|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |

Vectorii
Notiuni generale despre artropode
Insectele: Ordinul Diptera, Ordinul Anoplura, Ordinul Siphonaptera, Ordinul Hemiptera, Ordinul Dictyoptera
Acarienii: Acarina, Argasidae, Ixodidae
Notiuni generale despre artropode
Artropodele, ce corespund unui tip de organizare bine definit, constituie una dintre increngaturile cele mai importante ale regnului animal atat ca numar de specii (80-85% din speciile animale cunoscute), cat si prin numarul de indivizi.
Numarul de specii descrise depaseste un milion, iar inventarierea nu s-a terminat inca. Dintre acestea, numai categoria insectelor reprezinta trei sferturi dintre artropode. Mai mult, prin varietatea lor morfologica si datorita marii lor plasticitati ecologice, aceste nevertebrate in particular insectele au reusit sa colonizeze majoritatea habitatelor naturale, adaptandu-se la un mare numar de moduri de viata.
In cele ce urmeaza vor fi prezentate cateva notiuni generale referitoare la morfologia, dezvoltarea si clasificarea artropodelor.
Morfologie si dezvoltare
Sub aspect morfologic, artropodele sunt caracterizate de prezenta unei cuticule (cel putin la stadiul de adult) ce inveleste la exterior intregul animal, indeplinind rolul unui schelet extern. Aceasta cuticula rigida (exceptie la nivelul membranelor articulare unde isi pastreaza supletea) este alcatuita din straturi de chitina (acetatul unui polizaharid al carui element principal il reprezinta glucozamina) si o proteina hidrosolubila (artropodina). Prezenta membranelor articulare asigura mobilitatea diferitelor segmente ale corpului unele in raport fata de celelalte, precum si a articulatiilor diverselor segmente ale apendicelor (piese bucale, picioare, gonopode). Aceasta particularitate sta la baza denumirii artropodelor (din greaca "arthron" = articulatie si podos = picior).
Existenta scheletului extern rigid implica o crestere discontinua ce se realizeaza prin naparliri succesive.
Corpul artropodelor este alcatuit dintr-o serie de elemente relativ asemanatoare, dispuse cap la cap, denumite segmente, somite sau metamere.
Unele dintre aceste segmente s-au reunit pentru a forma regiuni sau tagmata (tagma) ce au o individualitate proprie. Aceste regrupari de segmente nu au un aspect uniform pentru toate clasele de artropode si de aceea s-au adoptat denumiri particulare pentru fiecare situatie in parte. La insecte distingem cap, torace si abdomen; la crustacee sunt utilizati termeni ca cephalon, percion si pleon; la chelicerate, ce nu au niciodata o regiune cefalica individualizata, avem de a face cu: prosoma (cefalotorace la aranee si scorpioni) si opistosoma pentru regiunea posterioara. Folosirea acestor termeni are avantajul excluderii recunoasterii implicite a omologiilor inexacte.
Divizarea corpului in segmente, precum si regruparile in tagmata poate fi profund modificata la diferitele grupe datorita fenomenelor de fuziune sau de reducere. In plus, se pot produce modificari evolutive marcante la nivelul apendicelor segmentelor datorita pierderii unor functii (astfel picioarele ambulatoare pot sa indeplineasca functii noi senzoriale, masticatoare, prehensile sau se pot specializa pentru functiile de reproducere). De aceea, pentru studiile de morfologie se impune o analiza separata a diferitelor ordine.
Pe planul organizarii interne artropodele prezinta urmatoarele caracteristici importante:
Tubul digestiv este format din trei regiuni: una anterioara sau stomodeum si una posterioara sau proctodeum, ambele de origine ectodermica; regiunea mediana sau mesenteron este de origine endodermica; functia excretoare este asigurata de coeca si tuburile Malpighi ce se deschid la limita mezenteronului cu proctodeum.
Un element important al tubului digestiv este reprezentat de catre membranele peritrofice (MP) ce se regasesc la cateva dintre filumurile regnului animal. In general este acceptat faptul ca MP, sunt alcatuite din mai multe straturi, ce contin proteine, mucopolyzaharide si microfibrile chitinoase; ulterior pot sa apara trasaturi specifice ale texturii care difera chiar la larvele si adultii aceleiasi specii. Aceste microfibrile par sa protejeze epiteliul intestinal de particulele dure de hrana si intr-o anumita masura de patogeni. Acest fapt este important pentru abilitatea unor artropode ectoparazite ce actioneaza ca vectori pentru endoparaziti. De aceea este foarte probabil ca parazitii si alti patogeni sa poata trece prin MP numai in momente si prin zone particulare. Spre exemplu, ookinetii mobili ai speciilor de Plasmodium ce se formeaza foarte rapid (4-9 h) dupa ingerarea sangelui de catre femelele de Anopheles spp. pot trece prin MP incomplet formata si solidificata (PLANSA 8); acest proces necesita cel putin 13 ore in Anopheles gambiae si 32 h in Anopheles stephensi care sunt vectori foarte comuni pentru parazitii malariei umane. Aceleasi aspecte par sa fie valabile si in cazul trypanosomelor ce ajung in intestinul mustelor tsetse (Glossina spp.), la care formarea continua a MP se declanseaza numai dupa emerjare; astfel tinerele muste ce pot avea un pranz infectiv la 15 h de la iesirea din puparium, ajung la rate de infectare a glandelor salivare de pana la 24%, pe cand mustele mai batrane sunt infective cu o rata mult mai scazuta (0,04% - 1%). Probabil, MP actioneaza ca un ultrafiltru selectiv ce are ca efect reducerea considerabila a incarcarii parazitare si deci diminuarea efectelor patologice asupra insectelor gazda.
La modul general, formarea MP se poate realiza pe doua cai principale. Fie prin delaminarea in intregime a epiteliului intestinului mediu, secretarea de catre partea anterioara fiind minora. Acest tip de formare a MP se produce la adultii de Tabanomorpha si Nematocera cand primul pranz sangvin incomplet digerat este inclus in cel de al doilea care la randul sau este invelit de o MP. De aceea, parazitii sau patogenii inclusi in primul pranz au de strapuns doua membrane peritrofice.
Al doilea mod de formare al MP - prin secretie este intalnit la toate larvele de diptere si la adultii de Muscomorpha (Muscidae, Hippoboscidae, Calliphoridae, Sarcophagidae, Oestridae). Numai o zona scurta cu cateva randuri de celule separate situata la debutul intestinului mediu este capabila sa produca un tub asemanator cu MP. La majoritatea larvelor de nematocere este secretata numai o singura MP. Adultii muscomorfelor au cateva MP cu origini distincte si din zone celulare separate (Plansa 8).
Zonele de formare si valvula cardiaca sunt cunoscute de regula ca proventricul, lucru incorect deoarece adevaratul proventricul este numai o portiune din intestinul anterior. Totusi aceasta regiune poate fi numita cardia daca formeaza MP fara alte delaminari aditionale ale epiteliului intestinului mediu adiacent.
La unele artropode de importanta medicala, MP lipseste complet (spre exemplu toate stadiile de dezvoltare ale plosnitelor si ale paduchilor ca si la puricii adulti). Capusele si acarienii hematofagi nu au in general MP, exceptie fac unele specii de Ixodes sp., ce prezinta un strat de celule intestinale asemanator MP, fapt ce face ca digestia lor sa difere semnificativ de a celorlalte capuse.
Aparatul circulator, foarte redus in general, nu este inchis, sangele "hemolimfa" ce pare sa aiba mai ales rol nutritiv este difuzat in cavitatea generala a corpului (hemocoel) printr-un organ propulsor vasul dorsal ("inima") la nivelul caruia se deschid orificii numite ostiole.
Sistemul nervos este format dintr-un "creier" dorsal, dintr-un inel peri-esofagian si din lantul nervos dublu, situat ventral, ce are cate o pereche de ganglioni pentru fiecare segment.
Aparatul respirator variaza mult in functie de increngatura. Majoritatea artropodelor acvatice respira prin branhii; rar ele pot avea si o respiratie cutanata. Artropodele terestre respira cu ajutorul "plamanilor" sau "trahee".
Musculatura artropodelor este de tip striat (in mare parte) si prezinta un aranjament caracteristic. Muschii nu acopera corpul in lungimea lui ci sunt aranjati pe segmente si se insera la nivelul tendoanelor dintre segmente. Legaturile musculare pot actiona in toate directiile dar in unele cazuri au activitati antagonistice. Extremitatile (ca piesele bucale, picioarele si aripile cand sunt prezente) au muschi independenti de cei ai corpului. In plus, fata de acesti muschi striati cu actiune rapida, intestinul, alte organe interne si apendicele externe sunt insotite de muschi netezi ce contin relativ putine filamente.
Clasificarea
Clasificarea zoologica este aranjarea ierarhica a categoriilor taxonomice intr-o ordine naturala (este de presupus). Acesta este obiectul unei discipline particulare: sistematica, ce are la baza un ansamblu de principii si de reguli ce constituie taxonomia.
Este prezentata o cheie pictoriala simpla pentru clasele si ordinele majore de artropode adulte de importanta medicala (Plansa 6
Marea diversitate observata in structurile morfologice ale artropodelor au condus la subdivizarea increngaturii intr-un numar de clase, ordine si familii. Din numeroasele sisteme existente, in lucrarea de fata, este utilizata cea propusa de Brusca & Brusca (2002).
Filumul ARTHROPODA Brusca & Brusca (2002).
Subfilumul TRILOBITA (=TRILOBITOMORPHA). Extinct ~ 4,000 specii.
Subfilumul CHELICERIFORMES (=CHELICERATA).
Clasa CHELICERATA (=EUCHELICERATA).
Subclasa MEROSTOMATA.
Ordinul Xiphosura 5 specii; Ordinul Eurypterida. Extinct
Subclasa ARACHNIDA.
Ordinul Palpigradi ~ 60 specii; Ordinul Uropygi ~ 100 specii; Ordinul Amblypigi ~ 70 specii; Ordinul Ricinulei 35 specii; Ordinul Scorpiones ~ 1.200 specii; Ordinul Pseudoscorpionida ~ 2.000 specii; Ordinul Solpugida (=Solifugae) ~ 900 specii; Ordinul Araneae ~ 35.000 specii; Ordinul Opiliones (= Phalangida) ~ 5.000 specii; Ordinul Acari ~ 30.000 specii.
Clasa PYCNOGONIDA. 1.000 specii.
Subfilumul CRUSTACEA >30.000 specii, include Pentastomida cu pozitie filogenetica nesigura. Daca sunt incluse si Hexapoda = PANCRUSTACEA sau TETRACONATA.
Subfilumul UNIRAMIA nu este valid clade, datorita pozitiei problematice a Crustacea. Apelate si ATELOCERATA sau TRACHEATA.
Clasa MYRIAPODA
Subclasa CHILOPODA ~ 2.500 specii.
Subclasa DIPLOPODA ~ 10.000 specii.
Subclasa SYMPHYLA. ~ 120 specii.
Subclasa PAUROPODA. ~ 500 specii.
Superclasa HEXAPODA
Clasa COLLEMBOLA (Oligentomata) ~ 6,000 specii.
Clasa PROTURA (Myrientomata) ~ 500 specii.
Clasa DIPLURA (Diplurata) ~ 800 specii.
Clasa INSECTA. > 800,000 specii.
Subclasa Apterygota (Zygentomata).
Ordinul Archaeognatha [Microcoryphia] ~ 500 specii; Ordinul Zygentoma [Thysanura] ~ 400 specii.
Subclasa Pterygota
Infraclasa Paleoptera
Ordinul Ephemeroptera [Ephemerida] ~ 3.000 specii; Ordinul Odonata ~ 5.500 specii.
Infraclasa Neoptera
Divizia Exopterygota, Neoptera cu metamorfoza incompleta - Hemimetabole
Include insectele orthopteroide : Ordinul Isoptera ~ 2.800 specii; Ordinul Dictyoptera ~ 4.300 specii (subordinul Blattodea, subordinul Mantodea); Ordinul Dermaptera ~ 2.000 specii; Ordinul Grylloblattaria (Grylloblattodia) ~ 27 specii; Ordinul Plecoptera ~ 2.000 specii; Ordinul Orthoptera ~ 21.000 specii; Ordinul Phasmatodea ~ 3.000 specii; Ordinul Embioptera (Embiidina) ~ 300 specii; Ordinul Zoraptera ~ 30 specii; Ordinul Mantophasmatodea ~ 13 specii si cele hemipteroide Paraneoptera (Acercaria) ce includ ordinele: Ordinul Psocoptera ~ 3.000 specii, Ordinul Phthiraptera ~ 5.000 specii, Ordinul Thysanoptera ~ 5.500 specii; Ordinul Hemiptera ~ 90.000 specii.
Divizia Endopterygota. Neoptera cu metamorfoza completa - Holometabole.
Include: Ordinul Neuroptera ~ 6.000 specii; Ordinul Megaloptera ~ 300 specii; Ordinul Raphidioptera ~ 350 specii, Ordinul Coleoptera ~ 350.000 specii; Ordinul Strepsiptera ~ 550 specii; Ordinul Diptera ~ 125.000 specii; Ordinul Mecoptera ~ 600 specii; Ordinul Siphonaptera ~ 2.500 specii; Ordinul Trichoptera ~ 11.000 specii; Ordinul Lepidoptera ~ 160.000 specii; Ordinul Hymenoptera ~ 100.000 specii. (cu rosu sunt grupele de importanta medicala)
PLANSA 6.
Cheie pictoriala
pentru clasele si ordinele majore de artropode
adulte de importanta medicala
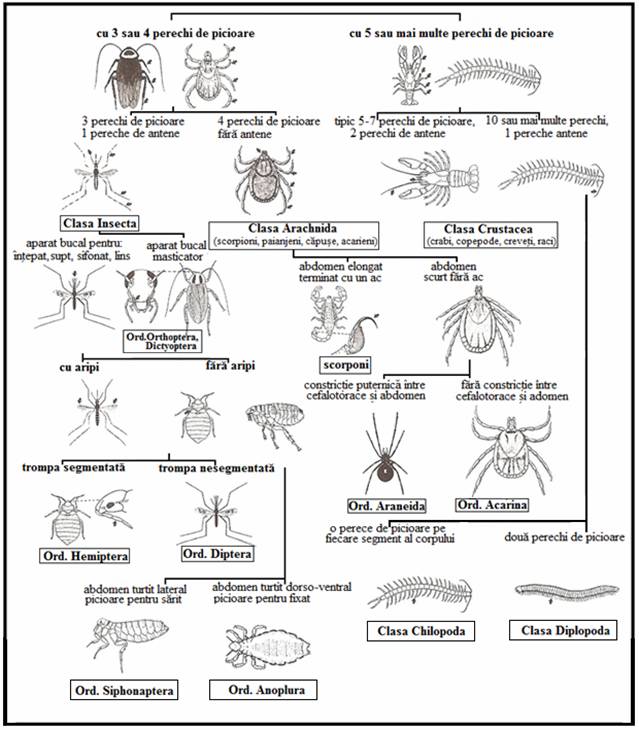
(adaptare dupa Harry D. Pratt & Chester J.Stojanovich)
Insectele constituie grupul cel mai mare de animale sub aspectul numarului de specii (~ 773,000) si ca numar de indivizi. Clasificarea lor se bazeaza pe prezenta (Pterygota) sau absenta (Apterygota) aripilor. Subclasa Pterygota ce s-a dezvoltat mai tarziu include toate speciile parazitice importante; unele dintre ele si-au pierdut in aparenta aripile primare ca urmare a adaptarii lor la parazitism (puricii).
Insectele pot actiona ca ectoparazite, atunci cand se hranesc cu sange la suprafata corpului gazdelor lor (tantarii) sau pot fi endoparazite, cand patrund in pielea sau in tracturile intestinale si/sau respiratorii ale diferitelor gazde (mustele ce provoaca miaze; Dermatobia hominis). In plus, insectele mai pot fi implicate direct sau indirect in ciclurile de viata ale unui mare numar de agenti patogeni (paraziti).
Morfologie externa si dezvoltare
Organizarea corpului insectelor parazitice este cel mai adesea puternic adaptata la modul lor particular de viata si pentru nevoile speciale legate de modul de hranire.
Corpul insectelor adulte este divizat in trei regiuni bine diferentiate: cap (caput), torace (thorax) si abdomen (trunk), fiecare parte consta din cateva segmente specifice (vizibile sau nu din exterior) (Plansa 7).
La exterior se afla tegumentul (cuticula, exoscheletul) ce acopera intregul corp al insectelor precum si portiunea anterioara si posterioara a intestinului (Plansa 8). Masa cuticulara inerta este excretata de catre epiderma (hipoderma) care consista dintr-un singur strat de celule situate in lamina bazala (membrana bazala). Hipoderma include o varietate de celule diferite ca celulele epidermice tipice, celulele din care se formeaza perii (celulele tomogene si trichogene), oenocitele, celulele senzoriale si diferite tipuri de celule ale glandelor dermale. Acestea pot forma protruzii tubulare citoplasmatice lungi ce se extind pana aproape de suprafata (porii cuticulari). Celulele hipodermice "normale" produc cuticula inerta care este compusa din trei straturi distincte: epi-, exo- si endocuticula. Stratul cel mai intern, endocuticula, subtire, include filamente chitinoase si proteine incolore si ramane flexibila. Exocuticula de asemenea relativ subtire, reprezinta principala componenta a exoscheletului deoarece este alcatuita din chitina si proteine colorate (sclerotina). La exterior exocuticula este acoperita de epicuticula care in general are o grosime de 1-3 μm; epicuticula este compusa dintr-un strat intern de lipoproteina (cuticulina), un strat de polifenol, un strat cerat si in final este acoperita de un strat de ciment, toate asigurand impermeabilitatea si supravietuirea in atmosferele nesaturate in apa.
Aceasta constructie tipica a cuticulei se schimba regulat prin naparlire si poate fi alterata in zonele in care este nevoie de mai multa flexibilitate. De aceea, membranele dintre sclerite (segmente) sunt lipsite de o exocuticula rigida iar endo- si epicuticula raman netede pentru a permite indoirea corpului. Naparlirea se produce datorita activitatii unor hormoni ca ecdysona, neotenina etc.
Capul ale carui segmente formeaza o capsula puternica, poarta dorsal o pereche de antene segmentate si ochii. In general, acestia sunt simpli (oceli) sau compusi si sunt dispusi aproape de baza antenelor; ei sunt alcatuiti din numeroase omatidii iar in rare cazuri (purici) sunt prezente una sau cateva omatidii.
Ventral se gasesc piesele bucale adaptate pentru modalitatile speciale de hranire. La insectele parazite piesele bucale se organizeaza in mai multe tipuri, in functie de regimul alimentar (Plansa 9).
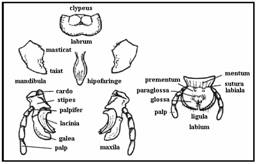
Tipul masticator este considerat tipul de baza de la care au evoluat celelalte tipuri. Este comun multor specii ce traiesc liber (gandaci, furnici etc.) si este caracteristic pentru majoritatea larvelor. El este alcatuit din labrum, o pereche de mandibule mari (pentru masticarea hranei), o pereche de maxile (ce servesc la impingerea in gura a fragmentelor de hrana) si din labium.
Tipul absorbant pentru supt) sau pentru lins este intalnit la majoritatea dipterelor neintepatoare (Musca, Calliphora). Mandibulele si maxilele sunt nefunctionale, partile ramase formeaza un proboscis cu o buza largita in suprafata ce consta din doua formatiuni ce inconjoara gura (labella). Hrana dizolvata de catre secretiile salivare este ingerata in forma lichida via canalele capilare superficiale, care o conduc catre gura.
Tipul taietor, absorbant sau lins este caracteristic tabanidelor si glosinelor. Mandibulele lor sunt modificate in niste lame taioase iar maxilele au forma unor stileti lungi; ambele pot taia pielea gazdei. Sugerea sangelui se produce cu ajutorul unui labium asemanator unui burete asociat cu hipo- si epifaringele.
Tipul pentru intepat si supt este prezent la o serie de ectoparaziti hematofagi ca tantarii, musca tsetse, alte muste, paduchi, plosnite si purici. Modificarile intalnite la diferitele grupe sunt asa de mari incat omologiile dintre piesele bucale pot fi regasite foarte rar. In orice caz, daca doua canale diferite sunt formate de catre piesele bucale, cel mai larg dintre ele este folosit pentru a conduce hrana iar celalalt conduce saliva, ce contine un anticoagulant precum si alte substante.
Marimea si forma pieselor bucale cu rol in strapungerea pielii este legata de cele doua modalitati de hranire cu sange. Piesele bucale ale insectelor ce se hranesc cu sange din vasele capilare (solenofagie) unele plosnite (Cimex spp., Rhodnius spp.), puricii, unii tantari (Anopheles spp.) patrund in lumenul capilarelor de calibrul dorit. In cazul insectelor care se hranesc prin baltirea sangelui (telmofagie pool feeders majoritatea nematocerelor (Simuliidae), unele muste (Stomoxys spp., Glossinidae) tabanide si capuse acestea distrug vasele de sange periferice prin taierea lor cu ajutorul pieselor bucale conformate ca niste lame si asteapta ca la locul taieturii sa se stranga o cantitate suficienta de sange ce este ingerat rapid.
La plosnite, purici, paduchi, tabanide si unele muste (muscide, musca tsetse) ambele sexe sunt hematofage, pe cand la nematocere (culicide, simulide, flebotomi si ceratopogonide) numai femelele sunt hematofage.
Toracele este alcatuit din trei segmente (pro-, mezo- si metatorace), fiecare poarta ventral o pereche de picioare. Aceste picioare articulate sunt compuse din cinci parti distincte (coxa, trohanter, femur, tibie si tars); tarsul este alcatuit din 1-5 segmente si este echipat cu sisteme de fixare specifice speciilor (gheare, carlige etc.). Mezo- si/sau metatoracele poate purta aripi membranoase tipice (formate de catre integument) ce se misca cu ajutorul unor puternici muschi interni. Uneori aripile sunt reduse secundar la unele grupe (purici, paduchi, plosnita de pat).
Abdomenul. Segmentele abdominale (11 la formele primitive, 10 la cele evoluate) nu formeaza extremitati ventrale cu exceptia unor apendice copulatoare specifice. In interiorul abdomenului se gasesc o serie de sisteme importante ale insectei (sistemul digestiv, sistemul nervos, sistemul circulator, sistemul excretor (tuburile Malpighi, gonadele etc.).
Dimorfismul sexual este evidentiat de catre dezvoltarea apendicelor cu rol in copulatie. In general, deschiderea genitala a femelelor este situata ventral la marginea posterioara a segmentului 8 (sternit) iar in cazul masculilor aceasta este situata de-a lungul liniei mediane ventrale a segmentului 9 ce poarta si apendicele copulatoare.
Organizatia interna a insectelor
Functiile de relatie sunt indeplinite de: musculatura, sistemul nervos si organele de simt;
Functiile de nutritie sunt indeplinite de: sistemul digestiv, sistemul circulator si sistemul respirator;
Functiile de reproducere sunt indeplinite de sistemul reproducator.
Organizatia interna a corpului insectelor cuprinde urmatoarele sisteme principale: tegumentul, muscular, digestiv, circulator, respirator, excretor, nervos si reproducator. La exteriorul corpului se afla tegumentul iar in interior cavitatea generala mixta (mixocel) in care se afla organele interne (Plansa 7
Doua diafragme (dorsala si ventrala) fibro-musculare, impart mixocelul in trei sinusuri longitudinale:
sinusul pericardial - dispus dorsal;
sinusul perivisceral - situat median;
sinusul perineural - situat ventral.
Diafragmele sunt evidente numai in abdomen iar sinusurile comunica intre ele facand astfel posibila circulatia sangelui in spatiile dintre organe.
Sistemul digestiv este format din tubul digestiv si organele anexe.
Tubul digestiv primeste, prelucreaza si transporta hrana, el este un tub asimetric ce strabate toata lungimea corpului si consta din urmatoarele regiuni (Plansa ):
stomodeum (intestinul anterior);
ventriculus (intestinul mediu), si
proctodeum (intestinul posterior).
La partea anterioara a stomodeumului se deschide gura care este localizata in cavitatea preorala si este conectata cu canalele excretoare ale perechilor de glande salivare. Gura este inarmata (original) cu trei perechi de apendice ale capului modificate ca piese bucale specifice cu specia si adaptate la modalitati particulare de hranire. Gura continua cu faringele care eventual actioneaza ca o pompa musculara si conduce hrana prin esofag catre gusa ce actioneaza ca un sistem de depozitare. Gusa care la diptere este un diverticul (fund de sac) ce inchide esofagul, se deschide printr-o valva ingusta proventricul, ce previne regurgitarea hranei din intestinul mediu.
Stomodeumul intestinul anterior) are peretii interni alcatuiti dintr-o membrana cuticulara (intima) si din muschi netezi. Este alcatuit din cinci segmente: gura, faringe (mai dezvoltat la insectele sugatoare, aici se deschid glandele salivare), esofag are glande digestive , gusa (rezervor de hrana) si proventricul stomacul masticator are perti grosi, chitinosi, cu rol in triturarea si filtrarea hranei.
Ventriculus (intestinul mediu, mezenteron) este principalul organ digestiv (stomacul propriuzis) care la unele insecte este captusit cu un tub intern neaderent alcatuit din componente chitinoase (membrana peritrofica) ce are o mare importanta in transmisia agentilor patogeni. Intestinul mediu poate fi divizat in diferite regiuni cu functii de concentrare, digestive si de absorbtie. Are o portiune anterioara cardia si o valvula cardiaca, aici se gasesc cecumurile gastrice cu rolul de a spori suprafata de absorbtie.
Proctodeumul (intestinul posterior) alcatuit din pilor, intestin subtire si rect, se intinde posterior fata de intestinul mediu de care este separat printr-un sfincter piloric. Principala sa functie este aceea de resorbtie a apei din fecale si urina. Componente ale celei din urma sunt colectate de catre tuburile Malpighi ce se deschid la granita dintre intestinul mediu si cel posterior. Intestinul posterior are de regula o parte intestinala (ileum) continuata de catre colon si rectum, ca in final sa se deschida prin anus localizat ventral). Comunicarea dintre diferitele regiuni ale tubului digestiv se face prin orificii inchise de valvule.
Lungimea si forma tubului digestiv este variabila si depinde foarte mult de tipul de hrana consumata, la speciile fitofage fiind mai lung iar la cele zoofage este mai scurt.
Glandele anexe ce contribuie la realizarea digestiei sunt urmatoarele:
glandele salivare, secreta saliva, 2-3 perechi, pot lipsi;
cecumurile gastrice secreta enzime, conduc substantele asimilabile in sange;
celulele glandulare hormonale sunt doua: corpora allata (secreta hormoni ce controleaza procese trofice si naparlirea) si corpora cardiaca (influenteaza metabolismul si metamorfoza);
glandele de excretie sunt tuburile lui Malpighi.
Intestinul anterior si cel posterior sunt de origine ectodermica iar intestinul mediu este de origine endodermica.
Sistemul circulator este format din vasul dorsal si sange, este un sistem lacunar deschis.
Vasul dorsal se intinde intre zona posterioara a abdomenului si cap, este inchis posterior si deschis anterior, alcatuit din doua parti:
inima - organul pulsator, situata in abdomen si partial in torace. Inima are mai multe camarute piriforme (ventriculite) ce comunica prin orificii prevazute cu valvule indreptate inainte (sangele circula numai postero-anterior). Sangele din sinusul pericardial patrunde in inima prin niste orificii perechi situate bazal (ostiole si ele au valvule). Aproape fiecare segment abdominal are un ventriculit.
aorta - tub situat anterior, deschizandu-se in zona cefalica.
Sangele reprezinta intre 20-40% din masa corpului, este alcatuit din hemolimfa (plasma), elemente celulare figurate (hemocite - proleucocite, fagocite, leucocite), saruri anorganice, substante organice; de regula este incolor, ocupand spatiile dintre organe ce constituie hemocelul. Asociate circulatiei intalnim ca organite speciale: oenocitele si celulele pericardiale.
Circulatia sangelui este asigurata de miscarile diafragmelor, ale muschilor ventriculitelor, ale muschilor aliformi si ai abdomenului.
Sangele trece din inima in aorta si de aici in capsula cefalica iar mai apoi in tot corpul. Prin contractia diafragmelor si a muschilor abdominali sangele circula in sens ascendent si antero-posterior. Pot exista si organe pulsatorii anexe ampule (antenale, pedale).
Sangele are rol trofic, de excretie, imunitar si endocrin.
Sistemul respirator al insectelor este un sistem foarte complex ce asigura schimbul de gaze dintre organism si mediu si realizeaza transportul oxigenului pana la nivelul tesuturilor unde se realizeaza respiratia tisulara.
Respiratia insectelor este de tip trahean, doar in cazul larvelor acvatice ea este de tipul traheo-branhial si uneori se poate realiza cutanat (tegumentar).
Sistemul trahean este alcatuit dintr-un mare numar de trahei (tuburi de origine ectodermica) ramificate si anastomozate. El comunica cu exteriorul prin orificii pereche (stigme, spiracule) ce se deschid lateral pe mezo- si metatorace precum si pe segmentele abdominale. Numarul de stigme este foarte variabil (10 perechi-lepidoptera, 5-6 perechi diptera).
Stigmele au forma ovala sau rotunda fiind incadrate la exterior de un element chitinos (peritrem) continuat cu un spatiu (atrium) alcatuit din doua camarute ce au perisori pentru filtrare si un sistem de inchidere.
Traheele sunt tubusoare ce pornesc de la stigme si se ramifica in tot corpul. Tubul unei trahei este format din doua straturi: la exterior se afla matricea (hipoderma tegumentului) iar la interior intima.
Traheile ce pornesc din stigme formeaza trunchiuri traheale ce pot fi: longitudinale, laterale si mediane ce sunt legate prin trahei anastomozate. Rezulta astfel trei ramuri:
dorsala deserveste inima, musculatura si tegumentul;
ventrala deserveste lantul nervos ventral, musculatura si tegumentul;
viscerala deserveste organele din sinusul visceral.
Traheile ce pornesc din atriul stigmelor formeaza la randul lor cate trei
ramificatii: dorsala, ventrala si mijlocie. Ramurile lor se subdivizeaza dichotomic, in tuburi tot mai subtiri formand o vasta retea de traheide si traheole ce se termina cu celule traheene. Unele insecte prezinta dilatari ale tuburilor (saci cu aer rezervoare).
Sistemul nervos este de tip ganglionar, se bazeaza in principal pe un ganglion ventral (chord) mai dezvoltat, cu cate o pereche de ganglioni pentru fiecare segment, ce se aseamana cu o scara de franghie. La unele grupe de insecte aceasta dispunere este mai condensata.
Sistemul nervos este format din trei diviziuni: sistemul nervos central (al vietii de relatie), sistemul nervos simpatic (visceral), sistemul nervos periferic (senzorial).
Sistemul nervos central principala diviziune, este alcatuit din:
Ganglionii cerebroizi (supraesofagieni - creierul sau cerebrum); se afla in capsula cefalica si sunt reprezentati de trei perechi de ganglioni:
Protocerebrum este mai mare si inerveaza ochii compusi si functioneaza ca un centru pentru asocieri.
Deutocerebrum este mai mic si inerveaza antenele, echipate cu numeroase sensillae.
Tritocerebrum formeaza o comisura ce se continua pe sub intestin (inerveaza fata si labrum). Creierul controleaza activitatea tuturor ganglionilor din corp.
"Creierul" (ganglionul cerebral) este alcatuit dintr-o portiune largita supra = ganglionul epiesofagian care este conectat prin conexiuni cu ganglionul subesofagian, care s-a format prin fuziunea ganglionilor pieselor bucale.
Masa nervoasa subesofagiana este situata tot in capsula cefalica, sub esofag, coordoneaza organele bucale si glandele salivare, are trei portiuni: mandibulara, maxilara si labiala;
Catena ganglionara scalariforma toraco-abdominala este formata de regula din 3 perechi de ganglioni toracici si 8 perechi de ganglioni abdominali. Perechile de ganglioni ale segmentelor pro-, mezo- si metatoracice inerveaza cele trei perechi de picioare si - daca sunt prezente - muschii aripilor iar ganglionii abdominali actioneaza asupra diferitelor organe interne. Ganglionii dintr-un segment se leaga cu cei din segmentele invecinate prin fibre nervoase longitudinale (conective). Ganglionii aceluiasi segment se leaga intre ei prin fibre transversale (comisuri).
De la ganglioni pornesc spre tot corpul nervii ce pot fi senzitivi si motori.
Sistemul nervos simpatic visceral (vegetativ) consista din trei regiuni:
Portiunea stomatogastrica (legat de tritocerebrum prin nervul recurent) ce inerveaza mezenteronul, inima si alte organe un nerv simpatic singular ventral ce inerveaza stigmatele, traheile si partea ventrala a corpului: si un nerv simpatic caudal ce inerveaza partea posterioara este responsabil de intestin si de gonade.
Sistemul nervos periferic are urmatoarea alcatuire:
cordoane nervoase senzitive (leaga receptorii din tegument de ganglioni primesc excitatia din exterior);
cordoane nervoase motoare (leaga ganglionii, prin nervul motor, de muschi transmitand raspunsul si inchizand astfel arcul reflex prin care se realizeaza legatura cu mediul).
Sistemul excretor este reprezentat de catre tuburile Malpighi care sunt de origine ectodermica si functioneaza ca principalul sistem excretor; anatomic, ele sunt apendici tubulare (ca un fund de sac) ale intestinului, fiziologic au functie de rinichi. Forma si numarul lor variaza intre 1-2 perechi (heteroptere, afaniptere, diptere) si 160 (ortoptere). Hemolimfa ce contine produsele de dejectie circula in hemocel in apropierea acestor structuri, numarul lor este specific fiecarei specii. Ele lipsesc la colembole si afide, excretia realizandu-se cu ajutorul glandelor labiale in primul caz si prin glandele rectale in al doilea caz. Principala functie a tuburilor Malpighi este aceea de absorbi acidul uric si uratii (ca saruri de sodiu si potasiu) si de a le deversa in lumenul intestinului, de unde produsele de excretie se amesteca cu fecalele. Uneori au rol de organe secretoare de enzime sau de organe luminoase (diptere). La unele insecte excretia este cutanee. La excretie mai participa corpul gras si celulele pericardice.
Corpul gras (tesutul gras), alcatuit din celule poliedrice sau rotunde, de natura mezodermica, se afla in cavitatea generala a corpului; este mai dezvoltat la larve si la stadiile hibernante si indeplineste functii de excretie (celule de urati, oenocite) si mai ales de acumulare pentru substantele nutritive de rezerva (globule uleioase, trofocite);
Celulele pericardice (nefrocite si fagocite) sunt perechi de celule dispuse in jurul vasului sanguin dorsal (in sinusul pericardial), au functii de excretie si de acumulare. Absorb substantele coloidale.
Sistemul secretor este reprezentat de glandele exocrine si cele endocrine.
Glandele exocrine au canale speciale prin care secreta la exteriorul organismului diferite substante. Ele au functii si origini diferite:
glandele sericigene situate in regiunea intestinului unor omizi, secreta fire de matase;
glandele cerifere secreta ceara cu rol protector pentru oua (lepidoptere), corp (afide), fiind situate in diferitele regiuni ale corpului;
glandele urticante ce secreta substante toxice folosite la aparare (larve de Lymantria, Hyphantria, Euproctis etc.);
glande atractante (pentru sexul opus - lepidoptere etc.);
glande repulsive (emana un miros neplacut, plosnitele).
Glandele endocrine au secretie interna, produsii lor fiind eliminati direct in sange. Hormonii produsi influenteaza metabolismul, cresterea, naparlirea si reproducerea. Ele pot fi:
glande neurosecretorii ale ganglionilor cerebroizi, secreta hormonul "creierului" ce dirijeaza cresterea si dezvoltarea prin glandele protoracale si "corpora cardiaca";
glandele protoracale secreta ecdysona (asigura naparlirea si dezvoltarea larvelor);
corpora allata dispusa posterior fata de ganglionii cerebroizi, secreta hormonul juvenil prin care sunt controlate numeroase procese (metamorfoza, histogeneza, histoliza, crestere si dezvoltare);
corpora cardiaca situata inainte de corpora allata cu care se leaga, fiind conectata si cu sistemul nervos central si cu vasul dorsal. Are un rol mai putin cunoscut.
Aparatul reproducator si reproducerea la majoritatea insectelor reproducerea este sexuata deci sexele sunt separate. Aparatul reproducator este alcatuit din glande sexuale (gonade), caile genitale si organe anexe.
Aparatul genital mascul este format din testicule (ce produc spermatozoizii), canale deferente, canalul median ejaculator, glande anexe si penis (aedeagus - ce poate fi foarte sclerificat: sistematica genitaliilor). Gonoporul este in spatele segmentului 9 abdominal.
Aparatul genital femel este alcatuit din doua ovare, oviducte laterale, oviduct comun, vagin, vulva si glande anexe. Ovarele sunt formate dintr-un numar variabil de ovariole (4, 6 sau 8 pe ovar, mai mult de 100 Diptere si pana la 2400 la femelele de Termite). Acestea secreta ovulele (celulele sexuale femele). Gonoporul se afla pe segmentul 8 abdominal (camera genitala sub sternitul 8).
Oviparitatea este regula generala la insecte, ouale pot fi depuse intr-un stadiu mai mult sau mai putin embrionat, ce poate ajunge pana la ovoviviparitate (eclozarea larvelor chiar inainte de depunere sau in acelasi moment). Viviparitatea este rara si se intalneste numai la glossine si la Pupiare; la fel de rara este si partenogeneza.
O singura imperechere este regula pentru insecte si implica inseminarea interna si umplerea spermatecii. Atunci cand oul matur este depus, intregul sac ovarian se contracta dar adesea acesta ramane ca un corp rezidual, aspect ce poate indica varsta fiziologica a individului (de ajutor la analiza populatiilor de Glossina spp., Simulium spp. si Anopheles spp.). Cand ouale trec prin deschiderea spermatecii se produce fertilizarea iar forma finala a oului este reglata de catre excretiile glandelor accesorii. Unii spermatozoizi patrund total via micropil. In timp ce spermatozoizii trec prin "galbenus", nucleul femel se divide meiotic in patru nuclei haploizi. Trei dintre ei degenereaza iar cel ramas fuzioneaza cu nucleul spermatozoidului care ajunge primul. Aceasta duce la realizarea unui set de cromozomi diploizi al caror numar variaza cu specia si chiar rasa (ex., Culex pipiens are sase cromozomi (= 2 n). Acest ou demareaza embrionarea care la insectele parazite se produce superficial deoarece celulele se divid la inceput ca un strat la suprafata "galbenusului" central.
Ca la toate artropodele, datorita prezentei exoscheletului rigid cresterea nu este continua si nu se face decat cu ocazia naparlirilor cand se produce schimbarea pielii (exuvie). Cresterea se produce la stadiile de larva, adultul (imago) este un organism a carui crestere s-a terminat.
La insecte intalnim doua tipuri de dezvoltare: heterometabola si holometabola.
Metamorfoza heterometabola (hemimetabolie) este o metamorfoza incompleta deoarece lipseste stadiul de pupa, este caracteristica pterigotelor inferioare. Unii autori disting pentru dezvoltarea acestor insecte doua stadii: larva si nimfa; altii folosesc numai notiunea de nimfa deoarece in multe cazuri juvenilii seamana cu adultul. Denumirea de pupa este folosita numai pentru metamorfoza completa (holometabola).
Metamorfoza holometabola insecta trece prin patru stadii in timpul dezvoltarii: ou, larva, pupa si adult. Larvele lor se deosebesc de adulti ca morfologie externa, ca structura interna si sub aspect fiziologic. Numarul stadiilor larvare variaza foarte mult in functie de grup: 3 stadii larvare la muste si alte diptere ciclorafe, la purici si paduchi; 4 stadii la tantari si flebotomi; 5 stadii la plosnite; 6-8 la simulide; 7 si mai mult la tabanide etc.
Stadiul de larva reprezinta un stadiu activ al dezvoltarii insectelor. Se caracterizeaza prin hranirea intensiva si acumularea substantelor nutritive necesare ulterior. O serie de caractere morfologice comune permit gruparea larvelor in urmatoarele tipuri:
larve protopode corpul lor este profund afectat de viata parazitara, apendicele cefalice si cele toracice sunt rudimentare, sunt dezvoltate doar mandibulele, abdomenul incomplet segmentat;
larve polipode (sau eruciforme) pot fi terestre sau acvatice, segmentarea este evidenta, au picioare toracice si apendice abdominale (picioare false - pseudopode (pedes spurii) - omizile de lepidoptere 2-5 perechi, larvele viespilor 7-8 perechi); mai au apendice abdominale ce servesc ca branhii traheene capul bine dezvoltat, aparat bucal pentru rupt si masticat;
larve oligopode nu au picioare abdominale, au capul, aparatul bucal si picioarele toracice bine dezvoltate, uneori pe ultimul segment abdominal se afla un apendice locomotor (pygopodium);
larve postoligopode se aseamana cu adultii morfologic si biologic (toate insectele hemimetabole).
larve apode viermiforme, pot prezenta rudimente de picioare; dupa structura capului acestea pot fi:
o eucefale (capsula cefalica normala, aparat bucal pentru masticat unele lepidoptere, coleoptere);
o hemicefale (au capsula cefalica invaginata in torace, aparat bucal pentru supt - unele diptere-tipulide);
o acefale (lipsite de capsula cefalica, capul complet invaginat - unele diptere-brahicere);
Durata stadiului larvar variaza cu specia si conditiile ecologice de la 3-4 zile la cateva luni si ani (10-17).
Stadiul de pupa ultimul stadiu larvar se transforma in nimfa la insectele heterometabole sau pupa la cele holometabole. Unele larve isi confectioneaza inainte de impupare un invelis (cocon) din matase sau alte materiale.
Morfologic, pupele se impart in doua categorii:
pupa dectica au tecile aripilor si picioarele libere, mandibulele sclerificate si active (servesc la ruperea coconului pupal);
pupa adectica nu are mandibulele sclerificate; se deosebesc urmatoarele tipuri: pupa libera (apendicele corpului sunt libere si pot executa unele miscari - coleoptere, himenoptere, diptere); pupa obtecta - mumie (apendicele lipite de corp, acoperite de o membrana); pupa coarctata, este o pupa libera in interiorul unui puparium.
Pupa este stadiul imobil din dezvoltarea holometabolelor (nu se hraneste si nu seamana nici cu larva nici cu adultul), acum au loc o serie de procese interne complexe ce se finalizeaza cu emerjarea insectei adulte (imago).
Daca luam in considerare exclusiv grupele parazitice, este acceptata urmatoarea clasificare sumara:
Subclasa: Apterygota (formele fara aripi)
Subclasa: Pterygota (primar cu aripi ce pot fi reduse la unele grupe)
Ordinul: Phthiraptera (paduchii animali)
Subordinul: Anoplura (paduchii hematofagi)
Subordinul: Mallophaga (paduchii rozatori)
Ordinul: Rhynchota (Hemiptera, plosnite)
Familia: Reduviidae (plosnite pradatoare)
Familia: Cimicidae (plosnita de pat)
Ordinul: Diptera (aripile anterioare dezvoltate, cele posterioare = haltere )
Familia: Phlebotomidae (flebotomi)
Familia: Culicidae tantari)
Familia: Ceratopogonidae (ceratopogonide)
Familia: Simuliidae (simulide)
Subordinul: Brachycera
Familia: Tabanidae (tauni)
Subordinul: Cyclorrhapha
Familia: Muscidae (muste)
Familia: Calliphoridae (muste)
Familia: Faniidae (muste)
Familia: Sarcophagidae (muste)
Familia: Glossinidae (musca tsetse)
Familia: Oestridae (muste)
Familia: Gasterophilidae (muste)
Familia: Hypodermatidae (muste)
Familia: Hippoboscidae (muste)
Ordinul: Siphonaptera (Aphaniptera, purici
Ordinul: Dictyoptera (Blattodea, gandacii de bucatarie)
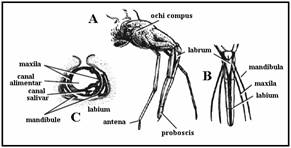
PLANSA 7.
Generalitati insecte (femela de lacusta)
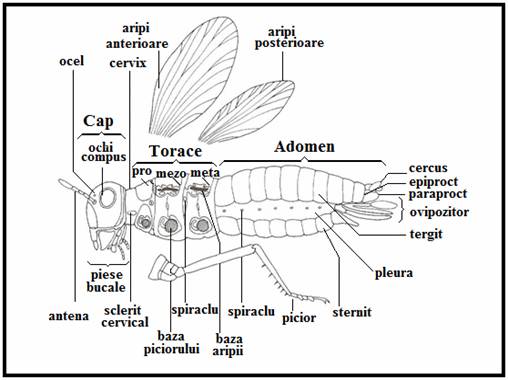
(adaptare dupa Wiliam S. Romoser & John G. Stoffolano, 1994)
Morfologia externa
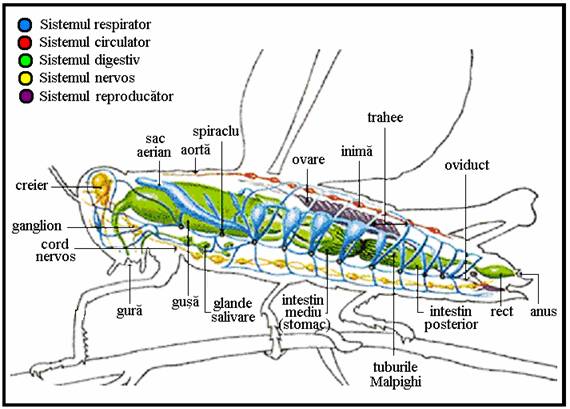
Anatomia interna
PLANSA 8.
Reprezentarea diagramatica a cuticulei tipice de insecte
si membranele peritrofice (MP)
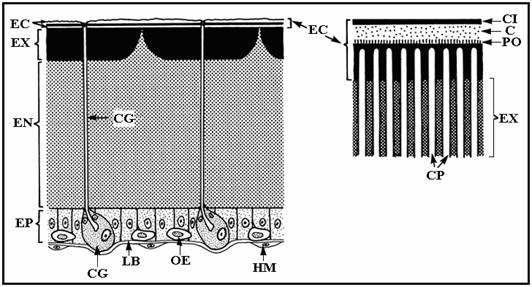
LB, lamina bazala; CI, stratul de cement; EC, epicuticula; EN, endocuticula; EP, epiderma; EX, exocuticula; CG, celula glandulara; CG, canal glandular; HM, hemocit; OE, oenocit; CP, canal porifer; PO, stratul de polifenol; C, stratul de ceara.
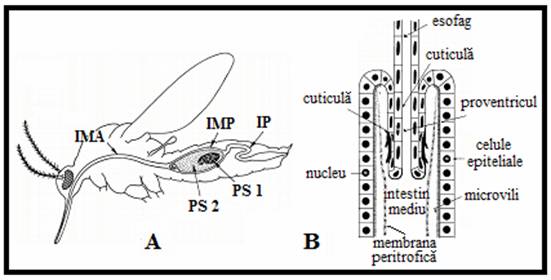
(dupa Peters 1976).
A. In femelele adulte de tantari diferitele parti ale membranei peritrofice sunt formate in doua regiuni ale intestinului mediu. O mica portiune este secretata de catre celulele regiunii anterioare (IMA) si este transportata catre jonctiunea dintre intestinul mediu anterior si cel posterior, totusi cea mai mare parte a membranei este produsa de catre regiunea posterioara a intestinului mediu (IMP) pentru a inveli sangele ingerat (PS). (PS1, PS2, primul si al doilea pranz sanguin; IP, intestinul posterior.
B. Reprezentarea cardia, pentru acele insecte la care numai o zona redusa de celule specializate situata la debutul intestinului mediu produce MP.
PLANSA 9.
Diferite tipuri de aparate bucale
Tipul masticator - unele larve Tipul taietor, absorbant sau lins muste

tantari Tipul pentru intepat si supt purici
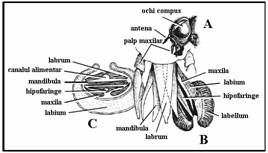
Tipul taietor, absorbant sau lins Tipul pentru intepat si supt
tabanide plosnite
Ordinul Diptera (mustele adevarate: muste, musculite, tantari)
Numele deriva din "di" - doua, "ptera" aripi, cuprinde:
Ordinul Diptera cuprinde peste 125.000 de specii grupate in peste 120 de familii. Ca numar de specii se plaseaza pe locul patru dupa Coleoptera (350.000 de specii), Lepidoptera (150.000 de specii) si Hymenoptera (100.000 de specii). Dipterele ocupa insa primul loc din punctul de vedere a medicinii umane si al celei veterinare, fie datorita rolului lor vector pentru diferite organisme patogene fie disconfortului pe care il genereaza.
Sunt insecte care in stadiul de adult au numai o pereche de aripi. Larvele lor sunt apode.
Alternanta pe parcursul ciclului de viata a doua "morfologii" si "biologii" diferite cea larvara si cea adulta este legata de dezvoltarea holometabola (metamorfoza completa).
De regula, sunt insecte ectoparazite, dar exista si specii de diptere endoparazite. Desi prezinta o serie de trasaturi comune, organizarea de baza a corpului lor este modificata conform diferitelor lor modalitati de viata.
Morfologie si dezvoltare
Adultii dipterelor au corpul net divizat in trei parti: cap, torace si abdomen.
Caracteristice ordinului sunt antenele si piesele bucale. Antenele pot fi fie filiforme (Nematocera), fie scurte, alcatuite din trei articule (Brachycera). Ochii sunt in general mari si compusi din numeroase omatidii. Aparatul bucal este adaptat pentru absorbit, taiat si absorbit, pentru intepat si supt (la hematofage) sau de tip masticator (Plansa 11).
Tipul masticator este intalnit la larve si este comun multor specii ce traiesc liber sau sunt parazite.
Tipul absorbant este intalnit la majoritatea dipterelor neintepatoare (Musca, Calliphora).
Tipul taietor, absorbant este caracteristic tabanidelor si glosinelor.
Tipul pentru intepat si supt este prezent la o serie de ectoparaziti hematofagi (tantari, musca tsetse, alte muste). Marimea si forma pieselor bucale cu rol in strapungerea pielii, este legata de cele doua modalitati de hranire cu sange telmofagie si solenofagie.
Cele trei segmente ale toracelui sunt inegal dezvoltate. Cel de al doilea segment mezotoracele este cel mai bine dezvoltat deoarece poarta singura pereche de aripi. Aripile posterioare sunt reduse la o pereche de structuri numite haltere. Functia lor este cea de organ de echilibru in timpul zborului si mai contribuie si la orientarea spatiala (datorita grupului de senzori ce detecteaza stresul mecanic impus de vibratiile halterelor).
Adultii parasesc invelisul nimfal printr-o fanta rectilinie - diptere ortorafe sau printr-o deschidere circulara - diptere ciclorafe in acest caz ele poseda la cap un organ special - ptilinum. Adultii sunt entomofagi, fitofagi, floricoli, hematofagi.
Larvele se deosebesc unele de altele prin gradul de dezvoltare al capsulei cefalice. Larvele eucefale au o capsula cefalica bine dezvoltata ce individualizeaza capul de restul corpului. Larvele hemicefale au o capsula cefalica incompleta, ascunsa aproape total in torace. Larvele acefale sunt lipsite de capsula cefalica, prezentand crosete bucale ce semnalizeaza zona anterioara a corpului. Numarul de stadii larvare difera de la un grup la altul. "Mustele" si dipterele muscoide au intotdeauna trei stadii, majoritatea hematofagelor au patru stadii, unele familii de Simuliidae si Tabanidae au 6-7 stadii. Deoarece au nevoie de multa umiditate le intalnim in zonele umede sau in apa. Ca regim alimentar larvele pot fi: carnivore, detritofage, parazite etc.
Nimfele adesea imobile pot fi incluse uneori intr-un puparium sau pot fi libere.
Ciclul de viata (Plansa 10) corespunde dezvoltarii holometabole ce include larvele apode (fara picioare), o pupa ce nu se hraneste si adultii bisexuati. Larvele si adultii pot trai ca paraziti; larvele in general ca endoparaziti iar adultii ca ectoparaziti. Cu exceptia catorva specii (Hipoboscidae), dipterele adulte se hranesc periodic pe diferitele lor gazde. Aproximativ 10% dintre speciile de diptere sunt acvatice in stadiul larvar. Ouale si pupele acestor specii sunt de asemenea acvatice, adultii insa sunt terestri. Majoritatea dipterelor sunt ovipare, unele depun larve (Glossinidae) altele depun pupe (Hipoboscidae, Streblidae, Nycterbidae), foarte putine se reproduc in stadiul de larva prin pedogeneza (Cecidomyidae).
Clasificarea
De regula, clasificarea se stabileste dupa caracterele adultilor. Lungimea antenelor este primul caracter utilizat. Apoi urmeaza fanta ptilinala ce indica modul de iesire din pupariu. Dipterele se impart in doua subordine Nematocera si Brachycera. Acestea pot fi deosebite morfologic prin venatiunea aripilor, prin tipurile de antene ale adultilor si prin habitatele ecologice (Tabel 1).
Subordinul Nematocera include cateva specii importante din punct de vedere medical si veterinar. Tantarii, simulidele (Simulium sp.) si ceratopogonidele sunt incluse in acest subordin. Majoritatea adultilor au antene compuse din numeroase segmente alungite, antene filamentoase. Majoritatea mustelor din acest subordin au corpul de dimensiuni reduse (cativa milimetri lungime) si pot fi clasate ca avand o structura delicata. Adultii parasesc exuvia nimfala printr-o fanta rectilinie situata dorsal cefalotoracelui. Familiile mai importante sunt: Phlebotomidae, Psychodidae, Culicidae, Ceratopogonidae, Simuliidae, Chironomidae, Tipulidae, Bibionidae, Anisopodidae, Mycetophilidae, Cecidomyidae etc.
Subordinul Brachycera sunt caracterizate de antenele alcatuite din trei segmente (cel terminal fiind de tipul annulat). Asta inseamna ca pe el se afla constrictiuni circulare ce dau aparenta mai multor segmente separate.
Aceste muste au corpul mai mare, 5-20 mm si mai robust. Din acest subordin fac parte tabanidele si celelalte muste. Adultii se diferentiaza prin prezenta aristei (arista) de pe segmentul terminal al antenei. La abdomenul formelor superioare de Brachycera rareori este vizibil segmentul sapte. Aripile au o venatiune mai redusa decat la Nematocera si decat la formele inferioare de Brachycera.
Toate Dipterele sunt holometabole, au o metamorfoza completa: ou - larva - pupa adult. De regula au patru stadii larvare, totusi la unele sunt vizibile numai trei (Cyclorrhapha), cel de al patrulea se dezvolta in exuvium (cuticula durificata) a celui de al treilea stadiu larvar. Exuviumul durificat este, de regula, inchis la culoare si poarta numele de puparium. Majoritatea Nematocera si Brachycera au larve acvatice, la unele Cyclorrhapha pupariumul se dezvolta in sol.
Tabel 1. Comparatie sintetica intre cele doua subordine.
|
Sub-ordinul NEMATOCERA |
Sub-ordinul BRACHYCERA |
|
Antene multi-articulate (+ de 10 articole) mai lungi sau de aceiasi lungime cu capul + toracele |
Antenele reduse si aristate |
|
Cap prognat |
Cap ortognat |
|
Palpi maxilari lungi (4-6 articole) |
Palpi maxilari scurti (1-2 articole) |
|
Larve eucefale |
Larve acefale |
|
Nimfe libere, mobile sau foarte mobile |
Nimfe cel mai ades inchise in ultima exsuvie larvara (pupa) |
In cele ce urmeaza vor fi prezentate principalele familii din ordinul Diptera, cu importanta pentru Entomologia medicala (Tabel 2).
Mustele intepatoare - in majoritatea cazurilor numai femelele adulte se hranesc cu sange (sunt hematofage).
Culicidae (mosquitoes - tantarii) - pot raspandi malaria, encefalite, febra galbena, filarioze si alte boli.
Tabanidae (horse flies / deer flies - tabanidele) - pot raspandi tularemia, loaiaza, trypanosomiaza si alte boli.
Simuliidae (black flies - simulidele) pot raspandi oncocercoza umana si infectiile cu leucocytozoon la pasarile domestice.
Psychodidae (moth flies - flebotomii) pot raspandi leishmanioze, febra de trei zile si alte boli.
Ceratopogonidae (punkies, no-see-ums - ceratopogonidele) - mici insecte intepatoare de care se leaga raspandirea unor patogeni ca viermi, protozoare si virusuri la oameni si la alte animale.
Muscidae (House flies - musca de casa) - sunt printre cele mai cosmopolite dintre toate insectele. Unele specii au aparat bucal de tip intepator, altele de tipul masticator. Boli ca dizenteria, holera si miazele pot fi transmise de piesele lor bucale si de alte parti ale corpului lor.
Mustele parazite - larvele sunt paraziti sau parazitoizi ai altor animale.
Oestridae (bot flies / warble flies) - larvele sunt endoparazite ale mamiferelor, inclusiv la om.
Hippoboscidae (louse flies - musca paduche) - adultii sunt hematofagi, ectoparaziti ai pasarilor si mamiferelor.
Tabel 2. Dipterele mai importante si unii dintre agentii patogeni transmisi.
|
Familie / Gen |
Bolile umane (patogeni) |
Bolile animalelor domestice (patogeni) |
|
Subordinul Nematocera | ||
Culicidae | ||
Aedes |
febra galbena (V), febrele tip Denga (V), filarioza (N) |
mixomatoza la iepuri (V) |
Culex |
encefalita St.Louis (V),filarioza (N) |
encefalita equina (N), malaria la gaini (P), filarioza canina (N), |
Anopheles |
malaria (P),filarioze, elefantiazis (N) |
filarioze (N) |
|
Mansonia (Coquilettidia) |
filarioze (N) |
filarioze (N) |
|
Simuliidae | ||
|
Simulium |
oncocercoza (N) |
malaria Leucozyozoon la pasari |
|
Phlebotomidae | ||
|
Phlebotomus, |
bartoneloza (R,B), | |
|
Lutzomyia |
febra Papataci (V), leismanioza (P) |
leismanioza canina (P) |
|
Subordinul Brachycera | ||
|
Tabanidae | ||
|
Chrysops |
tularemie (B), loaiaza (N) |
Surra (P) |
|
Tabanus |
loaiaza (N) ? |
anaplasmoza (R) |
|
Haematopota |
filarioze (N) |
|
Muscidae | ||
Musca |
poliomelite (V), shigielloza (B), salmonelloze (B), holera (B), trachoma (V), ameebiaze (P), miaze produse de larve |
habronemiaza ecvida (N), thelaziaza (N) |
|
Stomoxys |
poliomelite (V), bacterioze (B) |
boala somnului (P), spirochetoza gainilor (S), habronemiaza |
|
Subordinul Cyclorrhapha | ||
|
Glossinidae | ||
|
Glossina |
boala somnului (P) |
Nagana, Surra (P) |
|
Sarcophagidae | ||
|
Sarcophaga |
miaze produse de larve |
miaze produse de larve |
|
Wohlfartia |
miaze produse de larve |
miaze produse de larve |
|
Calliphoridae | ||
|
Callitroga |
miaze produse de larve |
miaze produse de larve |
|
Gasterophilidae | ||
Gasterophilus |
miaze produse de larve |
miaze produse de larve |
|
Oestridae | ||
|
Oestrus |
miaze produse de larve |
miaze produse de larve |
|
Hypoderma |
miaze produse de larve |
miaze produse de larve |
|
Dermatobia |
miaze produse de larve |
miaze produse de larve |
Legenda: V=virusuri; P= protozoare; N=nematode; B=bacterii; R=ricketssii.
PLANSA 10.
Stadiile de dezvoltare ale ciclului
holometabolic de la unele
grupe importante de Diptera
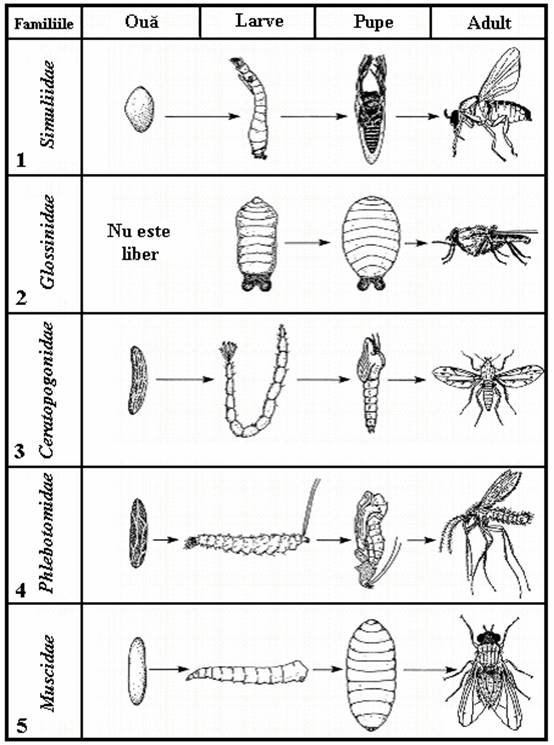
De retinut, ca pot exista mai multe stadii larvare, temperatura si alti factori externi le regleaza viteza de dezvoltare.
PLANSA 11.
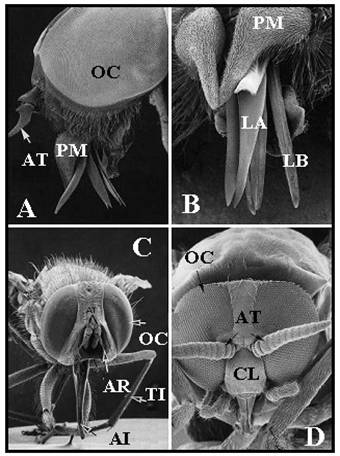
Imagini electrono-microscopice ale capului unor diptere hematofage
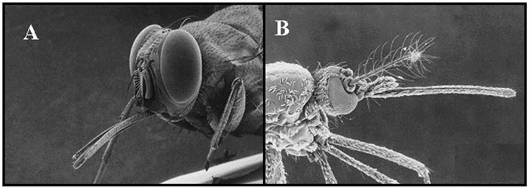
Imagini electrono-microscopice ale pieselor bucale ale unor diptere hematofage. Ambele isi pastreaza stiletii intepatori intr-un invelis retractil. A. Musca tsetse Glossina morsitans × 50. B. Tantarul Aedes aegypti care transmite febra galbena
(Sursa Internet)
Imagini electrono-microscopice ale capului unor diptere hematofage. A/B. Tabanus sp.; aspect lateral si piesele taietoare marite (A × 40, B × 60). C. Stomoxys calcitrans (× 40). D. Simulium damnosum, cap de femela (× 80). AR, arista; AT, antena; CL , clypeus; OC, ochi compusi; LA, labrum; LB, labium; PM, palp maxilar; AI, aparatul intepator; TI, tibia.

Familia cea mai importanta sub aspect medical dintre Nematocere (insecte cu antene filiforme) este Culcidae.
Tantarii au o vechime de peste 50 de milioane de ani, cu mult inainte de adaptarea ulterioara la oameni. Toate populatiile umane sunt afectate de catre tantari, in principal prin intepaturi si prin transmisia bolilor. Au fost identificate peste 3500 de specii, genurile cele mai importante Anopheles Culex Mansonia si tribul Aedes fac parte din subfamiliile Anophelinae, Culicinae si Toxorhynchitinae)
Tantarii au fost primele insecte la care a fost observat un agent patogen al unei boli, filarioza Bancroftiana (1877). Cunostintele ulterioare au demonstrat ca ei sunt vectori pentru agenti patogeni ce fac parte din mai multe grupe taxonomice: virusuri, bacterii, protozoare si helminti, desi in principal sunt recunoscuti ca vectori ai malariei.
Tantarii se intalnesc aproape peste tot in lume, in majoritatea tipurilor de zone ecologice, lipsind doar din Antarctica si din cateva insule.
Sunt insecte holometabole, larvele si pupele traiesc in apa iar adultii in mediul terestru. La acest grup de diptere, numai femelele sunt hematofage. Adultii se disting de alte Nematocere ne-hematofage (ex. Chironomide) prin solzii de pe nervurile aripilor si prin proboscisul lung, indreptat anterior.
Morfologie si ciclul de dezvoltare (Plansa 12, 13
Adultii tantarii adulti au in general intre 5-20 mm lungime.
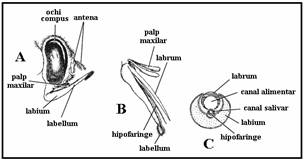
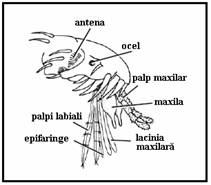
Capul globulos, prezinta doi ochi mari compusi (fara oceli) si antene filamentoase lungi (15 articule masculul, 16 articule femela). Articulele flagelului au la baza verticile de peri a caror lungime si numar variaza cu sexul (lungi si numeroase la mascul - antene plumoase). Acestea poarta organele de simt cu ajutorul carora recunosc gazdele si locurile pentru ovipozitie precum si organul lui Johnston cu ajutorul caruia masculii recunosc bataile aripilor femelelor. Piesele bucale de tip intepator sunt proeminente si au o lungime egala cu ce a regiunii cap/torace fiind formate din labium ce inveleste stiletii care s-au dezvoltat din labrum (ce formeaza canalul alimentar), cele doua lacinii mandibulare si hipofaringele nepereche contin canalul salivar. Ansamblul lor formeaza trompa. Lungimea, forma si pilozitatea palpilor maxilari alcatuiti din cinci segmente difera cu specia si sexul, fiind reduse la masculi deoarece nu se hranesc cu sange, ci cu substante zaharoase si nectar.
Toracele este globulos, alcatuit din trei segmente sudate si poarta dorsal o singura pereche de aripi. Solzi de forme, culori si cu dispunere variata acopera aripile, segmentele toracice si picioarele.
Abdomenul este format din 10 segmente (8 vizibile la exterior) este ornat cu solzi si peri (solzii lipsesc la Anopheles). Ultimele segmente abdominale constituie genitalia; cele ale masculilor fiind mult utilizate in sistematica. Masculii si femelele pot fi usor separati cu ajutorul antenelor, care sunt mai paroase la masculi si cu ajutorul pieselor bucale care la masculi sunt mai slab dezvoltate. Dupa emerjare, genitaliile masculului se rotesc cu 180°, pentru a usura copulatia in timpul zborului.
Adultii genului Anopheles au, in pozitie de repaus, toate partile corpului (proboscisul, capul, toracele, abdomenul) dispuse in linie dreapta, formand cu suprafata un unghi de 30° 45°. Venele aripilor sunt acoperite in mod caracteristic cu solzi de culoare deschisa sau inchisa. De regula, acestia lipsesc total de pe sternitele abdominale. Ambele sexe au palpi lungi si negri. La adultii aflati in repaus, corpul este aproape paralel cu suprafata. Sternitele si tergitele sunt acoperite cu solzi densi iar palpii femelelor au o treime din lungimea proboscisului.
Longevitatea adultilor variaza puternic in functie de regiunea climatica, in medie doua saptamani la tropice si patru sau cinci saptamani in regiunile temperate dar si cateva luni pentru femelele speciilor ce hiberneaza sau estiveaza.
Larvele se intalnesc patru stadii ca larve, cu o morfologie comparabila, diferentele fiind date in principal de talie. Ele sunt alungite, au o capsula cefalica bine sclerotizata, ce sustine mandibulele puternic sclerotizate, maxilele si periile bucale, ce ajuta la razuirea suprafetelor vegetale pentru desprinderea particulelor de hrana. Pe segmentele 8/9 fuzionate sunt situate o pereche de spiracule ce se deschid la suprafata la Anophelinae sau la capatul unui sifon sclerotizat la Culicinae. La toate speciile ultimul segment are o regiune sclerotizata ce poarta o perie ventrala utilizata ca o carma, la inot.
Normal, dezvoltarea embrionara este completata la cateva ore de la depunerea pontelor concretizandu-se cu aparitia larvelor de stadiul I. In cazul genului Aedes larvele complet dezvoltate raman in invelisul oului pana ouale sunt inundate si pot ramane asa pentru lungi perioade de timp (in functie de temperatura si umiditate chiar 4-5 ani). Larvele sunt acvatice si apar in principal in apele dulci, unele specii se pot dezvolta si in ape sarate. Marimea habitatului poate fi foarte mica, ex. scorburile, baza petiolului frunzelor, diferite recipiente artificiale. Durata totala a celor patru stadii larvale variaza mult, chiar si in cazul aceleiasi specii, depinzand in special de temperatura si cantitatea de hrana. La tropice ea poate fi completata intr-o saptamana, in regiunile temperate cateva luni sau chiar mai mult in cazurile in care exista diapauza. Unele specii sunt tolerante la frig altele traiesc la 50°C. Larvele se hranesc cu detritus sau cu plancton (filtratorii) sau sunt pradatorii altor larve. Unele larve sunt filtratori de suprafata (fara sifon) si atunci cand nu sunt deranjate ele raman paralele cu suprafata apei. In contrast cu anofelinele, culicinele pot fi descrise astfel: larvele stau intr-un unghi de circa 40° fata de suprafata apei sau fixate de plantele acvatice (Mansonia
Nimfele dezvoltarea pupelor este de asemenea acvatica si temperaturo-dependenta, durand intre o zi si trei saptamani. Ele au forma de virgula si prezinta pe cefalotorace o pereche de trompete respiratoare cu care respira la suprafata apei. Abdomenul recurbat alcatuit din 8 segmente se termina cu doua palete inotatoare dispuse la capatul posterior.
Ponta ouale sunt depuse, de obicei, pe suprafata apei si plutesc datorita tensiunii superficiale sau datorita prezentei flotorilor laterali sau apicali.
Cateva criterii ne permit sa facem distinctia dintre Anophelinae si Culicinae (Plansa): ouale de Anophelinae au forma de barcuta si sunt depuse cate unul, plutind la suprafata apei cu ajutorul unor flotori. In cazul culicinelor, ouale celor trei genuri se pot distinge astfel: ouale negre de Aedes sunt depuse cate unul, cele de Culex sunt grupate, iar cele de Mansonia sunt lipite pe plantele submerse.
Reproducerea
In natura, de la cateva ore la o zi de la emergenta, adultii sunt gata pentru imperecherea ce se produce in timpul zborului. Masculii roiesc in zone speciale, acuplandu-se cu femelele ce trec pe acolo. La unele specii, masculii isi introduc secretiile glandelor accesorii in timpul copulatiei. Majoritatea femelelor trebuie sa se hraneasca cu sange pentru dezvoltarea oualor (anautogenie) deoarece zaharurile, ingerate de catre masculi si femele, sunt consumate in principal pentru zbor. Dezvoltarea oualor este indusa prin distensia intestinului mediu. Uneori primul ciclu ovarian poate fi completat fara hranirea cu sange (autogenie). Numarul de cicluri ovariene (depunerea pontelor), numarul de hraniri cu sange, precum si riscul transmisiei parazitilor este indicat de catre schimbarile de la nivelul ovariolelor. Fiecare femela are doua ovare si fiecare dintre acestea 50-200 de ovariole. In fiecare ciclu ovarian se dezvolta un singur ou/ovariola. In natura se produc, de regula, patru sau cinci ovipozitii, fiecare dintre ele cu 30-500 de oua. Locurile pentru ovipozitie sunt specifice fiecarei specii si sunt alese in functie de chimismul apei si de ritmul circadian. Inmultirea tantarilor in laborator este posibila pentru cateva specii, fara mari dificultati.
Cautarea gazdelor (Figura 1) de catre femelele de tantar este numai unul dintre aspectele comportamentale ca raspandirea, selectia microhabitatului, evitarea pradatorilor, bautul apei, hranirea cu zaharuri, imperecherea, copulatia si ovipozitia. Daca tantarii cauta sau nu o gazda si cat de intens se exprima comportamentul lor de cautare, depinde de starea lor fiziologica. La unele specii este necesara o mare cantitate de sange pentru producerea oualor, dar daca nu este o urgenta legata de hranirea cu sange, comportamentul de cautare a gazdei poate fi inhibat. Aceasta poate fi o adaptare la rata mare a mortalitatii tantarilor, cauzata de comportamentul defensiv al gazdei. La speciile cu capacitate intestinala mare comportamentul de cautare este inhibat dupa hranirea cu sange, datorita activarii receptorilor de presiune abdominali.
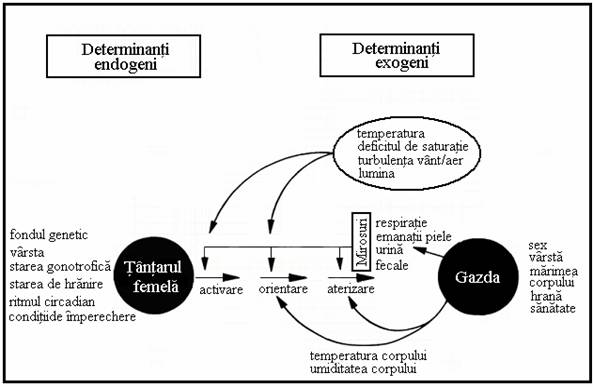
(dupa Takken 1996)
Figura 1. Diagrama factorilor endogeni si exogeni ce afecteaza localizarea gazdelor de catre femelele de tantari.
Dupa digerarea sangelui distensia abdominala se reduce iar cautarea unei noi gazde este inhibata de catre ouale in dezvoltare. Aceasta inhibare este rezultatul interactiunii complexe dintre ovare, corpul gras, celulele neurosecretoare si substantele provenite de la masculi in timpul fecundarii. In timpul fazei de graviditate pot fi, de asemenea, inhibati neuronii chemosenitivi antenali care raspund la semnalele gazdei. Alti factori fiziologici ca varsta, starea de nutritie, conditiile de imperechere, ritmul circadian, si numarul de cicluri gonotrofice completate pot modifica comportamentul de cautare al gazdelor.
Diferitele specii de tantari pot folosi diferite comportamente de cautare a gazdelor; ele pot sa prefere diferite tipuri de gazda iar pe gazde selecteaza diferite zone pentru intepare. Chiar si in cazul aceleiasi specii, pot sa apara mari variatii geografice privitoare la preferintele fata de gazda, aceasta selectie este determinata genetic. Cautarea gazdelor poate fi divizata in mai multe faze succesive: activarea, orientarea zborului catre gazda, asezarea pe gazda, tatonarea, imbibarea, retragerea si parasirea locului de hranire. Dintre factorii ce stimuleaza alegerea gazdei, orientarea zborului este cel mai bine studiat.
Orientarea zborului. Tantarii isi localizeaza gazdele prin anemotaxie, ei zboara dupa "dara" lasata de emanatiile gazdei. Miscarile lor sunt controlate de catre raspunsurile optomotoare la miscarile aparente ale suportului. Formele oportuniste ce se hranesc pe un spectru larg de gazde par sa fie atrase in principal de aerul exhalat, ce are dioxidul de carbon drept component stimulator, pe cand speciile cu o specificitate ridicata par sa raspunda mai ales la emanatiile particulare ale pielii. Orientarea pe distante lungi (peste 70 m) se realizeaza cu ajutorul mirosului pe distante scurte (1-2 m) temperatura corpului si umiditatea sunt factori suplimentari de atractie. Dioxidul de carbon in concentratii fluctuante nu actioneaza numai ca un kairomon in sine ci poate modula si efectele altor mirosuri ale gazdei. Unele mirosuri sunt mult mai atractive in prezenta dioxidului de carbon. Acesti stimuli par sa fie integrati cu stimulul reprezentat de dioxidul de carbon (si de asemenea, cu efectele umiditatii) la nivelul nervos central si sunt o modificare a raspunsurilor electrice a receptorilor. S-a constatat ca unele substante chimice atrag tantarii in conditii artificiale sau in anumite concentratii. Totusi, se cunosc putine lucruri despre natura chimica a atractantilor reali ai gazdelor. Acidul L-lactic in combinatie cu emanatii mirositoare ale pielii atrag pe Aedes aegypti iar 1-octen-3-olul combinat cu dioxidul de carbon atrage unele specii de tantari zoofili. Anopheles gambiae - antropofil, arata o preferinta ridicata pentru mirosul picioarelor.
Apropierea tantarilor de gazda mai implica stimulii vizuali, temperatura corpului si umiditatea asociata curentilor de convectie ce se ridica de la suprafata corpului gazdei. Alte substante chimice precum amino acizii pot de asemenea sa aiba efect. Raspunsurile fata de stimulii vizuali difera la diferitele specii ale aceluiasi gen. Se pare ca majoritatea speciilor prefera culorile inchise cu reflectivitate scazuta.
Tatonarea este stimulata de gradientii termali, umiditate, dioxidul de carbon, calitatea mecanica a suprafetei si unele substante chimice ca acizii grasi cu catena scurta.
S-a constatat ca ingestia sangelui este declansata de catre plachetetele sanguine si de catre diferite nucleotide de adenina in combinatie cu conditiile osmotice isotonice din sange (la specii de Aedes si Culex). Mecanisme complexe sunt responsabile de ajungerea sangelui in intestinul mediu. In final, terminarea hranirii este controlata de catre receptorii de presiune din abdomen.
Comportamentul de hranire si transmisia bolilor
Tantarii apar de regula in apropierea locurilor de emergenta. In functie de distanta dintre zonele de reproducere si zonele in care se gasesc gazdele pentru hranire deplasarea prin zbor poate acoperi cativa (zeci) km.
Hranirea cu sange urmeaza ritmul circadian specific fiecarei specii, in principal nocturn. Tantarii se hranesc cu sangele de la nivelul vaselor capilare terminand hranirea (4-10 μl) in cateva minute. Saliva lor contine o serie de compusi pentru a spori fluxul de sange, pentru a bloca coagularea, pentru anestezia locala si pentru eliminarea unor enzime ce faciliteaza localizarea vaselor de sange. Pompele cibariala si faringiana transporta sangele direct prin esofag la intestinul mediu. Lichidele zaharoase sunt directionate mai intai spre gusa si apoi spre intestin.
Interactiunea dintre Vector si Parazit
Dupa ingestia sangelui, arbovirusurile se multiplica in celulele intestinului mediu sau il strapung si se multiplica in hemolimfa inainte sa invadeze glandele salivare.
Atunci cand este ingerat sange (ce contine eritrocite cu gametocitii masculi si femeli de Plasmodium), scaderea temperaturii si un factor de activare al gametocitilor produs de catre tantar induce dezvoltarea lor la micro- sau macrogameti. Dupa fertilizare, zigotul care rezulta isi schimba proprietatile de suprafata transformandu-se intr-un ookinet alungit indicat de catre sensibilitatea fata de proteazele din intestin. Ookinetul produce o chitinaza cu ajutorul careia digera tesuturile catre membrana peritrofica (MP), penetreaza peretii intestinului intra- sau extracelular si raman in apropierea laminei bazale a peretelui intestinal. Aici el se dezvolta intr-un oocist ce se sparge si da nastere la mii de sporozoiti. Sporozoitii sunt transportati prin corp de catre hemolimfa si ajung la glandele salivare. Recunoasterea si penetrarea celulelor glandelor salivare (ca si in cazul celulelor intestinale) pare sa fie reglata de catre interactiunile lectina/zaharuri. Numai sporozoitii din glandele salivare, nu si cei eliberati din oocist, pot infecta hepatocitele. Inducerea pasilor acestei dezvoltari pare sa varieze in diferitele sisteme parazit/vector si a fost obtinuta in investigatii asupra malariei non-umane.
Dupa ingestia sangelui cu microfilarii, acestea se inchisteaza, strapung peretele stomacului, migreaza catre muschii aripilor din torace, cresc si naparlesc de doua ori si migreaza catre zona mai moale a labiumului pieselor bucale pe care il perforeaza in timpul hranirii.
Efectele parazitului asupra vectorului difera in functie de boala transmisa. In cazul tantarilor infectati de catre virusuri, longevitatea si capacitatea de hibernare sunt mult reduse. La tantarii infectati cu Plasmodium-se schimba compozitia aminoacizilor din hemolimfa, se reduce durata de zbor, longevitatea si fertilitatea. Aceste efecte par sa apara numai la infectia unor anumite specii de tantari fiind provocate de anumite tipuri de Plasmodium. Sunt suspectate si modificari ale comportamentului de hranire si de orientare datorita distrugerii celulelor glandelor salivare. Concentratia apirazei se reduce afectand recunoasterea vaselor de sange iar tantarii infectati tatoneaza mai mult decat specimenele sanatoase. In infectiile cu filarii, o densitate ridicata de paraziti poate reduce capacitatea de zbor si longevitatea.
Culicidele reprezinta grupul de insecte cel mai important pentru sanatatea publica, prin rolul lor de vectori in transmisia si mentinerea unor boli pe suprafete extrem de intinse ale globului. Ele reprezinta, de asemenea, elemente importante de disconfort, mai ales in aglomerarile urbane, in zonele industriale si turistice. Ca vectori si insecte de disconfort, prezenta culicidelor are consecinte economico-sociale negative dintre cele mai importante. De aceea, se impune mentinerea populatiilor de culicide la nivele cat mai scazute de abundenta in anumite perioade si zone, astfel incat sa fie eliminate riscurile epidemiologice si de impact economic legate de prezenta lor. Dar controlul populatiilor de culicide nu trebuie sa perturbe echilibrul ecosistemelor in care se intervine. Din aceasta cauza, este necesara cunoasterea populatiilor de culicide ca elemente componente importante ale ecosistemelor terestre si acvatice, naturale si antropizate.
Culicidele - componente ale ecosistemelor acvatice si terestre
Culicidele sunt insecte cu metamorfoza completa care isi desfasoara obligatoriu o parte a ciclului lor biologic in mediul acvatic iar alta in mediul terestru, fiind astfel componente ale biocenozelor acvatice dar si ale celor terestre.
Relatiile lor biocenotice sunt rezultatul evolutiei si stabilizarii ecosistemelor din care fac parte si se prezinta de regula sub forma unor sisteme parazit-gazda si prada-pradator, ce sunt sincronizate cu ciclurile biologice ale celorlalte componente ale biocenozelor.
Larvele - componente ale ecosistemelor acvatice
Larvele de culicide au fost gasite in cele mai diferite zone geografice, intr-o mare varietate de habitate acvatice. Ele populeaza anumite zone din marile complexe acvatice ca rauri si lacuri, cat si zonele mlastinoase, baltile de inundatie si de alt tip si, in general, orice acumulare de apa mai mare sau mai mica, naturala sau artificiala. De regula, larvele prefera zonele cu apa stagnanta, de adancime mica, bogate in vegetatie acvatica si partial insorite (NICOLESCU 1983, 1986; SERVICE 1969, 1976; DAHL & WHITE 1978).
Larvele poseda numeroase adaptari fiziologice (pot fi eurihaline sau stenohaline, euriterme sau stenoterme etc.), adaptari ale ciclului biologic (la unele specii ele traverseaza sezonul rece), adaptari de comportament fata de mediul in care se afla (dispozitia grupata etc.). Aceasta adaptabilitate le permite sa se dezvolte pe de o parte in habitate putin favorabile uneori si specifice (ape poluate, balti temporare in care trebuie sa-si completeze repede dezvoltarea larvara, scorburi de copaci etc.), pe de alta parte permite multor specii sa traiasca intr-o mare gama de tipuri de habitate, in ape dulci sau salmastre, in zona litorala a marilor lacuri sau in mici depresiuni din sol, pe marginea paraielor repezi sau in balti, in ape tulburi sau foarte limpezi, in ape reci sau calde, cu sau fara vegetatie (BECKER 1989, DABROWSKA-PROT 1979). Atunci cand habitatele in care se dezvolta larvele de culicide sunt supuse unor mari fluctuatii ale factorilor abiotici, fauna de culicide este reprezentata prin specii euritope, policiclice, caracterizate printr-o mare plasticitate ecologica (BECKER 1989). Populatiile speciilor univoltine sunt de obicei stenotope, se dezvolta in habitate specifice iar fluctuatii mai insemnate ale factorilor de mediu le pot elimina foarte usor.
O serie de factori biotici si abiotici (temperatura, lumina, compusii minerali si organici din apa, oxigenul dizolvat, pH-ul, salinitatea, actiunea valurilor si a curentilor, variatiile de nivel ale apei, prezenta sau absenta vegetatiei, prezenta sau absenta pradatorilor, a parazitilor etc.) influenteaza prezenta si distributia larvelor de culicide intr-un habitat (DABROWSKA-PROT 1977, GIURCA si col. 1983, CIOLPAN 1992, CLEMENTS 1963, PRITCHARD 1983, SKIERSKA 1969).
Apa ca mediu de viata pentru larvele de culicide este esentiala pentru nivelul abundentei acestora. Regimul de precipitatii influenteaza numarul si suprafata habitatelor de dezvoltare a larvelor si implicit este unul dintre factorii cei mai importanti de reglare naturala a efectivelor acestora.
Temperatura influenteaza toate procesele metabolice ale organismului, anumite valori ale sale declansand eclozarea larvelor si determinand apoi rata si viteza lor de crestere si dezvoltare.
Datorita dezvoltarii sezoniere si aparitiei in densitati foarte mari, larvele de culicide reprezinta componente de baza in structura trofica a unor ecosisteme acvatice (in special in cazul unor biocenoze cu o structura simpla). Larvele utilizeaza neselectiv ca hrana, planctonul, precum si materia organica sub forma de particule coloidale si adesea sub forma de detritus. Mai rar se hranesc cu larvele unor insecte sau cu alte organisme vii, observandu-se un pradatorism activ. Ca urmare a modului de hranire omnivor, larvele de culicide se incadreaza in nivelele trofice ale consumatorilor de ordinul I si II (DABROWSKA-PROT 1979, HARD si col. 1984, SERVICE 1969).
La randul lor, ouale si larvele de culicide constituie hrana altor animale acvatice. Daca ouale sunt consumate doar sporadic de catre pradatori (moluste, acarieni, insecte ortoptere, formicide, carabide), reducerea numerica la nivelul stadiilor larvare este de 80-95% in majoritatea biocenozelor acvatice si aceasta se datoreaza in mare parte pradatorilor.
Majoritatea nevertebratelor care se hranesc cu larve de culicide fac parte din urmatoarele grupe: Coelenterata (Hydra), Platyhelmintes (Planaria), Rotifera, Mollusca (Limnea) si Arthropoda cu cel mai mare numar de pradatori, printre care: Crustacea (Entomostraca, Macrura), Arachnida si Insecta cu ordinele: Ephemeroptera, Odonata, Neuroptera, Heteroptera (Notonectidae, Corixidae, Nepidae, Belostomatidae, Naucoridae, Valiidae, Hydrometridae), Coleoptera (Dytiscidae, Gyrinidae, Hydrophylidae, Cicindelidae), Diptera (Chironomidae, Tipulidae, Calliphoridae, Anthomyidae, Dolichopodidae, Empididae). Numeroase vertebrate din grupele Pisces, Amphibia, Reptilia si Aves sunt pradatori ai larvelor de culicide.
La reducerea numerica a populatiilor de larve de culicide, in anumite momente si habitate pe langa pradatori contribuie insa parazitii si patogenii larvelor, reprezentati prin numeroase virusuri, bacterii, fungi, protozoare, nematode, acarieni (JENKINS 1964).
In anumite cazuri, variatia puternica a unor factori, mai ales abiotici, poate determina aparitia unor adevarate epizootii in populatiile larvelor de culicide, datorate inmultirii excesive a unor paraziti sau patogeni ai acestora. Izolarea serotipului H-14 al bacteriei sporogene Bacillus thuringiensis, patogen pentru larvele de culicide, utilizat apoi in controlul acestora, a avut loc tocmai in urma unei epizootii intr-un habitat cu apa puternic poluata organic (de BARJAC 1978, de BARJAC si col. 1980, GOLDBERG & MARGALIT 1977, NICOLESCU 1981, 1982).
Se poate conchide ca larvele de culicide, prin integrarea in circuitul trofic din unele ecosisteme acvatice, participa la circuitul materiei si la transferul energiei in cadrul acestora. Pe de alta parte, ele au o importanta deosebita prin faptul ca se numara printre primele grupe de animale care populeaza habitatele acvatice nou aparute.
Adultii - componente ale ecosistemelor terestre
Fauna de culicide din ecosistemele terestre difera, in cele mai multe cazuri, prin compozitia sa pe specii, de cea din ecosistemele acvatice din care provine. In ecosistemele terestre, structura pe specii a faunei de culicide adulte este determinata de reducerea selectiva a numarului de larve in timpul vietii acvatice, de diferentele interspecifice ale raspunsurilor fata de variatiile factorilor de mediu, de numarul de femele si de fecunditatea lor, de longevitatea lor.
Conditiile de dezvoltare a stadiilor acvatice ale culicidelor determina natura faunei adulte sub diferite aspecte, la diferite niveluri: individual (longevitatea femelelor, fecunditatea lor, posibilitatea autogeniei etc.) ; populational (intensitatea reducerii numerice in mediul acvatic determina numarul initial de adulti, conditiile in care se dezvolta larvele determina relatiile reproductive la nivelul populatiilor etc.); biocenotic (reducerea selectiva a numarului de larve determina structura pe specii a comunitatilor de adulti etc.)
In urma dispersiei, a modificarii diversitatii specifice si a densitatilor numerice (datorate relatiilor biocenotice si factorilor abiotici), fauna de culicide din ecosistemele terestre difera de cea din ecosistemele acvatice din care provine, iar comunitatile de culicide aparute primavara difera de cele din vara sau de cele aparute toamna (SKIERSKA 1965).
Culicidele adulte sunt o prezenta importanta atat in ecosistemele naturale cat si in cele antropizate, determinata in buna masura de relatiile lor trofice, care sunt foarte variate, fiind animale polifage. Nectarul florilor si seva plantelor constituie o importanta sursa de energie pentru toate speciile de culicide (CLEMENTS 1963, SANDHOLM 1962). Daca pentru masculi hrana vegetala constituie singura sursa de energie, pentru femele sangele animalelor reprezinta o sursa suplimentara necesara pentru dezvoltarea oualor. Preferintele de hranire cu sange ale femelelor de culicide pot fi indreptate catre animale aflate in mediul natural (specii exofile) sau catre om si mediul sinantropic (specii endofile). Pe de alta parte, adultii de culicide pot sa reprezinte o prada pentru reprezentantii multor grupe de nevertebrate (arahnide, insecte odonate, hemiptere, himenoptere, coleoptere, diptere) si vertebrate (amfibieni, reptile, pasari, mamifere). De asemenea, ei pot fi infectati sau parazitati de unele virusuri, bacterii, ciuperci, protozoare, nematode, trematode, acarieni (JENKINS 1964).
Rezulta deci ca adultii de culicide participa la circuitul materiei si la transferul energiei in ecosistem, cu consecinte importante la nivel biocenotic (de ex. schimbari la nivelul structurii circuitelor trofice prin densitatile foarte mari realizate in anumite perioade si prin migratie etc.) Dar, datorita hematofagiei lor si posibilitatii pe care o au de a suporta, gazdui si transmite diferite organisme patogene ei au un rol reglator important in numeroase ecosisteme naturale dar mai ales in cele antropizate.
Clasificarea Culicidelor
In general, Familia Culicidae este impartita in trei subfamilii: Toxorhynchitinae (aproximativ 94 de specii), Anophelinae (aproximativ 484 de specii) si Culicinae (aproximativ 2750 de specii). Separarea lor se face pe baza diferentelor existente la nivel morfologic la toate nivelurile de dezvoltare (oua, larve, imago).
Subfamilia Toxorhynchitinae traieste mai ales in zonele tropicale. Sunt tantari de dimensiuni mari. Adultii nu sunt hematofagi deci fara importanta in transmisia unor patogeni. Larvele sunt pradatoare si se pot hrani cu larvele altor specii de culicide. De aici si utilizarea lor in experimente de virologie precum si ca potentiali agenti in lupta biologica.
Subfamilia Anophelinae
Caractere generale: adultul are palpii la fel de lungi ca si trompa la ambele sexe, abdomenul fara solzi, oua-ovale (forma de canoe), prevazute cu flotori laterali. Larva nu are sifon respirator, segmentele abdominale prezinta peri palmati si placi tergale chitinoase. Acesti tantari sunt responsabili, printre altele, de transmiterea formelor de Plasmodium ce provoaca malaria.
Subfamilia Culicinae
Caractere generale: adultul are corpul acoperit de solzi, femelele au palpii scurti. Larva - are sifon respirator. Ouale fara flotori, depuse grupat (barcute) Culex, depuse individual Aedes sau prinse pe plante Mansonia.
Pentru toate cele trei subfamilii este valabila subimpartirea in triburi, genuri si specii.
Pe langa elementele de morfologie externa la identificare se mai utilizeaza si metode genetice sau biochimice.
Genetica
In experimentele de incrucisare si prin cercetarile de morfometrie sau cele ecologice, au fost gasite grupe de populatii distincte genetic precum si specii gemene. Complexele de specii importante sunt: Anopheles gambiae, Anopheles maculipennis, Aedes scutellaris si Culex pipiens. In cazul acestor complexe, speciile pot fi separate cu ajutorul diferentelor evidentiate de tehnicile non-morfologice.
Date Biochimice/Moleculare
Tehnicile biochimice (electroforeza enzimelor si gaz-cromatografia), hidrocarbonatii cuticulari si probele de ADN au fost utilizate cu succes pentru a distinge speciile similare morfologic dintr-un complex de specii. Deoarece tantarii sunt cei mai importanti vectori, s-au efectuat o serie de investigatii biochimice si de biologie moleculara: asupra inhibitorilor ce provoaca coagularea sangelui, asupra interactiunii parazitilor malariei si digestia sangelui sau a reactiilor imunitare ale vectorului, rezistenta la insecticide, vitelogeneza etc.
Importanta medicala a Culicidelor
Culicidele au un important rol epidemiologic si disconfortant, al carui suport este hematofagia lor.
Astfel, culicidele gazduiesc, mentin, transporta si raspandesc organisme cu rol patogen pentru numeroase animale si pentru om, pe zone foarte intinse care devin endemice pentru bolile produse de acestea. De asemenea, culicidele pot determina modificari importante in caracteristicile si comportamentul unor animale salbatice si domestice dar mai ales ale populatiilor umane, cu consecinte grave de diverse feluri, cuantificabile prin efecte economice si sociale negative.
Culicidele ca vectori
Culicidele transmit urmatoarele tipuri de organisme patogene pentru om si animale:
virusuri produc arboviroze cu simptomatologie de tip
encefalitic sau hemoragic ;
bacterii Francisella tularensis
plasmodii protozoare care produc malaria;
- nematode din genurile Wuchereria si Brugia ( fam. Filariidae),
care produc filarioze limfatice.
Culicidele au importanta scazuta in transmisia unor patogeni de tip bacterian. S-au semnalat cazuri de infectie cu agentul tularemiei, Francisella tularensis, la om si la pasari, in transmisie fiind implicate specii de Aëdes (OLSUFIEV 1939).
In cadrul mecanismului de transmitere vectoriala a unei boli, in vector se deruleaza o faza obligatorie a ciclului biologic al agentului patogen (protozoar, nematod) sau o multiplicare a acestuia (virus).
Pe o anumita treapta in evolutia mecanismelor vectoriale s-a ajuns la asocierea obligatorie a unui organism patogen cu un anumit vector: transmisia malariei la om nu se poate face fara anumite specii de anofeli si in lipsa acestora, boala dispare; de asemenea, anumite specii de culicide transmit anumite virusuri sau anumite nematode.
In prezent se stie mai mult despre relatiile agent patogen-gazda vertebrata, dar este tot mai necesara cunoasterea sistemelor agent patogen-vector tantar, pentru ca la acest nivel se poate interveni cu mai multa eficacitate pentru stavilirea transmiterii bolilor respective (RODHAIN 1985).
La nivelul sistemelor agent patogen-vector se succed mai multe fenomene (infectarea vectorului, multiplicarea sau transformarea agentului patogen in organismul vectorului si aparitia formei infectante pentru gazda vertebrata, migrarea acesteia in zona de unde va fi transmisa, eventuala aparitie a posibilitatilor de transmitere transovariana sau sexuala in populatia vectorului, transmisia la gazda vertebrata, alte interactiuni in cadrul sistemului) care se desfasoara numai in anumite conditii, incluzand si pe cele de mediu exterior. Daca aceste conditii nu sunt intrunite, nu se atinge, in final, transmisia bolii.
Infectarea vectorului nu se face decat prin forma infectanta pentru acesta (gametociti de Plasmodium de ex.) cand se atinge pragul de infectivitate (nivelul minim cantitativ al formei infectante, necesar pentru infectare) (CORNEL & JUPP 1989, MEYER si col. 1986, MITCHELL 1983, STÜRCHLER si col. 1990).
In timpul incubatiei extrinseci (perioada de transformare si/sau multiplicare a agentului patogen in vectorul care devine astfel infectant) agentul patogen intampina factori calitativi si cantitativi limitativi la nivelul intestinului, in hemolimfa, in organele tinta (glande salivare, ovare etc.). Pe de alta parte, pentru ca incubatia extrinseca sa fie completa, trebuie ca vectorul sa aiba o supravietuire adecvata, aceasta depinzand in mare masura de conditiile de mediu (BAIN & PETIT 1989, MEYER si col. 1986, RODHAIN 1985); supravietuirea pe perioade mai lungi in populatii abundente a vectorilor in zonele tropicale si ecuatoriale duce la endemii mai puternice in aceste zone (BUSVINE 1980). Cand transmisia transovariana si cea sexuala au frecvente ridicate, populatia vectoare are si rol de rezervor natural pentru agentul patogen respectiv, fenomen frecvent in cazul vectorilor de arbovirusuri (BARDOS si col. 1975, RODHAIN 1985).
La infectarea vertebratului, pe langa receptivitatea acestuia si pe langa starea sistemului agent patogen-vector, intervin si elemente care depind de ecologia proprie sistemului vector infectant-vertebrat (intensitatea contactelor cu vectorii infectanti, conditiile de mediu etc.) (JAENSON 1990, STÜRCHLER si col.1990).
Interactiunile agent patogen-vector se desfasoara in permanenta in ambele sensuri, astfel incat intre masa de agent patogen absorbita de vector si cea transmisa exista diferente notabile iar la randul lor comportamentul si longevitatea vectorului sunt adesea influentate de prezenta agentului patogen, rezultand efecte importante din punct de vedere epidemiologic (HAYES 1978 ).
In cadrul acestor interactiuni se manifesta si mecanisme imunitare in care sunt implicate hemocitele iar sistemele compatibile vector-agent patogen sunt foarte eficiente in contracararea activa a mecanismelor imunitare destructive (CHRISTENSEN & TRACY 1989, RODHAIN 1985).
Realizarea completa a unor cicluri de infectare presupune o coadaptare stransa intre cele doua elemente ale sistemului agent patogen-vector, concretizata in faptul ca pentru un agent patogen dat, intr-un mediu dat, diferiti vectori manifesta o capacitate vectoriala diferita determinata de rezultanta dificultatilor prin care trece agentul patogen la nivelul barierelor succesive intalnite in vector. Mecanismele intime nu sunt cunoscute inca dar controlul este de natura genetica; un rol important este jucat de factorii de mediu, ceea ce explica faptul ca formarea acestor sisteme functionale si adaptative, care sunt sistemele agent patogen-vector, intervine la nivelul populatiilor naturale ale agentului patogen ca si ale vectorului (BAIN si PETIT 1989, CORNEL & JUPP 1989, MEYER si col. 1986, TEODORESCU 1983, TEODORESCU si col. 1978).
In zona noastra este important de urmarit in permanenta capacitatea vectoriala a speciilor de culicide, in cadrul unei epidemiologii preventive (BILBIE si col. 1978, NICOLESCU 1983, NOSEC si col. 1978, TEODORESCU 1975).
Rolul culicidelor in transmisia arbovirusurilor
Arbovirusurile sunt virusuri mentinute in natura prin cicluri biologice de transmisie intre gazdele vertebrate sensibile (pasari, mamifere, reptile) si artropode hematofage. Infectiile produse de acestea sunt zoonoze cu focalitate naturala, larg raspandite pe toate continentele, mai mult de jumatate dintre ele fiind transmise de culicide.
Unele arboviroze sunt boli foarte grave (circa 25), de tip encefalitic, cu implicatii neurologice amintind poliomielita sau de tip hemoragic, cu sangerari din gingii, nas, organe interne. Manifestarile clinice mai putin grave se rezuma la dureri destul de severe in articulatii si oase care pot persista chiar cativa ani si la simptome de tip gripal (JAENSEN 1990, CALISHER 1988, ELDRIGE 1990, HAYES 1989). Intensitatea si durata viremiei variaza la diferite vertebrate dar in general aceasta este scurta. Vectorii reprezinta adesea si principalele surse de infectie, indeplinind si functia de rezervoare, deoarece virusul se mentine pe toata durata vietii vectorului, existand si posibilitatea de transmitere trans-ovariana si trans-sexuala.
Din cele aproape 500 arbovirusuri inregistrate, 49,1% au fost izolate din culicide. Din acestea, 139 virusuri ( 65,9% ) au fost izolate din specii ale genului Culex. Urmeaza genurile Aëdes cu 40,8% dintre virusurile izolate, Anopheles cu 27%, Mansonia cu 12,8%, Coquillettidia cu 11,8%, Psorophora cu 9,5%, Culiseta cu 7,1% si alte 13 genuri a caror pondere insumata reprezinta 18,5% (CALISHER & KARABATSOS 1988, ELDRIGE 1990, MEYER si col. 1986, MITCHEL 1983). Este de mentionat ca genurile Aëdes, Culex, Anopheles, Culiseta si Coquillettidia sunt reprezentate si in fauna tarii noastre (NICOLESCU 1986, 1995).
Arbovirozele transmise de culicide care au importanta cea mai mare prin gravitatea simptomelor si intinderea teritoriilor afectate in lume sunt febra galbena, denga si denga hemoragica, encefalita japoneza, encefalitele California, Saint Louis, encefalitele equine de est si vest (BUSVINE 1980, HAYES 1978, MAURICE & PIERCE 1987, WHO/VBC 89.967, OMS 719-1985).
In Europa, arbovirusurile izolate din culicide apartin urmatoarelor familii:
Togaviridae: Sindbis (ANDERSON & JAENSON 1987, JAENSON 1990, LUNDSTRÖM 1990);
Flaviviridae : West Nile (LABUDA 1983, HANNOUN si col. 1966, FELIPE & de ANDRADE 1990);
Bunyaviridae, cu reprezentanti mai numerosi: Batai (ASPÖCK si col. 1970, BRUDNJAK si col. 1970, JAENSON 1990, SMETANA si col. 1967), Inkoo (BRUMMER-KORVENKONTIO si col. 1973, NIKLASSON 1985, TRAAVIK 1978 ), Tahyna (ARCAN si col. 1975, CHIPPAUX si col. 1970, HANNOUN si col. 1966 a, LABUDA & KOZUCH 1982, LVOV si col. 1985, MALKOVA si col. 1984, ROSICKY & MALKOVA 1980), Lednice (CALISHER & KARABATSOS 1988, MALKOVA si col. 1972) si Zaliv Terpeniya (LVOV si col. 1985). Vectorii acestor virusuri fac parte din cele 5 genuri citate anterior, se gasesc si in tara noastra si sunt urmatorii: Aëdes cinereus, Aë. vexans, Aë. flavescens, Aë. cantans, Aë. sticticus, Aë. communis, Aë. punctor, Aë. caspius, Aë. dorsalis, Culex pipiens, Cx. modestus, Cx. torrentium, Culiseta annulata, Cu. morsitans, Coquillettidia richiardii, Anopheles atroparvus, An. maculipennis s.l.
Circulatia virusurilor West Nile, Tahyna si Batai a fost semnalata si in tara noastra prin depistarea anticorpilor la om si la animale sau prin izolarea virusului din culicide capturate in natura (DRAGANESCU 1979, ARCAN si col. 1974, UNGUREANU si col. 1988, NICOLESCU si col. 1991), stabilindu-se o schema generala de distributie a acestora pe teritoriul tarii. Daca virusul Batai pare ca nu este transmis la om, celelalte doua s-au depistat atat la animale domestice cat si la om. Virusul West Nile are o distributie destul de uniforma pe teritoriul tarii, cu o seropozitivitate de 4,4-7,1%, dar ajungand la 10,2% in populatia umana si in medie de 5%, dar atingand si 17,3% la animale domestice. Virusul Tahyna se gaseste in zona de campie situata in lungul cursului Dunarii, in Dobrogea si Delta Dunarii, unde se inregistreaza o seropozitivitate de 10,5-21,7% in populatia umana si 10,5-22,5% la animale domestice. Circulatia acestor arbovirusuri in zone diferite din tara este legata de conditiile ecologice locale care determina si caracteristicile faunei de culicide vectoare.
In perioada iulie-octombrie 1996, in partea de sud-est a Romaniei a avut loc cea mai mare epidemie de meningoencefalita cu virusul West Nile care s-a inregistrat pana in prezent (TSAI et al. 1998; NICOLESCU 1998). Acest flavivirus este transmis intr-un ciclu pasari tantari in Africa si Asia dar in Europa s-a raportat sporadic si niciodata in astfel de proportii epidemice. Infectarea mamiferelor si a omului duce la un capat mort al ciclului de transmisie, deoarece viremia este foarte scurta la acestea si reinfectarea unor culicide de la aceste organisme este sporadica si extrem de scazuta.
Aspectele relevate de epidemia de meningoencefalita provocata de virusul West Nile in Romania arata ca "febra cu West Nile" (gripa de vara) este o boala care isi face aparitia in Europa unde se poate manifesta sub formele ei cele mai grave. De aceea, in prezent este necesara extinderea si aprofundarea cunoasterii populatiilor de culicide potential vectoare pentru arbovirusuri, inclusiv pentru virusul West Nile ca si a sistemelor vectoriale existente la noi in tara si a implicarii lor in patologia umana, in vederea prevenirii si combaterii unor imbolnaviri grave la om.
Rolul culicidelor in transmisia protozoarelor din Genul Plasmodium
Malaria este produsa de catre protozoare din genul Plasmodium, patru specii infectand omul: falciparum, vivax, malariae si ovale.
Ciclul de transmisie se deruleaza astfel: formele sexuate ale agentului patogen (gametocitii) sunt ingerate de catre femelele unor specii din genul Anopheles la hranirea cu sangele unui bolnav; agentul patogen isi desfasoara ciclul sporogonic in anofel rezultand sporozoiti infectanti pentru om; in timp ce femela de anofel se hraneste cu sange, acestia sunt inoculati unei gazde umane sensibile, in care are loc ciclul schizogonic care duce la aparitia gametocitilor, forma infectanta pentru femela de anofel.
Simptomele clinice la om apar, in general, dupa 8-30 zile de la inocularea sporozoitilor infectanti. Ele constau din accese succesive tipice de febra, insotite de frisoane si de transpiratie. Accesele coincid cu multiplicarea sincrona a agentului patogen in hematii urmata de aparitia sa in circulatia sanguina prin ruperea acestora. Pe masura ce creste abundenta sa in sange, simptomele se intensifica. Parazitemia ridicata poate produce ea insasi moartea sau aceasta intervine prin complicatii cerebrale, renale sau pulmonare (KWIATKOWSKI 1989, STÜRCHLER si col. 1990, WARREL si col. 1990.
Malaria terta maligna (cu falciparum) este forma cea mai periculoasa a bolii. Malaria terta benigna (cu vivax) si malaria cvarta (cu malariae) nu pun viata in pericol dar se pot caracteriza prin simptome severe. Dupa reinsanatosire, in urma tratamentului, se pot inregistra recaderi la intervale de pana la un an (falciparum), pana la 5 ani (vivax si ovale ) si chiar dupa 20-25 de ani (malariae) (CIUCA 1966, MAURICE si PEARCE 1987).
In prezent malaria este boala endemica cu raspandirea cea mai mare pe glob. Estimarile cele mai recente (TDR News, No. 41 1992 ) arata ca 300 milioane de persoane sunt infectate in lume (275 milioane in Africa), exista peste 120 milioane cazuri clinice pe an (110 milioane in Africa) iar numarul anual de decese este de 1,1 milioane (1 milion in Africa).
Existenta malariei se face simtita pe zona ecuatoriala si tropicala din cele doua emisfere care depaseste limitele teoretice reprezentate de izotermele de + 15°C (temperatura necesara pentru completarea ciclului sporogonic in anofel pentru P. vivax ) si + 21°C (pentru P. falciparum). In afara acestor limite, malaria poate sa apara in numeroase habitate in care adultii de culicide gasesc locuri de adapostire cu microclimat adecvat completarii sporogoniei (BOYD 1949, GARNHAM 1945, WHO/VBC 89.967, SCHULTZ 1989).
Eforturile actuale de stavilire a malariei sunt ingreunate de numerosi factori: rezistenta tulpinilor de Plasmodium fata de medicamente, toxicitatea ridicata a acestora, rezistenta culicidelor fata de insecticide dar mai ales cunoasterea inadecvata a epidemiologiei locale a malariei (HAWORTH 1988). Cresterea incidentei malariei in ultimii ani se datoreaza si altor cauze: cresterea temperaturii globale din cauza efectului de sera in urma poluarii, miscari masive de populatie determinate de existenta unor conflicte armate pe zone intinse, circulatia intensa in scopuri turistice, de afaceri etc., procese de urbanizare haotica si rapida etc. (HAWORTH 1988, MAURICE si PEARCE 1987, SCHULTZ 1989, JETTEN & TAKKEN 1994).
Dupa anii 70 OMS a elaborat conceptul de "stratificare a malariei" pentru delimitarea si clasificarea zonelor endemice de pe glob. Acestea reprezinta zone epidemiologic omogene, legate de prezenta anumitor specii de anofeli vectori, legati la randul lor de anumite conditii de mediu, in care se pot aplica programe adecvate de supraveghere si control (OMS 735-1986, 1974, 680-1982). De aceea, in prezent se tine seama de cele 12 mari zone epidemiologice de malarie din lume, stabilite inca din 1957 de catre McDONALD (WHO/VBC 89.967). In interiorul acestor zone principale se pot diferentia zone din ce in ce mai mici, cu caracteristici epidemiologice foarte precise, in care s-ar putea aplica programe coerente si adecvate de stavilire a epidemiilor.
In zona europeana malaria a disparut in cursul acestui secol, cu exceptia recaderilor cu P. malariae. Cu toate acestea, cateva mii de cazuri importate se semnaleaza anual astfel incat, in ultimii 10 ani ele au insumat mai mult de 45.000 (HAWORTH 1988, PHILLIPS-HOWARD 1990, SCHULTZ 1989). In ultimele doua decenii s-au inregistrat si cateva zeci de cazuri aparent autohtone de malarie, la rezidenti din imediata vecinatate a unor aeroporturi din Belgia, Franta, Olanda, Elvetia, Marea Britanie, care pot fi atribuite unor anofeli tropicali infectati, adusi cu avioanele din zone endemice. Acestia au supravietuit in timpul verii in zonele invecinate aeroporturilor si au putut transmite agentul patogen (CURTIS & WHITE 1984, HAWORTH 1988 ). De aceea, in prezent exista temeri ca malaria ar putea fi reintrodusa in Europa, cel putin in zonele ei sudice (JETTEN & TAKKEN 1994).
Rolul culicidelor in transmisia nematodelor producatoare de filarioze
Filariozele limfatice apar in focare tropicale umede din Africa, cele doua Americi, Asia si numeroase insule din oceanul Pacific (HAWKING & DENHAM 1976, HAWKING 1876, 1977, 1979) fiind produse de viermii Brugia malayi si B. timori (filarioza brugiana) si Wuchereria bancrofti (filarioza bancroftiana) (BAIN & PETIT 1989, WHO / VBC / 89.967).
Spre deosebire de malarie sau arboviroze, care pot fi transmise prin intepatura unui singur vector infectant, infectiile cu filarii necesita inoculari repetate cu larve infectante, probabil sute pe an, pentru ca viermii sa se reproduca eficient si sa produca microfilaremia. Adesea, boala este asimptomatica initial, cu episoade ulterioare de inflamare acuta a sistemului limfatic si febra, cu un stadiu cronic de "elefantiazis" care se dezvolta dupa mai multi ani sau nu, timp in care microfilariile dispar din circulatie (BAIN & PETIT 1989).
Multe dintre speciile de Anopheles care sunt vectori principali de malarie in zonele respective sunt si vectori pentru filarii. La acestia se adauga diferite specii de Aëdes, Culex si Mansonia. In focarele urbane, vectorii sunt inevitabil Culex quinquefasciatus in zonele tropicale sau Cx. pipiens pallens in cele temperate. Toate cele trei specii de paraziti citate au de obicei o periodicitate nocturna a microfilariilor in sangele periferic al gazdei, aceasta fiind o adaptare pentru a fi luate si transmise de catre vectori care se hranesc numai noaptea. Exista situatii cand tulpini subperiodice ale acestor paraziti (B. malayi si W. bancrofti) s-au adaptat pentru transmisia de catre vectori activi in timpul zilei din genurile Aëdes (Aë. scutellaris, Aë. niveus) si Mansonia.
Culicidele ca generatoare de disconfort
Culicidele sunt generatoare de disconfort prin prezenta lor in populatii extrem de abundente in anumite zone si momente si prin atacurile lor nediscriminante asupra omului si a mamiferelor mari, domestice sau salbatice, atat ziua, cat si noaptea (EDMAN 1989, OMS 1988). Din acest motiv unele regiuni pot fi handicapate in dezvoltarea lor economica. Poate fi afectata punerea in valoare a unor zone de interes turistic, pot fi incomodate activitatile legate de agricultura, silvicultura, constructiile hidrotehnice, de drumuri. Animalele domestice pot fi impiedicate sa-si desfasoare normal munca, hranirea si odihna, ceea ce duce la scaderea randamentului si la diminuarea cantitativa si calitativa a productiei de lapte si carne.
Pe langa neajunsurile provocate de diminuarea perioadei de somn, actiunea disconfortanta a culicidelor include si efectele substantelor toxice pe care femelele le elibereaza odata cu saliva la locul intepaturii, concretizate printr-o gama foarte larga de reactii de tip alergic ale gazdei. Gravitatea si caracterul acestora sunt corelate cu specia de culicid, cu numarul de intepaturi, cu sensibilitatea gazdei, astfel incat ele pot fi de la aproape imperceptibile sau marginite la locul intepaturii pana la reactii uneori generalizate in urma sensibilizarii. Adesea apar suprainfectii in urma transmiterii de catre culicide, in mod mecanic, a unor bacterii sau ca urmare a gratajului (HERMS & JAMES 1961).
Procesul de urbanizare accelerata si necontrolata duce, prin modificari ale habitatului, la aparitia unor conditii favorabile dezvoltarii anumitor specii de culicide, adevarati indicatori pentru gradul de antropizare a mediului (COUSSERANS 1975, COUSSERANS si col. 1976, ROBINSON & ATKINS 1983, SCHOFIELD & WHITE 1984).
Rolul disconfortant cel mai accentuat in mediul urban la noi se datoreaza speciei Culex pipiens, cu formele sale de exterior si interior (NICOLESCU 1983, 1986). Larvele acestei specii prefera apele poluate, incarcate cu materii organice, chiar pe cele lipsite complet de oxigen, deoarece ele respira aerul atmosferic. Larvele formei de exterior populeaza habitate acvatice exterioare. In aglomerarile de tip urban sau in zone turistice, ele se dezvolta in orice mica balta pe sol, in zone mlastinoase, multe dintre ele periurbane, in apele uzate deversate necorespunzator, in obiecte aruncate care pot retine apa etc. Adesea, densitatile lor sunt impresionante. Larvele formei molestus a speciei Cx. pipiens populeaza acumulari de apa care se formeaza in interioare de felul subsolurilor inundate ale multor blocuri. Apa este bogata in material organic, temperatura se mentine destul de ridicata in tot timpul anului, ceea ce permite dezvoltarea permanenta a larvelor. Femelele acestei forme sunt autogene, fiind capabile sa depuna cel putin o ponta fara sa se hraneasca cu sange dar cand au posibilitatea sa o faca, ele manifesta o puternica antropofilie. In timpul sezonului cald, abundenta populatiilor de Cx. pipiens creste foarte mult; adultii formei molestus ies de cele mai multe ori si in exterior, depun oua si larvele lor se dezvolta impreuna cu cele ale formei de exterior in extrem de numeroasele habitate caracteristice acestei specii.
Prezenta culicidelor din genul Aëdes se face simtita mai ales primavara si la inceputul verii, in special in parcuri, in zone periurbane, in zone turistice, in paduri, pe campii inundabile sau irigate, acolo unde se gasesc si habitatele favorabile dezvoltarii larvelor. Acestea sunt zone cu solul destul de umed, adesea inierbat, care formeaza mici gropi, santuri sau denivelari, unde toamna femelele depun ouale direct pe sol iar primavara prin inundare se formeaza balti temporare in care se dezvolta larvele speciilor de Aëdes. Adesea, in zonele urbane se inregistreaza si o crestere a anofelismului, mai ales in apropierea unor lacuri insuficient ingrijite, larvele de anofeli preferand ochiurile de apa partial umbrite de vegetatia de pe maluri si placardele de vegetatie care plutesc pe suprafata apei. Specii din genurile Culiseta, Coquillettidia si Uranotaenia apar uneori in zonele urbane sau in cele turistice, fiind legate de anumite habitate specifice. Astfel, speciile de Coquillettidia isi fac aparitia la sfarsitul lunii mai inceputul lunii iunie si se mentin doua-trei luni in apropierea ecosistemelor acvatice cu mult stuf si papura, in care larvele lor se dezvolta fixate cu sifonul respirator modificat pe partile submerse ale acestor plante. Majoritatea speciilor apartinand acestor genuri sunt in mare masura antropofile si de aceea prezenta lor provoaca un puternic disconfort.
Problema disconfortului produs de culicide este strans legata de rolul vector al acestora in comunitatile umane unde provoaca de obicei ambele neajunsuri (RODHAIN 1983, OMS 1988). De aceea, necesitatea mentinerii unui anumit nivel al densitatii populatiilor de culicide intr-o aglomerare umana are dublul scop de a evita orice risc epidemic si de a mentine cel putin un grad minim de confort pentru populatie (BILBIE si NICOLESCU 1986).
Atingerea acestor obiective se realizeaza numai prin aplicarea unor programe de control integrat al culicidelor, adaptate cadrului ecologic, programe de o importanta socio-economica si medico-sanitara deosebita in anumite comunitati umane.
Un rol esential in elaborarea si aplicarea acestor programe este cunoasterea exacta a locului si rolului culicidelor in ecosistemele vizate astfel incat, efectele benefice asteptate in plan socio-economic si al sanatatii publice prin aplicarea acestor programe sa nu fie umbrite pe termen mai scurt sau mai lung de dezechilibre ecologice care in ultima instanta se rasfrang tot asupra sanatatii omului.
Aspecte privind Culicidele din Romania
La noi in tara au fost semnalate pana in prezent 56 de specii de culicide. Ele sunt incluse in 8 genuri si 15 subgenuri. Pozitia lor sistematica este urmatoarea: Ordinul Diptera, Subordinul Nematocera, Familia Culicidae cu Subfamiliile: Anophelinae (genul Anopheles Plansa 16, 17 si Culicinae (genurile: Culex Plansa 20, 21, Coquillettidia Plansa 15, 21, Culiseta- Plansa 17, Uranotaenia Plansa 21, Orthopodomyia Plansa 21 si tribul Aedes Plansa 18, 19
"Limnofauna Europaea" (1968), prezinta regiunile zoogeografice, fauna acvatica caracteristica inclusiv distributia speciilor de culicide (semnalate si posibile).
Tara noastra este impartita in patru regiuni zoogeografice:
Zona carpatica 10 ce cuprinde lantul carpatin si zona circumscrisa de acesta ce se intinde in Ucraina si Ungaria;
Pusta ungara 11 cuprinde zona de vest a tarii ce se intinde si in Ungaria si partial in Iugoslavia;
Provincia pontica 12 cuprinde Campia Dunarii si trece in Moldova;
Estul sesului european 16 ce cuprinde N-E extracarpatic cu extindere in Moldova.
La sud de Dunare se afla regiunile Balcanica de Est 7 si de Vest-5 ce acopera teritorii din Bulgaria si respectiv Serbia (PLAN A
Corespunzator acestei zonari se face si distributia pe specii a culicidelor: in fostul U.R.S.S. s-au semnalat 60 de specii de aedini (56, 57), in Ungaria 23 de specii (89, 90), in Iugoslavia 21 de specii (136) si 15 specii in Bulgaria (18).
Tribul Aedes ocupa locul dominant este reprezentat prin subgenurile: Aedes (2 specii), Aedimorphus (1 specie), Finlaya (1 specie) si Ochlerotatus (21 specii). Pe baza unor caractere taxonomice subgenul Ochlerotatus a fost impartit in mai multe grupuri de specii: Rusticus, Intrudens, Dorsalis-Caspius, Annulipes-Cantans si Communis.
Dupa arealul de raspandire, speciile de aedini apartin regiunii palearctice. Datele prezentate pe baza Codului Biocartografic, autor A. Lehrer, 1977, realizat pe baza retelei UTM (Universal Transverse Mercator (PLANSA 14), ne permit sa ne facem doar o imagine partiala asupra diversitatii specifice si a raspandirii aedinilor la noi in tara. Urmeaza ca cercetari ulterioare sa ne completeze cunostintele.
Analiza zoogeografica a faunei de culicide din Romania s-a facut pe baza simplei semnalari in teritoriu, datele avute la dispozitie nu ofera suportul unor consideratii ecologice iar criteriul arealistic (specii palearctice) nu permite si o apreciere cantitativa.
Lista speciilor, arealul lor de raspandire precum si distributia lor geografica pe teritoriul tarii noastre sunt realizate pe baza datelor din literatura si pe baza unor observatii personale (Tabel 3).
Se poate constata ca speciile cel mai des semnalate au fost: Aedes vexans, Aedes caspius, Aedes geniculatus, urmeaza Aedes annulipes, Aedes cantans, Aedes cinereus, Aedes excrucians, Aedes pulchritarsis, Aedes communis si Aedes sticticus. Foarte rar au fost semnalate speciile: Aedes rossicus, Aedes behningii, Aedes refiki, Aedes riparius, Aedes intrudens si Aedes punctor. A mai fost semnalata specia Aedes mariae in Delta Dunarii care ulterior a fost confirmat ca Aedes zammitii, ce are ca areal partea de est a Mediteranei. Ultimele date arata ca specia Aedes duplex este sinonim cu Aedes caspius.
Tabel 3. Lista culicidelor din Romania.
|
Nr. crt |
S P E C I A |
ZONA |
|||||||
|
ROMANIA | |||||||||
|
GENUL AEDES Meigen, 1818 |
|||||||||
|
Subgenul AEDES Meigen 1818 |
|||||||||
|
Ae. cinereus Meigen 1818 | |||||||||
|
Ae. rossicus Dolb.Gor. Mitrof. | |||||||||
|
Subgenul A E D I M O R P H U S Theobald 1903 |
|||||||||
|
Ae.vexans Meigen 1830 | |||||||||
|
Subgenul FINLAYA Theobald 1903 |
|||||||||
|
Fin.geniculatus Olivier 1791 | |||||||||
|
Subgenul OCHLEROTATUS Lynch Arribalzaga 1891 |
|||||||||
|
Oc. annulipes Meigen 1830 | |||||||||
|
Oc. behningi Martini 1926 | |||||||||
|
Oc. cantans Meigen 1818 | |||||||||
|
Oc. caspius Pallas 1771 | |||||||||
|
Oc. cataphylla Dyar 1916 | |||||||||
|
Oc.communis De Geer 1776 | |||||||||
|
Oc. cyprius Ludlow 1919 | |||||||||
|
Oc. detritus Haliday 1833 | |||||||||
|
Oc.diantaeus How.Dyar, Knab 1912 | |||||||||
|
Oc. dorsalis Meigen 1830 | |||||||||
|
Oc. euedes How.Dyar,Knab 1912 | |||||||||
|
Oc. excrucians Walker 1856 | |||||||||
|
Oc. flavescens Müller 1764 | |||||||||
|
Oc. intrudens Dyar 1919 | |||||||||
|
Oc .leucomelas Meigen 1804 | |||||||||
|
Oc. nigrinus Eckstein 1918 | |||||||||
|
Oc. pulchritarsis Rondani 1872 | |||||||||
|
Oc. pullatus Coquillett 1904 | |||||||||
|
Oc. punctor Kirby 1837 | |||||||||
|
Oc. riparius Dyar,Knab 1907 | |||||||||
|
Oc. sticticus Meigen 1838 | |||||||||
|
Oc.subdiversus Martini 1926 | |||||||||
|
Oc. thibaulti Dyar,Knab 1909 | |||||||||
|
Oc. zammitii Theobald 1903 | |||||||||
|
Subgenul RUSTICOIDUS Shevchenko&Prudkina, 1973 |
|||||||||
|
Ru. refiki Medschid 1928 | |||||||||
|
Ru. rusticus Rossi 1790 | |||||||||
|
GENUL CULEX Linnaeus, 1758 |
|||||||||
|
Subgenul CULEX Linnaeus, 1758 |
|||||||||
|
Culex mimeticus Noe 1889 | |||||||||
|
Culex pipiens Linnaeus 1758 | |||||||||
|
Culex theileri Theobald 1903 | |||||||||
|
Culex torrentium Martini 1925 | |||||||||
|
Subgenul NEOCULEX Dyar, 1905 |
|||||||||
|
Neoculex impudicus Ficalbi 1890 | |||||||||
|
Neoculex martinii Medschid 1930 | |||||||||
|
Neoculex territans Walker 1856 | |||||||||
|
Subgenul BARRAUDIUS Edwards, 1921 |
|||||||||
|
Barraudius modestus Ficalbi 1889 | |||||||||
|
Subgenul MAILLOTIA Theobald, 1907 |
|||||||||
|
Maillotia hortensis Ficalbi 1889 | |||||||||
|
GENUL COQUILLETTIDIA Dyar, 1905 |
|||||||||
|
Cq. buxtoni Edwards 1925 | |||||||||
|
Cq. richiardi Ficalbi 1889 | |||||||||
|
GENUL URANOTAENIA Lynch-Arribalzaga, 1891 |
|||||||||
|
Pseudoficalbia unguiculata Edwards 1913 | |||||||||
|
GENUL ORTHOPODOMYIA Theobald, 1904 |
|||||||||
|
Or. pulchripalpis Rondani 1872 | |||||||||
|
GENUL ANOPHELES Meigen, 1818 |
|||||||||
|
An. atroparvus VanThiel 1927 | |||||||||
|
An. claviger (Meigen 1804) | |||||||||
|
An. daciae Linton, Niculescu & Harbach, 2004 | |||||||||
|
An. hyrcanus (Pallas 1771) | |||||||||
|
An. maculipennis Meigen 1818 | |||||||||
|
An. melanoon | |||||||||
|
An. messeae Falleroni 1926 | |||||||||
|
An. plumbaeus Stephens 1828 | |||||||||
|
An. sacharovi Favre 1903 | |||||||||
|
GENUL CULISETA Felt, 1904 |
|||||||||
|
Subgenul CULISETA Felt, 1904 |
|||||||||
|
Cu alaskaensis Ludlow 1906 | |||||||||
|
Cu annulata Schrank 1776 | |||||||||
|
Cu.glaphyroptera Schiner 1864 | |||||||||
|
Cu.subochrea Edwards 1921 | |||||||||
|
Subgenul CULICELLA Felt, 1904 |
|||||||||
|
Cul. fumipennis Theobald 1901 | |||||||||
|
Cul morsitans Stephens 1825 | |||||||||
|
Cul. silvestris Shingarev 1928 | |||||||||
|
Subgenul ALLOTHEOBALDIA Brolemann, 1919 |
|||||||||
|
Allotheobaldia longiareolata (Macquart 1919) | |||||||||
Legenda: + specia a fost semnalata; ? posibil ca specia sa se afle in zona; specia nu ajunge in aceasta zona.
Profilaxia
Fileele pentru pat si perdelele ofera o protectie mecanica impotriva tantarilor activi noaptea. Substantele repelente pot fi aplicate atat pe filee cat si pe asternuturi. Aplicarea pe piele are o valoare redusa deoarece transpiratia le deterioreaza. Lampile UV atrag numai masculii ne-hematofagi.
In plus, datorita riscului permanent, expunerea trebuie redusa in acele momente ale zilei in care vectorii respectivi sunt activi: unele specii de Aedes sunt active dimineata si seara. Majoritatea speciilor de Culex, Mansonia si Anopheles inteapa mai ales noaptea. Produsele anti-tantari ce produc un fum insecticid sunt larg utilizate in dormitoare.
Controlul
Campaniile pentru control pot viza larvele sau adultii. In Europa, in incercarea de a controla larvele, cele mai bune rezultate au fost obtinute prin drenarea mlastinilor. Daca speciile vectoare se reproduc in alte tipuri de habitate, succesul este indoielnic daca costurile ridicate nu pot fi acoperite sau daca alte elemente caracterizeaza mediul. Au fost utilizate si alte metode fizice, alese in functie de specia de vector ca: reinundarea intermitenta, despaduriri sau plantarea vegetatiei. In unele tari tropicale educatia reduce numarul zonelor de inmultire create prin depozitarea anvelopelor vechi, a canistrelor si a butoaielor sau a altor recipiente. Controlul chimic include stropirea suprafetei apei cu uleiuri minerale sau prin aplicarea acetoarseniatului de cupru (verde de Paris) prafuri ale altor insecticide (carbamati, organofosfati, piretroizi). Regulatorii de crestere ai insectelor opresc dezvoltarea larvelor sau interfera cu formarea cuticulei. In zonele sensibile sub aspect ecologic, controlul biologic se realizeaza in special cu ajutorul bacteriilor Bacillus thuringiensis var. israelensis si Bacillus sphaericus. Sporii lor contin o endotoxina cristalina ce induce liza celulelor din intestinul mediu al larvelor. Intr-o mai mica masura este utilizat nematodul Romanomaermis culicivorax si pestele larvivor Gambusia.
Controlul adultilor include metodele mentionate la aspectele de profilaxie : utilizarea plaselor anti-tantari impregnate sau nu cu insecticide, folosirea repelentilor sau a unor unguente anti-tantari. Insecticidele sunt adesea folosite impotriva tantarilor adulti; la tratamentul populatiilor pot fi utilizate masini, helicoptere, avioane. Insecticidele reziduale pot fi raspandite in locuinte dar populatiile pot supravietui daca tantarii parasesc casa imediat dupa hranire fara sa vina in contact cu peretii impregnati cu insecticide.
Rezistenta fata de diferitele insecticide s-a dezvoltat cel mai adesea datorita utilizarii acestora in campaniile de protectie a productiilor agricole.
Bibliografie
Bilbie Ionela, Nicolescu Gabriela . Insecte vectoare si generatoare de disconfort. [Vector and nuisance insects]. Editura Medicala, Bucuresti, 346 pp.
Ciolpan O. . Culicidele ca vectori ai unor agenti patogeni. [Mosquitoes as vectors of pathogens]. Bacteriologia, Virusologia, Parazitologia, Epidemiologia, XXXIII, 3: 207-223.
Ciolpan O., Nicolescu Gabriela, Velehorschi N., Ceianu Cornelia, Marcu Mihaela. . Investigatii asupra faunei de tantari (Diptera: Culicidae) din padurea Cernica, potential vectori pentru arbovirusuri. [Investigations on the mosquitoes (Diptera: Culicidae) potential vectors of arboviruses in the Cernica forest]. Analele Banatului, Stiintele Naturii 2, Editura Muzeul Banatului, Timisoara, 356-360.
Ciolpan O. . Bazele ecologice ale combaterii unor specii de tantari din genul Aedes. [The ecological basis of the mosquito control of some Aedes species]. Thesis. Universitatea Bucuresti.
Ciolpan O., Nicolescu Gabriela, Ceianu Cornelia, Ciugulea I. . Identification and characterization of ecosystems (as concerning biotic and abiotic factors) including specific habitats favourable to the development of Culicidae species. Romanian Archives of Microbiology and Immunology, 50, 4: 309-310.
Ciolpan O., Nicolescu Gabriela, Ceianu Cornelia, Vlad Monica . Observatii asupra asociatiilor de Culicidae din padurea Comana. [Investigations on the associations of mosquito species in the forest Comana]. Sesiunea Stiintifica a Institutului Cantacuzino, 29-30 noiembrie 1993, 64.
Ciolpan O., Nicolescu Gabriela, Velehorschi N., Ceianu Cornelia Cercetari asupra populatiilor de tantari (Diptera: Culicidae) in vederea fundamentarii combaterii lor pe baze ecologice. I. Studiul populatiilor de culicide juvenili din padurea Cernica. [Investigations upon the mosquito populations (Diptera: Culicidae) for their control on ecological basis. I. The study of larval mosquito populations in the Cernica forest]. The 6th National Conference of General and Applied Entomology, 27-29 October 1995, Iasi, Romania.
Ciolpan O., Nicolescu Gabriela, Velehorschi N., Ceianu Cornelia Cercetari asupra populatiilor de tantari (Diptera: Culicidae) in vederea fundamentarii combaterii lor pe baze ecologice. II. Studiul asociatiilor de culicide-adulti din padurea Cernica. [Investigations upon the mosquito populations (Diptera: Culicidae) for their control on ecological basis. II. The study of adult mosquito associations in the Cernica forest]. A VI-a Conferinta Nationala de Entomologie Generala si Aplicata, 27-29 octombrie 1995, Iasi, Romania.
Ciolpan O., Nicolescu Gabriela, Pop G. The mosquitoes (Diptera: Culicidae) in the area of the middle course of the river Somesul Mare (Romania): faunistical and ecological data. Romanian Archives of Microbiology and Immunology, 57, 1: 77-91.
Giurca Ileana . Note on the Aëdes species of mosquitoes recorded in Romania. Archives Roumaines de Pathologie experimentale et de Microbiologie, 41, 1: 73-84.
Giurca Ileana 1984 The Aëdes mosquitoes (Diptera: Culicidae) in the East of Romanian Plain. Distribution and biology. Archives Roumaines de Pathologie experimentale et de Microbiologie, 43, 1: 103-111.
Giurca Ileana, Velehorschi N., Nicolescu Gabriela . The mosquitoes in the forest of Cernica. Ecological data. Archives Roumaines de Pathologie experimentale et de Microbiologie, 42, 4: 353-363.
Nicolescu Gabriela 1986. Tantarii. In: Insecte vectoare si generatoare de disconfort. Red. Ionela Bilbie, Gabriela Nicolescu [Mosquitoes. In : Vector and nuisance insects. Eds. Ionela Bilbie & Gabriela Nicolescu], 18-103, Editura Medicala, Bucuresti.
Nicolescu Gabriela, Ceianu Cornelia, Velehorschi N., Ciolpan O., Chirila Eugenia, Petrescu Ana Maria, Teodorescu G. Evaluarea in conditii de teren a insecticidului bacterian I.C. obtinut din Bacillus thuringiensis israelensis. [Field evaluation of the I.C. bacterial insecticide from Bacillus thuringiensis israelensis]. Al VI-lea Simpozion de Microbiologie Industriala si Biotehnologie, septembrie 1987, Iasi, Romania.
Nicolescu Gabriela, Ciolpan O., Ceianu Cornelia, Marcu Mihaela Program in BASIC pentru calculul activitatii insecticide a preparatelor din Bacillus thuringiensis H-14 utilizate ca larvicide in combaterea culicidelor. [Program in BASIC for the calculation of insecticide activity of the preparations of Bacillus thuringiensis H-14 used as larvicides in the control of culicids]. Analele Banatului, Stiintele Naturii 2: 370-373.
Nicolescu Gabriela, Ceianu Cornelia, Ciolpan O., Chirila Eugenia, Petrescu Ana Maria, Vacaru Mariana, Velehorschi N., Marcu Mihaela, Ciugulea I., Mistretu M. Efectiveness of the bacterial insecticide, I. C. - Romthurin on the mosquitoes. Roumanian Archives of Microbiology and Immunology, 50, 4: 268.
Nicolescu Gabriela, Ungureanu Aurora, Ciolpan O., Velehorschi N., Ceianu Cornelia, Ciugulea I., Marcu Mihaela, Catanas Felicia, Ionita Ioana, Safta M., Tutoveanu A., Variu S. Investigations upon the circulation of some arboviruses transmitted by mosquitoes (Diptera: Culicidae) and ticks (Acarina: Ixodidae) in Romania. Romanian Archives of Microbiology and Immunology, 50, 4: 284.
Nicolescu Gabriela, Ceianu Cornelia, Ciolpan O., Velehorschi N., Marcu Mihaela, Ciugulea I. Quantitative and qualitative dynamics of anopheline populations in terms of the environmental factors in the formerly malaria endemic areas in the Romanian Plain and Dobrudja. Romanian Archives of Microbiology and Immunology, 50, 4: 310.
Nicolescu Gabriela, Ceianu Cornelia, Petrescu Ana Maria, Chirila Eugenia, Ciolpan O., Vacaru Mariana, Velehorschi N., Marcu Mihaela, Ciugulea I., Mistretu M. Study of the ecological aspects concerning the use of preparations obtained from Bacillus sphaericus in controlling Culicidae. Romanian Archives of Microbiology and Immunology, 50, 4: 311.
Nicolescu Gabriela, Ciolpan O., Ceianu Cornelia, Ciugulea I., Chirila Eugenia, Petrescu Ana Maria, Vacaru Mariana, Mistretu M. Investigarea conditiilor de introducere a insecticidului bacterian Romthurin (din Bacillus thuringiensis serotip H-14) in programele de combatere integrata a tantarilor (Diptera: Culicidae). [Investigation of the conditions of the introduction of the bacterial insecticide Romthurin (from Bacillus thuringiensis serotype H-14) in the integrated control programmes of mosquitoes (Diptera: Culicidae). Proceedings of the Second National Conference on the Environmental Protection - Biological and Biotechnological Methods, 29-31 May 1992, Brasov, Romania, 49-50.
Nicolescu Gabriela, Gaube Rodica, Ceianu Cornelia, Ciolpan O., Vlad Monica, Hoanca D. Investigatii asupra starii de sensibilitate / rezistenta a anofelilor potential vectori de malarie din fostele zone endemice din tara noastra. [Investigations upon the susceptibility / resistance status of the potential vector anophelines in the previously malaria endemic areas in Romania]. Book of Abstracts of the Scientific Session of Cantacuzino Institute, Bucuresti, 29-30 November 1993, 63.
Nicolescu Gabriela, Ceianu Cornelia, Ciolpan O., Sandu Monica, Hoanca D., Vladimirescu A., Stanca-Mustea Irina Predictive elements of the evolution of the anopheline populations and the risk of reintroduction of malaria in Romania. Romanian Archives of Microbiology and Immunology, 55, 1: 70.
Nicolescu Gabriela, Ciolpan O., Velehorschi N., Ceianu Cornelia, Giurca Ileana, Petrescu Simona, Radutu C. Investigatii faunistico-ecologice asupra tantarilor (Diptera: Culicidae) potential vectori de arbovirusuri in sudul tarii. [Faunistical and ecological investigations on the mosquitoes (Diptera: Culicidae), the potential vectors of arboviruses in the South of Romania]. Book of Abstracts of the Symposium: Epidemic of acute meningo-encephalitis in July - October 1996, November 1996, Bucuresti, 29- 30.
Nicolescu Gabriela, Ceianu Cornelia, Vladimirescu A., Ciolpan O., Sandu Monica, Hoanca D., Stanca-Mustea Irina Evolutia anofelismului si a riscului de reintroducere a malariei in Romania. [Evolution of the anophelism and the risk of malaria reintroduction in Romania]. Book of Abstracts of the 4th Congress of the epidemiologists, microbiologists and parasitologists from Republic of Moldova, 12 September 1997, Chisinau, 2 (b), 178-179.
Nicolescu Gabriela, Ceianu Cornelia, Vladimirescu A., Ciulacu-Purcarea Valeria, Ciolpan O., Marcu Mihaela, Ciugulea I. Evaluation of the risk of malaria reintroduction in Romania. Proceedings of the 1st Balkan Conference "Malaria and Mosquito Control", 5th - 7th April 2001, Lithotopos, Serres, Greece, 43- 45.
Gabriela Nicolescu, Valeria Purcarea-Ciulacu, Elena Claudia Coipan - 2001. A REVISED BIBLIOGRAPHY OF THE MOSQUITOES OF ROMANIA. Romanian Archives of Microbiology and Immunology, T 60, No. 2, 131-175.
Velehorschi N., Nicolescu Gabriela, Ceianu Cornelia, Giurca Ileana, Bilbie Ionela . The mosquitoes (Diptera: Culicidae) in Maliuc (Danube Delta) - faunistical and ecological data. Archives Roumaines de Pathologie experimentale et de Microbiologie, 49, 3: 269-282.
Copyright © 2025 - Toate drepturile rezervate