and Aedes aegypti. American Journal of Tropical Medicine and Hygiene, 32, 1108-1119.
Rudnick, A. - 1983. The ecology of the dengue virus complex in Peninsular Malaysia. In: Pang, T. & Pathmanathan, R. Proceedings of the International Congress on Dengue/ Dengue Haemorrhagic Fever, Kuala Lumpur, University of Malaya, pp. 7-14.
Sudia, W. D., Newhouse, V. F. - 1975. Epidemic Venezuelan Equine Encephalitis in North America: a summary of virus-vector-host relationships. American Journal of Epidemiology, 101: 1-13,.
Watts, D.M., De Foliart, G.R., Yuill, T.M. - 1976. Experimental transmission of Trivittatus Virus (California Virus group) by Aedes trivittatus. American Journal of Tropical Medicine and Hygiene, 25: 173-176.
White, G.B. - 1987. Medical entomology: mosquitoes. In: Manson-Bahr, P.E.C. & Bell, D.R. eds. Manson's Tropical Diseases, 19th Ed. London, Bailliere Tindall, pp. 1404-1435.
WHO. - 1980. Overall review of yellow fever in Africa and America, 1965-1979. Weekly Epidemiological Record, 55:356-360.
Copyright © 2024 - Toate drepturile rezervate