|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
SENESCENTA SI MOARTEA CELULARA PROGRAMATA
Introducere
1. Tipuri de moarte celulara la plante si animale
2. Moartea celulara programata (MCP) in ciclul celular la plante
3. Despre senescenta
3.1. Metabolismul pigmentilor in timpul senescentei
3.2. Metabolismul proteic in senescenta
3.3. Impactul senescentei asupra fotosintezei
3.4. Impactul senescentei asupra metabolismului oxidativ
3.5. Degradarea acizilor nucleici in senescenta
3.6. Reglarea activitatii metabolice in celulele senescente
3.7. Reglatorii endogeni de crestere ai plantei si senescenta
3.8. Influenta mediului in senescenta
4. Exemple de dezvoltare prin MCP la pante
5. Exemple de MCP ca raspuns al plantei la stres
6. Alte perspective de cercetare pentru MCP
INTRODUCERE
Moartea seturilor specifice de celule este, in mod paradoxal, o partea esentiala a dezvoltarii multor eucariote, plante si animale; in plus, moartea celulara poate fi un raspuns la stresul biotic si abiotic. Procesul poarta numele de moarte celulara programata (MCP) datorita faptului ca organismul controleaza initierea si executia procesului.
La plante, exista doua forme de MCP: senescenta si moartea celulara asociata cu raspunsul hipersensibil. Senescenta este moartea celulara relativ lenta, spre sfarsitul limitei de viata, cu dezorganizarea componentelor celulare din tesuturile senescente; procesul permite recuperarea maxima a nutrientilor din tesuturile senescente pentru reciclare si redictionare spre partile din planta care supravietuiesc. Moartea asociata cu hipersensibilitatea este diferita: in acest caz, mai degraba decat a fi recuperati nutrientii, tesutul gazda este omorat pentru ca patogenul sa nu se raspandeasca in planta. Prin urmare, din aceasta perspectiva se poate aprecia ca moartea care se produce cand si unde este necesar in corpul plantei devine o parte esentiala a vietii.
1. TIPURI DE MOARTE CELULARA LA ANIMALE SI PLANTE
Atat la plante, cat si la animale exista mai multe tipuri de moarte celulara programata, incluzand toate procesele care se realizeaza cu consum de energie, sunt reglate de seturi distincte de gene si contribuie la dezvoltarea morfologica si fiziologica a organismului.
La animale, exista doua tipuri majore de moarte celulara: apoptoza, proces inalt reglat, dependent de enegie si necroza, care rezulta in urma leziunilor. Apoptoza implica formarea de vezicule la suprafata externa a membranei celulare si a celei nucleare, condensarea cromatinei, clivajul ADN in fragmente internucleozomale caracteristice de 180 pb si formarea de corpuri apoptotice, ulterior degradate de celulele vecine. Necroza duce la ruperea membranei plasmatice si endomembranelor, avand drept consecinta eliberarea rapida a enzimelor hidrolitice si inflamare. Iata cateva exemple de MCP in embriologia si fiziologia animalelor:
a) in embriologie:
moartea tesuturilor interdigitale in timpul dezvoltarii embrionare, la membrele mamiferelor, ducand la formarea degetelor. La pasari, acest proces este ilustrat de diferenta intre piciorul de gaina si cel de rata;
eliminarea celulelor musculare in metamorfoza insectelor;
resorbtia cozii mormolocului, la amfibieni;
in dezvoltarea la Caenorhabdites elegans, din cele 1090 de celule ale organismului, 131 de celule specifice mor, conform unei scheme precise;
dezvoltarea embrionara normala a creierului si ochilor la soareci
b) in fiziologie:
reglarea fiziologica a tesutului placentar;
raspunsul patologic la mutageni, toxine si hormoni;
eliminarea leucocitelor autoreactive din sistemul imun.
La plante, au fost descrise mai multe tipuri de moarte celulara, unele dintre ele fiind considerate non-apoptotice. Apoptoza este relativ rara rara la plante, deoarece prezenta peretelui celular impiedica formarea de corpuri apoptotice, iar celulele plantelor nu contin fagocite; in plus, condensarea cromatinei nu acompaniaza tipurile de moarte celulara programata la plante.
Prin ce cai, totusi, "scapa" plantele de celulele moarte? O cale este reprezentata de autofagie - similara cu cea observata la drojdii, dupa infometare: celulele produc autofagosomi (vezicule care inglobeaza portiuni din citosol, inclusiv organite intacte), inclusi apoi intr-o vacuola a celulei, unde sunt distrusi de catre enzime hidrolitice. Un tablou foarte similar exista in culturi de celule vegetale supuse infometarii. O alta forma de autofagie are loc in stratul aleuronic al cerealelor, in care numarul de organite este redus: vacuola centrala creste si ocupa intreaga celula, inainte de moartea acesteia. Al treilea tip este exemplificat in diferentierea elementelor traheale: acestea mor la maturitatea functionala, iar degradarea organitelor si a continutului celular are loc prin ruperea tonoplastului si eliberarea de hidrolaze; ca model de studiu in acest tip de reciclare a celulelor moarte este Zinnia.
Unele tipuri de MCP sunt unice la plante. Formarea endospermului central la cereale este un exemplu: timpuriu in formarea semintei, endospermul se diferentiaza in doua tipuri tisulare: un strat continand amidon si un strat exterior cu aleurona. Celulele endospermului amidonos acumleaza amidon si depoziteaza proteine in timpul maturarii semintei; la
maturitate, aceste celule mor, in timp ce celulele aleuronice inconjuratoare raman vii. Celulele moarte isi mentin insa continutul, inclusiv nucleul si organitele, mumificandu-se si nedegradandu-se pana la initierea germinarii si cresterii plantulei, chiar daca aceasta se intampla dupa luni sau ani. La initierea germinarii, enzimele secretate de stratul aleuronic ataca endospermul mumificat.
In sfarsit, un alt tip de MCP, inca incomplet elucidat, este moartea celulara ca raspuns hipersensibil al gazdei la patogeni. Evenimentele, in acest caz, variaza functie de planta si patogen. La modul general, acest tip de moarte celulara include: recunoasterea patogenului, influx de ioni de calciu in citosol, producerea de radicali de oxigen si de oxid nitric si activarea unei proteinkinaze mitogene care controleaza transmiterea semnalului de moarte celulara in cascada.
Celulele plantelor raman viabile aproape in tot timpul programului de dezvoltare care duce la moarte celulara. Pentru a intelege moartea celulara programata, trebuie facuta distinctia intre procesele care duc la moarte - in timpul carora celulele sunt viabile - si moartea propriu-zisa. In timp ce la animale, programul de moarte celulara, odata initiat nu poate fi oprit, la plante, faza care precede moartea poate fi oprita sau chiar reversata.
Senescenta celulelor din mezofil ilustreaza aceasta perioada de viabilitate: aceste celule raman viabile pana cand aproape toate resursele frunzei au fost transportate in alte parti ale plantei. Dupa salvarea plastidelor si a altor organite, frunzele senescente raman turgescente, ceea ce denota integritatea membranelor, a compartimentelor celulare. Pierderea turgescentei urmata de moartea celulara este un eveniment tarziu in programul de senescenta. Moartea propriu-zisa este probabil foarte rapida.
2. MCP IN CICLUL DE VIATA AL PLANTELOR
Moartea celulara programata permite dezvoltarea si plasticitatea biochimica a plantei. MCP intervine in aproape toate fazele ciclului de viata, de la germinare la dezvoltarea vegetativa si reproductiva. In plus, raspunsul la patogeni si stres abiotic implica, de asemenea, moartea celulara controlata (figura 1).
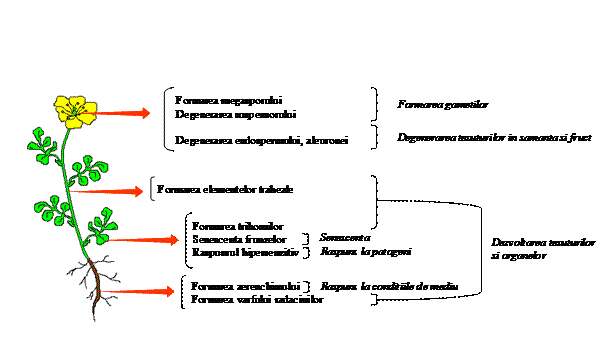
Fig. 1. Moartea celulara programata se petrece in majoritatea tipurilor tisulare vegetale si este implicata in numeroase procese.
MCP este esentiala pentru o dezvoltare reproductiva normala. De exemplu, dezvoltarea florii este afectata radical de MCP prin moartea unor celule sau grupe de celule selectate. La cele mai multe plante cu flori unisexuate, initial, floarea contine primordii atat pentru organe mascule, cat si femele, care in stadiile timpurii nu pot fi distinse; in timpul formarii florii, fie partea mascula, fie cea femela isi inceteaza cresterea si este eliminata prin moarte celulara. Astfel, la porumb, inflorescenta mascula, este spatial separata de inflorescenta femela. Florile tinere (in panicul) contin primordii atat pentru stamine cat si pentru gineceu, dar in dezvoltarea florii, celulele gineceului se opresc din diviziune si crestere, iar organitele si nucleul se fragmenteaza. La mutanta tasselseed2 (ts2), degenerarea gineceului nu are loc si apar flori femele in panicul. Prin urmare, gena TASSELSEED2 (Ts2) este necesara pentru moartea organelor femele din panicul: gena este exprimata in panicul, in celulele gineceului, chiar inainte de inceputul degenerarii acestuia, iar produsul codificat este o enzima similara cu hidroxisteroid dehidrogenaza, ce regleaza moartea celulara prin generarea unei molecule steroidice care actioneaza ca un semnal.
Tesuturile haploide la multe plante sunt reglate prin programe de moarte celulara. In timpul megagametogenezei la angiosperme, 3 din cei 4 megaspori care se formeaza dupa
meioza in celula mama a megasporului sufera MCP, lasand un singur megaspor sa dea nastere la celula-ou si la celelalte componente ale sacului embrionar. MCP joaca un rol si in microsporogeneza, unde tapetumul care inconjoara microsporocitele moare si se dezintegreaza.
MCP intervine in formarea semintei. Dupa fecundare, la cele mai multe angiosperme, prima diviziune mitotica a zigotului da nastere la doua celule: una produce embrionul, cealalta suspensorul. La randul sau, suspensorul parcurge cateva runde de mitoze, dar in final celulele sale sufera MCP.
Dezvoltarea vegetativa este dependenta de MCP. In timpul germinarii semintelor de monocotiledonate si dicotiledonate, endospermul sufera MCP. La cereale, dupa cum s-a mai aratat, endospermul amidonos formeaza un tesut mumificat; in alte seminte, endospermul moare dupa ce functia sa de depozitare sau protectiva s-a terminat. Astfel, la specii precum Arabidopsis, endospermul este o structura formata din doua straturi care inconjoara embrionul, cu functia de a pastra rezervele de proteine, lipide si carbohidrati; in plus, din cauza structurii unice a peretelui celular, acest tesut functioneaza si in controlul cresterii embrionului: inmuierea endospermului in timpul catorva ore de imbibare permite embrionului sa creasca, dupa care MCP duce la distrugerea sa completa prin autoliza.
MCP joaca un rol esential in primele stadii ale diferentierii tesutului mai ales in dezvoltarea xilemului. Formarea elementelor traheale mature cere ca citoplasma elementelor traheale initiale sa fie indepartata incat sa poata circula apa.
In afara de toate acestea, MCP are loc in momente diferite, specie-specifice, ale formarii organismelor. De exemplu, la Monstera, frunzele trec prin schimbari dramatice, pana ating forma specifica. De asemenea, in dezvoltarea vegetativa la cactusi (exemplu extrem, la care tulpina verde functioneaza ca frunze, iar frunzele sunt reduse la spini) multi peri, tepi, spini de pe suprafata tulpinii mor la maturitate. Glandele uleioase de la suprafata fructelor de citrice se formeaza prin moartea celulara a unui grup de celule subepidermale, formand cavitati care se umplu cu uleiuri esentiale; fenomenul se numeste lizigenie si este responsabil pentru diferentierea ductelor secretoare, cavitati sau canale la multe alte specii, intre care si Tilia cordata.
MCP este si componenta a raspunsului unor plante la stres. Un alt exemplu de lizigenie este formarea aerenchimului in radacinile plantelor supuse la stres hipoxic;
caracteristica unica a aerenchimului este ca peretele celular si protoplastul sunt indepartate din radacinile mature, formand canale prin care aerul poate trece din tulpina in radacina. Un alt exemplu de MCP ca raspuns la stres este raspunsul gazdei la o infectie cu patogen.raspunsul gazdei la o infectie cu patogen. Daca gazda recunoaste patogenul, raspunsul sau este moartea rapida a celulelor din imediata apropiere a ariei infectiei, avand drept consecinta moartea patogenului si, implicit, impiedicarea infectarii altor tesuturi.
3. DESPRE SENESCENTA
Senescenta este un proces de moarte celulara programata asociat cu stadiul terminal al dezvoltarii atat in organele vegetative, cat si in cele reproducatoare. Senescenta, si apoi moartea sunt fazele terminale in dezvoltarea tuturor organelor plantei (radacina, tulpina, frunze, flori). Senescenta remodeleaza forma plantei prin inlaturarea tesuturilor nedorite furnizand, in acelasi timp, azot si fosfor. Senescenta are loc dupa maturarea organelor si poate fi influentata dramatic de mediu si perturbari hormonale. Uneori senescenta este rapida, alteori poate dura ani; de exemplu, la Ipomea tricolor florile dureaza doar o zi, pe cand la Pinus longaeva, frunzele au o limita de viata de 45 de ani, iar singura frunza a plantei de desert Welwitschia mirabilis supravietuieste zeci de ani.
Nu se cunoaste exact cum decurge programul senescentei. Se presupune ca este prezent permanent un semnal de tipul "mori acum", iar tesutul sau organul "stie" cand sa raspunda la acest semnal. Mutatiile de tipul ramai verde (stay green), ca si necrozaca si necroza implica gene care raspund de reglarea desfasurarii sau localizarii senescentei normale si a MCP. Studiind aceste gene s-a constatat ca moartea celulara selectiva duce la diferentierea plantei.
Celulele care imbatranesc sufera reorganizare interna si sunt active metabolic. Senescenta este un proces reglat genetic, ce decurge in cateva etape, in care unele cai metabolice sunt activate, iar altele inhibate. De exemplu, in senescenta frunzei, evenimentele in cascada sunt declansate de stimuli din mediu sau hormonali, care duc la activarea sau inactivarea unor gene; in urmatoarea etapa are loc rediferentierea controlata a structurilor celulare si remobilizarea "materialelor de constructie"; in etapa terminala pot fi recunoscute
multe dintre caracteristicile MCP. Integritatea membranelor subcelulare si compartimentarea cailor biochimice sunt pastrate pana tarziu, in faza terminala.
Initiatorii Acceleratorii Inhibitorii
senescentei senescentei senescentei


 hormoni hormoni
hormoni
hormoni hormoni
hormoni
mediu mediu mediu
dezvoltare dezvoltare dezvoltare


 patogeni
patogeni


Transmiterea Activare sau
![]()
![]()
 semnalului inactivare genica
semnalului inactivare genica
Moarte celulara
Fig. 2. Mecanisme in senescenta frunzei
Senescenta frunzei si fructului sunt
caracterizate prin schimbari dramatice in organite, in particular in plastidele
mezofilului si parenchimului,
pericarpului fructului. De asemenea, cloroplastele mezofilului frunzei si ale
fructelor imature verzi rediferentiaza in gerontoplaste si, respectiv,
cromoplaste. In timpul MCP In timpul MCP, vacuolele care contin rezerve din
celulele cotiledoanelor si endospermului semintelor se schimba din organite de
depozit in vacuole centrale care mentin integritatea tonoplastului. Organitele
specializate continand lipide, denumite oleosomi, din cotiledoane si endosperm
sunt pierdute in timpul senescentei atat la mono- cat si la dicotiledonate. In
senescenta, tesuturile fotosintetizatoare si peroxizomii sunt concentrati in
organite noi numite glioxizomi. Prin urmare, senescenta nu este o simpla forma de necroza, ci un proces
programat. Aceste schimbari ultrastructurale in senescenta sunt acompaniate
de schimbari biochimice - degradarea clorofilei si depozitului de proteine.
Senescenta presupune si salvarea si redistribuirea metabolitilor, in mod special a rezervelor de azot si fosfor. Declinul fotosintezei implicit senescentei inseamna surplus de energie de remetabolizare, modificari ale metabolismului oxidativ si ale respiratiei.
Procesele implicate in senescenta se desfasoara in mai multe etape:
Etapa de initiere: alterarea metabolismului si a proceselor redox, activarea unor cai de semnalizare;
Etapa de reorganizare: activarea cailor de salvgardare, trecerea de la metabolism autotrof la heterotrof, detoxifiere, rediferentiere reversibila a organitelor;
Etapa terminala: acumulare de substante antibiotice substante antibiotice, eliberarea de radicali liberi, eliminarea metabolitilor ramasi, pierderea ireversibila a integritatii si a viabilitatii celulare.
Au fost izolate gene exprimate in timpul senescentei frunzei de Arabidopsis, Brassica, porumb, tomate, orz, Lolium. Genele asociate cu coacerea fructelor la tomate, banane si pepene includ omologi ai genelor asociate cu senescenta. Similar, omologi ai genelor asociate cu senescenta frunzei sunt exprimate si in senescenta unei parti a florii, a fructului si a semintelor care germineaza. Nomenclatura genelor asociate cu senescenta variaza de la specie la specie si in relatie cu descoperitorul lor: See la porumb, LSC la Brassica napus, SAG la Arabidopsis etc. S-au identificat, pe baza omologiei de secventa, cca. 50 de ADNc cu functii potentiale in senescenta: preponderente sunt genele care codifica enzime proteolitice, dar exista si gene impotriva organismelor patogene, si gene de raspuns la stres.
Pana in prezent nu a fost elaborat un model simplu de reglare a expresiei genice in timpul senescentei. Analiza structurii si functiilor promotorilor diferitelor gene asociate cu senescenta arata ca aceste gene difera mult intre ele. In plus, diferite studii au identificat mutante senescente - plante defective in activivtati enzimatice specifice senescentei sau activitati implicate in desfasurarea senescentei.plante defective in activitati enzimatice specifice senescentei sau activitati implicate in desfasurarea senescentei
Senescenta per se este un subiect relativ recent in genetica plantelor, cu toate ca senescenta cotiledonului a fost unul dintre cele sapte caractere studiate de Mendel. De exemplu, la mazare, cotiledoanele sunt verzi, asemanatoare frunzelor; alela recisiva i determina cotiledoane verzi, iar homozigotarea sa, ii duce la intarzierea ingalbenirii in senescenta; aceste plante homozigote sunt deficiente pentru enzima care degradeaza clorofila. La Phaseolus vulgaris var. flageolata exista, de asemenea, cotiledoane verzi, care nu ingalbenesc, dar mecanismul pare a fi diferit de blocajul catabolismului clorofilei. La soia au
fost descrisi in jur de 10 loci cu efecte directe, indirecte sau interactive asupra senescentei. Variantele genetice ale senescentei intarziate a frunzei, numite mutante stay green, sunt variante genetice cu desfasurare alterata a senescentei, nu cu defecte ale activitatii enzimatice.
Cerealele stay green si alte plante au importanta economica. De exemplu, in Illinois, in l985, a fost realizata o productie record de porumb de 24 t/ha, folosind o varietate de porumb stay green. Desi factorii care au contribuit la aceasta productivitate sunt complecsi, fenotipului stay green i se atribuie o contributie de cca. 30%. In zonele semiaride, caracterul stay green este de importanta vitala pentru populatiile existente, contribuind, de exemplu, la marirea rezistentei sorgului si meiului la boli si insecte, la adaptarea la mediu, maturizare, digestibilitate. Mutatia stay green a locusului sid la specii din genul Lolium si Festuca a fost studiata pentru stabilirea cailor biochimice ale degradarii clorofilei. La aceste specii, folosite ca furaje, mentinerea frunzelor verzi insemna mentinerea unei calitati superioare a proteinelor in frunze. Mentinerea plantei verzi este un caracter cantitativ, determinata de cativa loci identificati la sorg si mei; acestia sunt studiati intens cu ajutorul markerilor moleculari situati langa genele asociate senescentei. Au mai fost descrise pana acum trei mutatii stay green la Arabidopsis si se studiaza in continuare functiile acestor gene.
3.1. Metabolismul pigmentilor in timpul senescentei
Simptomul cel mai evident al senescentei la frunze este pierderea treptata a culorii verzi, datorate conversiei clorofilei la clorofilida, cu ajutorul clorofilazei. Alt produs al activitatii clorofilazei este fitolul, care se acumuleaza in globulele lipidice ale gerontoplastelor, in special sub forma de esteri. In unele cazuri, frunzele verzi devin rosii, ca in cazul lipsei de azot sau a trecerii la heterotrofie. Catabolizarea clorofilei permite observarea carotenoizilor foliari, care se acumuleaza sau se diminueaza in timpul senescentei, in functie de specia de plante. Astfel, la unele specii de copaci cu frunze cazatoare (dar si la alte plante), pierderea clorofilei merge in paralel cu evidentierea carotenoizilor, frunzele copacilor devenind colorate inainte de a cadea, iar fructele - colorate spre coacere; pierderea clorofilei permite, astfel, evidentierea pigmentilor de baza galbeni spre portocalii, ca si pigmentii noi acumulati. Genele pentru trei din enzimele cheie pentru sinteza carotenilor din diferite plante au fost clonate si au fost descrise mai multe mutante de culoare.
Carotenoizii sintetizati in plastidele fructelor in coacere sunt concentrati in structuri diferite (fibrile, cristale sau globule), care devin atat de numeroase, incat cloroplastele rediferentiaza in cromoplaste. Fibrilinele, proteine specifice asociate cu carotenoizii tesutului fructului au fost detectate si in picaturile de lipide care se acumuleaza in plastide in frunzele senescente.
De asemenea, in senescenta poate fi modificat si metabolismul fenilpropanoidului. Produsii de metabolism ai fenilpropanoidului (salicilatul, fenolii, taninurile, flavonoizii si ligninele) se acumuleaza ca pigmenti sau produsi secundari in tesuturile senescente, datorita crestrii ratei de alterare a sintezei produsilor fenolici. De exemplu, activitatea diferitelor enzime din metabolismul fenilpropanoidului se modifica in timpul dezvoltarii fructului capsunului. Studiul reglarii cailor metabolice secundare in senescenta si coacere este un domeniu aflat inca la inceputurile sale.
3.2. Metabolismul proteic in senescenta
Degradarea unor pigmenti in timpul senescentei este acompaniata de mobilizarea proteinelor cloroplastelor. Spre deosebire de mitocondrii, care sunt necesare pentru mentinerea viabilitatii, cloroplastele pot fi eliminate in timpul senescentei. In celulele verzi, mai multe proteine sunt localizate in cloroplaste, incat aceste organite sunt sursa principala de azot organic salvat din tesuturile senescente. Cea mai abundenta proteina este rubisco, localizata in stroma cloroplastului; descresterea cantitatii acestei proteine este legata de progresul senescentei. Un posibil mecanism a fost considerat exportul proteinelor din plastide pentru a fi degradate in alte locatii in celula, cum ar fi vacuola, compartiment celular litic; cu toate acestea, se pare ca primele faze ale proteolizei au loc in cloroplast. Au fost identificate enzimele necesare pentru degradarea proteinelor cloroplastice.
3.3. Impactul senescentei asupra fotosintezei
Rata descrescanda a fotosintezei in frunze si alte tesuturi verzi este simptomul cel mai clar al senescentei. Masurarea schimbului de gaze (fixarea CO2 si eliberarea de O2) este utilizata pentru estimarea diminuarii capacitatii fotosintetice, in paralel cu diminuarea clorofilei sau a continutului proteic.
Impactul senescentei asupra metabolismului oxidativ
Senescenta este corelata cu conversia peroxizomilor (organite care participa in fotorespiratie, la plantele C3) la glioxizomi (care convertesc lipidele in zaharuri). De asemenea, senescenta este sensibila la conditiile redox. Metabolismul oxidativ este esential la nivel celular in senescenta, unele cai de semnalizare reglatoare ale senescentei putand include componente redox. De exemplu, expresia genelor care codifica fibriline asociate cu pigmentii din cloroplaste si gerontoplaste este sensibila la oxigen.
In senescenta, exista multe surse de specii de radicali liberi de oxigen. De exemeplu H2O2 si superoxidul sunt produsi normali in reactiile enzimatice din peroxizomi, glioxizomi, cloroplaste. Enzimele de aparare impotriva speciilor reactive de oxigen sunt catalazele si dismutazele. Modelul de reglare al programului senescentei implica un echilibru intre speciile de oxigen si sistemele antioxidante. Radicalii de oxigen in exces pot distruge celula, pana la omorarea sa.
Degradarea acizilor nucleici in timpul senescentei.
Prin catabolizarea acizilor nucleici se elibereaza fosfat anorganic: fosforul organic din frunze intra in componenta acizilor nucleici care, ca si proteinele, reprezinta salvgardatori de substante de rezerva cu greutate moleculara mare. Fosforul organic din frunze este salvat in tesuturile imbatranite.
O diferenta chimica importanta intre fosforul si azotul din substantele organice din interiorul celulelor este ca fosforul nu se leaga direct la carbon, ci exista sub forma esterificata, care poate fi hidrolizata. Fosfatazele sunt active peste tot, in special in timpul senescentei deci, energetic si enzimatic exista conditii pentru eliberarea fosfatului din compusi organici. Nucleosidele sunt clivate in zaharuri, purine si pirimidine, catabolismul fiind puternic activat de senescenta. In final, prin catabolismul acizilor nucleici se elibereaza fosforul anorganic, care este apoi transportat prin sistemul vascular, sub forma de fosfat anorganic.
In timpul senescentei se intensifica si activitatea unor ribonucleaze. Cantitativ, fractia cea mai mare de acid nucleic care contribuie la remobilizarea fosforului este ARNr din plastide. Ribonucleazele cliveaza rapid ARN-ul in produsi cu masa moleculara mica. In plus, multe RNaze mitocondriale actioneaza extracelular, iar altele in RE sau vacuole. De exemplu
RNazele sunt sintetizate de novo in timpul senescentei florilor efemere, la zorele, iar la Arabidopsis au fost clonate trei RNaze puternic exprimate in frunzele si florile senescente.
Tabloul general al degradarii acizilor nucleici in senescenta seamana cu tabloul mobilizarii proteice: genele care codifica enzime sunt stimulate si creste activitatea hidrolitica; in schimb localizarea enzimelor nu pare sa corespunda cu principalele locuri de degradare. Pe de alta parte, este important sa se diferentieze intre rolul de reciclare al nutrientilor de catre RNaze in senescenta si alte functii ale enzimelor, de exemplu de turnover al ARN, procesarea ARN, lupta impotriva patogenilor si MCP. Uneori expresia crescuta a RNazelor apare mai degraba mai tarziu, in senescenta, ceea ce ar insemna ca este legata de momentul cand inevitabilul se produce - necroza si moartea.
3.6 Reglarea activitatii metabolice in celulele senescente
Exista cateva mecanisme care explica susceptibilitatea diferita a enzimelor la proteoliza. La inceputul senescentei, in celule sunt deja prezente unele mecanisme metabolice de degradare. Intrebarea este cum pot celulele in curs de imbatranire sa mentina integritatea functionala si structurala a unor enzime critice, intr-un mediu celular in care predomina proteoliza si alte procese litice. Cum fac sa supravietuiasca si sa-si indeplineasca rolul metabolic enzimele gluconeogenezei sau ale metabolismului aminoacizilor, in timp ce alte enzime, cum sunt cele ale fixarii carbonului fotosintetic, sunt degradate?
Se considera ca aceasta stabilitate este asigurata prin urmatoarele mecanisme:
a. cand enzimele sunt mai mult sau mai putin expuse la degradare, activarea diferita a expresiei genice poate face ca unele proteine sa fie inlocuite continuu, in timp ce altele sunt indepartate si neinlocuite;
b. compartimentarea: multe enzime citosolice sunt neafectate sau subreglate in timpul senescentei, in timp ce izoforme organitice ale aceleiasi proteine sunt degradate.
3.7. Factorii endogeni de crestere a plantei si senescenta
Senescenta cauzata de factori interni si externi este mediata de interactiuni complexe, inclusiv hormonale, de factori reglatori endogeni. In legatura cu reglatorii de crestere a plantei si rolul lor in senescenta trebuie subliniate doua probleme:
a. intrarea in senescenta depinde de diferite semnale, printre care si reglatorii endogeni de crestere;
b. raspunsul plantelor la reglatorii de crestere specifici, cu privire la senescenta, nu este acelasi in toate plantele. De exemplu, la cele mai multe plante etilena induce senescenta frunzei, pe cand la altele nu are niciun efect.
Etilena actioneaza primar ca promotor al senescentei, fiind cel mai puternic dintre acestia. La multe plante, tratamentul exogen cu etilena induce senescenta frunzelor, florilor si coacerea fructului. De exemplu, tratamentul frunzelor cu etilena la Arabidopsis induce schimbari asemenatoare cu senescenta naturala a frunzelor: frunzele incep sa se ingalbeneasca, scade expresia genelor pentru fotosinteza si creste expresia genelor asociate cu senescenta.
Mai mult, concentratia de etilena se coreleaza cu senescenta frunzelor si a florilor. La petunii, producerea de etilena poate fi detectata in 20 minute de la de polenizare si precede penetrarea tubului polinic in stigmat. Astfel, recunoasterea polenului de catre stigmat pare a fi suficienta pentru a induce producerea de etilena.
Din cauza importantei economice, s-a studiat mult reglarea senescentei de catre etilena in contextul coacerii fructului. In timpul dezvoltarii normale a fructului, la tomate, de exemplu, creste respiratia si producerea de etilena urmata de cresterea expresiei genelor specifice si a activitatilor legate de coacere, ducand la schimbari caracteristice in culoare, textura si parfum. Efectul etilenei exogene ca agent de coacere este cunoscut de mai mult de un secol si expunerea controlata la etilena este inca un factor major la succesul de piata al multor fructe. In concluzie, o crestere in concentratia de etilena coincide cu senescenta la multe plante, si tratamentul exogen cu etilena este declansatorul inceputului senescentei.
Alta dovada a rolului critic al etilenei in senescenta vine din experiente desemnate sa testeze daca detectarea etilenei este necesara pentru senescenta. Mutantele etr1, la Arabidopsis, si never ripe la tomate sunt insensibile la etilena, pentru ca nu au o proteina receptor al etilenei. La aceste plante, senescenta frunzelor si florilor este mult intarziata, iar fructele de tomate nu se coc niciodata complet.
De asemenea, reducerea experimentala a producerii de etilena in plante a confirmat rolul sau in senescenta: prin inginerie genetica s-au obtinut plante care produc mai putina etilena; drept consecinta, fructul nu se coace niciodata complet. Acid 1-aminociclo-propan-1-carboxilic (ACC) sintetaza si ACC oxidaza sunt doua enzime care catalizeaza ultimile etape
in biosinteza etilenei. Plantele modificate genetic care exprima aceste gene in orientare antisens au o concentratie redusa de etilena; cand aceste plante sunt crescute in aer liber, fructul nu se coace niciodata complet; cand sunt tratate exogen cu etilena, parcurg senescenta si se coc. In concluzie, rolul etilenei poate fi diferit in diferite organe ale aceleiasi plante: la mutantele de Arabidopsis si de tomate insensibile la etilena, senescenta frunzelor este doar intarziata, in timp ce fructele nu se coc niciodata. Prin urmare, etilena promoveaza, dar nu este esentiala pentru senescenta frunzelor, in timp ce este absolut necesara pentru coacerea fructului. Aceasta observatie are drept consecinta ipoteza ca etilena nu actioneaza singura pentru promovarea senescentei; tratamentul cu etilena pentru inducerea senescentei este eficient numai daca s-a atins un anumit stadiu de dezvoltare a plantei sau organului respectiv; in reglarea senescentei mai intervin, cu certitudine, si alti factori - inca neidentificati. In plus plantele de tomate manipulate genetic sa supraproduca etilena nu arata niciun fenotip alterat al senescentei frunzei, ceea ce sugereaza inca odata ca etilena singura este nu este suficienta pentru producerea senescentei.
Pe de alta parte, s-a demonstrat ca citokininele actioneaza ca antagonisti ai senescentei. Citokininele par sa joace un rol major in reglarea senescentei, dar cu un efect opus celui al etilenei, si anume de inhibitie a procesului de senescenta, astfel:
a. concentratiile de citokinine endogene reduc senescenta celor mai multe tesuturi;
b. citokininele endogene cauzeaza intarzierea senescentei in multe tesuturi.
Ca si in cazul etilenei, efectul citokininelor variaza cu varsta si tipul tesutului, si de la specie la specie. In prezent, cu ajutorul tehnicilor moleculare se studiaza rolul citokininelor in diferite stadii de dezvoltare, inclusiv in senescenta. S-au produs plante transgenice cu gena de biosinteza a citokininei exprimata in diferite tesuturi, demonstrand ca citokininele pot intarzia senescenta frunzelor. O consecinta practica a prelungirii limitei de viata fotosintetica a frunzelor este cresterea numarului de seminte produse. O posibila explicatie pentru efectele citokininelor de intarziere a senescentei la plante este amplificarea activitatii metabolice. Tesuturile cu continut mai mare de citokinine sunt cele mai puternice situri metabolice, si astfel majoritatea nutrientilor sunt directionate spre ele. Citokininele sunt produse de obicei in radacini si transferate in frunze. O ipoteza care explica senescenta frunzelor dupa inflorire este aceea ca citokininele formate in radacina sunt redirectionate spre seminte; samanta devine astfel un sit metabolic mai puternic, ducand la redirectionarea nutrientilor din frunze in
samanta si imbatranirea frunzei. Aceasta ipoteza nu a fost confirmata, insa, in experiente cu soia, unde concentratia citokininelor scade in timpul formarii semintei in toate tesuturile, inclusiv in samanta; in acest caz, explicatia ar fi ca citokininele probabil actioneaza la doua nivele: la distanta, prin promovarea diferentierii si activitatii metabolice puternice si local, prin represarea programului de senescenta.
Aceasta activitate duala a citokininelor este observata si in regenerare. La Nicotiana rustica, la inflorire, frunzele cele mai de jos, cele mai batrane, sunt galbene; daca se taie tulpina deasupra celui mai de jos nod, si planta se pune la lumina, frunzele redevin verzi. Procesul se accelereaza daca frunzele sunt tratate cu citokinine. Aceasta demonstreaza potenta citokininelor ca factor antisenescent, pe de o parte, dar si faptul ca senescenta este reversibila chiar si intr-un stadiu avansat si, deci, este fundamental distincta de alte procese de MCP.
In afara de citokinine si etilena au mai fost identificati si alti reglatori de crestere, cum sunt: reglatorii ABA, giberelinele, brasinosteroizii si poliaminele, desi rolul lor in senscenta este mai putin inteles.
3.8. Influenta mediului asupra senescentei
Senescenta este o activitate fiziologica, un raspuns la conditiile de mediu sub- sau supraoptime. De exemplu, la copaci, caderea frunzelor este un mod de pregatire pentru iarna. O alta caracterisica a sensibilitatii la mediu a senescentei este modul in care un factor dat determina initierea senescentei, dar efectul diferit pe care acelasi factor il poate avea daca senescenta deja este instalata. De exemplu, terenul mlastinos induce senescenta prematura, dar cantitatea mica de apa in timpul senescentei incetineste ingalbenirea.
4. Exemple de dezvoltare prin MCP la plante.
La plante, doua exemple de dezvoltare prin MCP sunt formarea de elemente traheale si mobilizarea endospermului cerealelor.
Dezvoltarea xilemului este un sistem model folositor pentru explicarea MCP la plante. Dezvoltarea xilemului este initiata in embriogeneza si continua toata viata plantei, noi elemente traheale fiind formate din celule de precambiu si cambiu. Xilemul este la inceput
compus din elemente traheale care, in timpul stadiului final de diferentiere, sufera ingrosari ale peretelui celular, urmate de moartea celulara si autoliza continutului celular. Tot ce ramane din celula este peretele celular cu ingrosari caracteristice. Moartea celulara programata in elementele traheale este asociata si cu liza membranelor vacuolare, iar reglatorii de crestere si calciul sunt implicati, de asemenea. Astfel, auxinele si citokininele sunt necesare pentru diferentierea elementelor traheale, iar calciul are un rol probabil in semnalizarea cailor de diferentiere. S-a demonstrat in cultura in vitro ca ionul de Ca2+ creste in cantitate in celulele care incep lignificarea.
Un al doilea sistem-model de analiza a reglarii MCP la plante, este reprezentat de cele doua tipuri celulare de endosperm, amidonos si cu aleurona. Ambele tipuri celulare sufera MCP, dar in moduri distincte. Endospermul amidonos al semintei mature este mort, dar spre deosebire de aproape toate celulele eucariote care sufera MCP, nu este degradat, ci mai degraba mumifiat. Etilena poate induce MCP in celulele endospermului amidonos. Cand samanta germineaza, sunt secretate enzime hidrolitice din stratul aleuronic si endospermul amidonos este degradat. Celulele stratului aleuronic, in schimb, raman vii in timpul germinarii; moartea celulelor stratului aleuronic are loc treptat in timpul germinarii, prin vacuolizare progresiva, urmata de moartea si colapsul protoplastului. Celulele stratului aleuronic sufera autoliza si mor numai dupa ce rezervele de amidon au fost mobilizate. Acidul giberelinic si ABA (acid abscisic) regleaza acest proces, primul prin stimularea debutului MCP in stratul aleuronic, iar al doilea - prin definitivarea procesului. In celulele stratului aleuronic, degradarea ADN nu duce la formarea de depozite apoptotice - o dovada in plus ca, in acest caz, MCP nu urmeaza calea apoptozei (cand ADN-ul sufera clivaje internucleozomale, generand fragmente de aproximativ 180 pb). Degradarea ADN in celulele aleuronice este un eveniment tarziu in MCP, si incepe dupa ce aproape intreg continutul celulei a fost autolizat; in plus, ADN din celulele aleuronice este degradat in fragmente mici, care nu sunt observate in cazul apoptozei.
5. Exemple de MCP ca raspuns al plantei la stres.
Formarea aerenchimului: este un raspuns la inecare, la limitarea accesului oxigenului la radacini. In aceste circumstante, plantele formeaza un tesut specializat, numit aerenchim, prin MCP la nivelul celulelor corticale ale radacinii localizate intre endoderm si hipoderm. Raspunsul la stresul hipoxic este rapid si duce la indepartarea celulelor corticale ale radacinii, localizate intre endoderm si hipoderm, formand un spatiu care faciliteaza miscarea oxigenului in radacini.
Unul din cele mai bine studiate exemple de MCP la plante este raspunsul de rezistenta al plantei gazda la patogeni (HR - host response). Acest raspuns de rezistenta al plantei la atacul patogenilor este moartea rapida a celulei gazda. HR este programat genetic de plante, este o consecinta a unor noi procese de transcriptie si translatie si este dat de interactii incompatibile intre plantele rezistente la boli si patogeni nevirulenti. Aceste interactii incompatibile sunt controlate de gene de rezistenta (R), care fac ca planta sa recunoasca si sa raspunda la patogeni care poarta gene specifice de avirulenta (Avr). In absenta alelei R la gazda si Avr la patogen, se instaleaza boala pentru ca planta nu recunoaste patogenul si are loc asa-numita reactie de compatibilitate.
Alternativ, HR poate fi o consecinta a mecanismului care omoara atat gazda, cat si celula microbiana. Moartea celulara poate fi si un simptom de boala in timpul interactiilor compatibile sau poate sa nu fie programata la planta, ci poate fi un rezultat provocat de patogen, prin toxinele lui. Totusi, se pare ca moartea celulara asociata cu HR in raspunsul la patogeni este, mai degraba, programata genetic de plante, decat rezultatul omorarii directe de catre patogen. Aceasta, pentru ca HR este un proces activ, care necesita transcriptie si translatie. Moleculele produse de patogen sau ca rezultat al intereactiei patogen-gazda (peptide si oligozaharide) aplicate la o planta care contine gene de rezistenta induc raspunsuri biochimice si celulare, caracteristice HR, inclusiv moartea celulara. Cand sunt aplicate la o gazda rezistenta, nu produc niciun raspuns. Deci moartea celulara la o gazda rezistenta nu este in mod necesar rezultatul efectelor toxice directe.
In ceea ce priveste mecanismul MCP care rezulta din interactia gazda-patogen, pentru a stabili precis cum mor celulele, s-au examinat celulele plantelor care sufera MCP in interactiunea gazda - patogen, in senescenta si, respectiv, in timpul dezvoltarii plantei. Celulele plantelor au cateva trasee spre moarte - unele unice plantelor, altele asemanatoare cu apoptoza. Exista din ce in ce mai multe dovezi ca, la plante, raspunsul celulelor la patogen
este moartea de tip apoptoza. Criteriul general de definire a apoptozei la animale include formarea de corpuri apoptotice, formarea de vezicule din membrana nucleara, condensarea cromatinei si clivajul ADN-ului nuclear in fragmente internucleozomale. Clivajul ordonat al ADN in fragmente de 50 kb a fost demonstrat in doua sisteme diferite gazda-patogen, iar alte caracteristici morfologice ale apoptozei au fost raportate in alte sisteme gazda-patogen. Cu toate acestea, desi exista unele paralele functionale intre HR la plante si apoptoza, la nivel molecular nu s-au dovedit asemanari - in sensul ca desi reglatori ai mortii celulare observati in sistemele animale au fost gasiti si la plante, nu au fost gasite inca izolate genele corespunzatoare la plante. Aceasta ar putea indica ca genele corespunzatoare plantelor s-au diversificat intr-atat incat nu mai sunt omoloage celor de la animale, dar functiile lor au fost conservate sau ca plantele au dezvoltat alte cai de control ale HR. Similar, genele care regleaza anumite forme de moarte celulara clonate la plante nu au corespondent in sistemele animale. Toate aceste dovezi arata ca, pe de o parte plantele si-au dezvoltat cai proprii de reglare a proceselor de moarte celulara care difera de cele de la animale, iar pe de alta parte, ca au conservat cai de reglaj ale proceselor de moarte celulara prezente si la animale.
La plante, diferitele forme de MCP sunt relativ similare la nivel de mecanisme, mai ales in ultima faza, cea de executie. Senescenta este relativ inceata si ordonata, maximizeaza recuperarea metabolitilor din tesut si poate fi reversibila pana la un stadiu tarziu. Moartea celulara asociata cu HR, pe de alta parte, minimalizeaza oportunitatile patogenului. Studii asupra expresiei genelor in diferite forme de MCP vor aduce noi indicii asupra cailor comune de desfasurare a acestui proces.
Concluzii
Moartea selectiva a celulelor, tesuturilor si organelor este o caracteristica a dezvoltarii si supravietuirii plantelor. Fenomenul de moarte celulara programata se refera la orice proces prin care protoplastul, cu sau fara perete celular, este eliminat, ca parte a unui eveniment de dezvoltare celulara sau de adaptare in ciclul de viata al plantei. Mecanismele prin care plantele fac acest lucru sunt: autofagia, lizigenia, mumificarea. MCP este esentiala pentru dezvoltarea reproductiva si vegetativa normala si pentru raspunsul la stresurile din mediu.
In timpul senescentei (proces asociat cu stadiile terminale ale dezvoltarii organului) sunt exprimate gene specifice, unele deja clonate si promotorii lor analizati. Mutantele stay green sunt variante genetice in care expresia genelor asociate cu senescenta este impiedicata.
Celulele senescente sunt metabolic active. Calea de degradare a clorofilei este deturnata si acompaniata de acumularea de carotenoizi sau alti pigmenti. Proteinele sunt degradate, iar azotul si fosforul organic sunt exportate din frunzele senescente. Prin catabolismul acizilor nucleici se elibereaza fosforul anorganic. Fotosinteza este in declin si peroxizomii sunt rediferentiati in glioxizomi care convertesc lipidele in zaharuri. Reglarea metabolica in timpul senescentei implica raspunsuri la conditiile redox celulare, compartimentalizarea si susceptibilitatea diferentiata a enzimelor la proteoliza. Senescenta este sensibila la reglatorii de crestere, in special la promotorul senescentei, etilena, si la citokinine, care activeaza ca antagonisti ai senescentei.
Senescenta este folosita atat ca parte a strategiilor adaptative pentru asigurarea supravietuirii sezoniere, cat si pentru lupta impotriva stresului. Formarea elementelor traheale si mobilizarea endospermului cerealelor sunt exemple de desfasurare a MCP. Celulele mezofilului de Zinnia, induse sa formeze elemente traheale in cultura sunt un model de moarte celulara. Cai diferite duc la moartea endospermului amidonos si celulelor aleuronice la cereale: etilena poate induce MCP in celulele endospermului amidonos, iar acidul giberelic si ABA regleaza moartea celulara in stratul aleuronic. Un exemplu de MCP in conditii de stres abiotic este formarea aerenchimului, ca raspuns la conditii de oxigen limitat, atunci cand radacinile sunt inundate. Etilena mediaza formarea aerenchimului, iar semnalizarea intre hipoxie si producerea de etilena este mediata de calciu.
Raspunsul de hipersensibilitate al gazdei este un exemplu de MCP raportat la stresul abiotic si are propriul efect asupra naturii programate a mortii celulare. Mutantele care mimeaza leziunea produsa de agenti patogeni sunt modele utile pentru analiza HR. La Arabidopsis aceste mutatii sunt de doua feluri: care dau raspunsul de aparare si care nu-l dau. Aceste mutante, mai intai afecteaza moartea celulara in contextul absentei raspunsului la patogen, neavand nici un alt efect asupra altor evenimente in desfasurarea mortii celulare. Radicalii liberi de oxigen sunt cheia pornirii MCP care acompaniaza HR. HR poate fi comparat cu apoptoza, tip de MCP la animale care a fost intensiv cercetat si a devenit un model general pentru MCP si reglarea sa. Directiile generale pentru cercetarea MCP includ relatii intre procesul mortii celulare programate si imbatranirea intregului organism.
Copyright © 2025 - Toate drepturile rezervate