|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
FIBRELE NERVOASE
In
tesutul nervos, termenul de fibra
nervoasa desemneaza, in ansamblu, prelungirile citoplasmatice
ale neuronului. Avand insa in vedere faptul ca axonii sunt prelungiri
mai lungi,
in marea majoritate a cazurilor organizate grupat, acest termen este utilizat
cu precadere pentru axon.
Exceptie evidenta fac dendritele
neuronilor pseudounipolari din ganglionii cerebrospinali, care se
distribuie spre periferie si constituie cai senzoriale.
Fibrele nervoase pot fi inconjurate de o serie de teci concentrice, in raport de prezenta sau absenta carora se descriu:
fibre nervoase mielinizate (mielinice), caracterizate prin existenta tecii de mielina (material complex lipoproteic) si a tecii Schwann, respectiv a tecii oligodendrocitare; sunt prezente in substanta alba a sistemului nervos central si in sistemul nervos periferic (cel mai frecvent tip existent, in special in nervii periferici micsti);
fibre nervoase amielinizate (amielinice), lipsite de teaca de mielina, dar avand teaca Schwann; sunt prezente in sistemul nervos periferic;
fibre nervoase nude, lipsite de teaca de mielina si de teaca Schwann, fiind astfel formate numai din cilindrax; in numar mic, provin de la neuroni de asociatie cu axon scurt si sunt intalnite la nivelul extremitatilor terminale ale ramificatiilor axonale sau in substanta cenusie a sistemului nervos central.
FIBRA NERVOASA MIELINIZATA
Fibrele nervoase mielinizate se caracterizeaza prin faptul ca poseda o teaca de mielina. Responsabile de organizarea acestei teci sunt anumite celule de sustinere ale tesutului nervos: celulele Schwann, in sistemul nervos periferic, si oligodendrocitele, in sistemul nervos central.
CARACTERISTICI IN CADRUL SISTEMULUI NERVOS PERIFERIC
Pe masura cresterii in lungime a axonului, celulele Schwann, dezvoltate din linia spongioblastica a crestelor neurale, migreaza de-a lungul acestuia, realizand un invelis celular pana in imediata vecinatate a extremitatii terminale.
|
Observatie |
Celula Schwann este aplatizata, de dimensiuni medii, prezentand peste membrana plasmatica un invelis de tip membrana bazala.
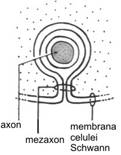
Nucleii apar in sectiune alungiti, iar citoplasma contine organite celulare in numar redus (aparat Golgi mic si cateva mitocondrii). Fiecare celula Schwann are proprietatea de a gazdui, intr-o depresiune a sa, un axon. Marginile acestei depresiuni sunt foarte apropiate, fara insa a fuziona intre ele, formand ceea ce a fost denumit un mezaxon. Aici celula vine in contact cu ea insasi, prin apozitia membranelor plasmatice externe, iar mezaxonul se inchide.
Intre membrana plasmatica a celulei Schwann si axolema exista un spatiu real foarte mic (15 nm) care, prin intermediul spatiului mezaxonic (intre cele doua margini ale depresiunii), comunica in mod liber cu fluidul intercelular. De-a lungul unei fibre se gasesc numeroase celule Schwann care realizeaza astfel, in ansamblul lor, un jgheab pentru fibra nervoasa.
Celulele Schwann nu prezinta jonctiuni intercelulare, dar prelungirile lor citoplasmatice se interdigiteaza.
Teaca de mielina se formeaza prin infasurarea, in multiple spirale (pana la 50), a membranelor celulare ale celulelor Schwann in jurul axonului. Altfel spus, se poate considera ca mezaxonul se alungeste si se ruleaza.
Maniera de infasurare este particulara si consta intr-o suprapunere a membranelor celulare, in straturi concentrice. Corpul celulelor Schwann ramane atasat pe suprafata tecii de mielina. Ansamblul celulelor Schwann asociate tecii de mielina poarta denumirea de teaca Schwann sau neurilema.
Teaca de mielina si teaca Schwann realizeaza un invelis izolator pentru axon, care este protejat astfel de interferenta directa cu mediul extern. Formarea tecii de mielina va fi detaliata in subsectiunea 1.1.
|
|
|
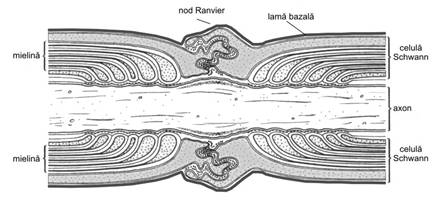
Fig. IV.1. Teaca de mielina |
|
|
|
Fig. IV. Teaca de mielina - sectiune longitudinala |
Aspecte histologice si biochimice
Pe preparatele histologice de rutina, spatiul ocupat de mielina (intre axolema si corpul celulei Schwann) apare ca o zona aproape goala, in care se poate uneori distinge o foarte discreta retea formata din neurokeratina.
Pe preparate realizate prin congelare si tratate cu coloranti pentru lipide, ca si dupa tratare cu saruri de osmiu, teaca de mielina apare intens colorata si omogena (fig. IV.1).
Longitudinal (fig. IV.2), teaca de mielina nu
formeaza un manson continuu, ci este alcatuita dintr-o
succesiune de mansoane cu dimensiuni intre
0,3-1,5 mm; grosimea mansoanelor se coreleaza cu lungimea acestora
(0,08-0,1 mm). Zona care separa doua mansoane vecine
formeaza nodurile Ranvier, iar mansoanele de
mielina dintre doua noduri sunt numite segmente internodale.
Nodurile Ranvier se succed la intervale regulate, iar la nivelul lor axonul
este expus, teaca de mielina fiind absenta. O singura
celula Schwann este responsabila pentru elaborarea mielinei unui
segment internodal. Astfel, pentru fiecare segment internodal in parte se
gaseste o celula Schwann, a carui nucleu apare alungit,
fuziform,
intr-o citoplasma relativ redusa, la suprafata sa.
Prin tehnici speciale, in fibrele nervoase periferice, de-a lungul unui segment, pe toata grosimea tecii de mielina, se identifica discrete depresiuni sau linii oblice, in forma de con; acestea au fost numite incizuri Schmidt-Lantermann, iar numarul lor este direct dependent de diametrul axonului. Microscopia electronica a relevat faptul ca ele corespund zonelor in care citoplasma celulelor Schwann este prinsa si retinuta in interiorul lamelelor de mielina. La acest nivel, citoplasma celulelor Schwann contine lizozomi, uneori mitocondrii, microtubuli si incluzii (corpi densi).
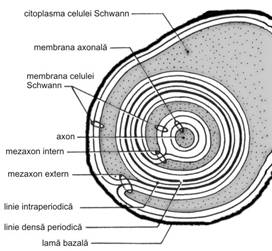
Examinata in difractie cu raze X, in sectiune transversala, teaca de mielina apare formata dintr-o suita de straturi concentrice suprapuse. Aceasta structura a fost confirmata de microscopia electronica, in care apare evidenta o organizare
|
|
|
|
|
|
|
|
|
Fig.
IV.3. Formarea tecii de mielina |
concentrica
de linii (benzi) electron-dense,
in alternanta cu linii (benzi) electron-clare, cu o repetitivitate (periodicitate) de 12 nm. Banda
electron-densa formeaza linia
densa majora
(3 nm). Banda electron-clara, numita linia intraperiodica, este
mai lata si impartita in doua
jumatati egale, printr-o fina linie opaca - linia densa minora
(pe unele preparate poate lipsi).
Cercetari
biochimice au pus in evidenta natura lipoproteica complexa a mielinei. S-au evidentiat: colesterol, fosfolipide (fosfatidil-colina, serina, etanolamina), glicolipide (galactocerebrozide,
sulfatide) si proteine -
proteolipide si proteine bazice mielinice. Dintre proteinele identificate,
P0 - glicoproteina din superfamilia imunoglobulinelor -
prezinta un domeniu extracelular cu rol de adeziune si un domeniu
intracelular, cu rol in fuziunea fetelor interne, iar proteina P2
este localizata la nivelul incizurilor Schmidt-Lantermann si in
zonele paranodale, avand un rol predominant metabolic.
Desi structura biochimica a mielinei sugereaza similitudini cu structura membranelor celulare, exista unele diferente intre mielina din sistemul nervos periferic si cea din sistemul nervos central.
Formarea tecii de mielina
Mielina trebuie interpretata ca fiind rezultatul dispunerii, in jurul unei fibre nervoase, a unui lambou format din membrane ale celulei Schwann, dintre care citoplasma se elimina progresiv, pana cand fetele interne ale membranelor plasmatice se ating si fuzioneaza (fig. IV.3). Procesul este comparat cu ceea ce se intampla la un tub de pasta de dinti - prin eliminarea continutului din portiunea terminala a acestuia, fetele interne ale tubului vin, in final, in contact. Fuziunea fetelor interne ale membranei plasmatice genereaza linia densa majora, iar fuziunea fetelor externe - linia intraperiodica.
Prin
microscopia electronica de inalta rezolutie, s-a observat
insa ca intre fetele externe nu exista o acolare
completa; ele raman la distanta de circa
1-2 nm, realizandu-se astfel un compartiment
extracelular intramielinic, numit deschizatura (sant,
gaura) intraperiodica. Acest compartiment, continuu de la nivelul
spatiului periaxonal la suprafata tecii de mielina,
prezinta o multitudine de complexe jonctionale care il separa de
spatiul extracelular general, accesul spre axon fiind permis numai pentru
molecule mici.
Trebuie mentionat faptul ca, la capatul celulei Schwann care, inrulandu-se, ajunge in proximitatea axonului, se mentine un inel fin de citoplasma, numit colier intern, iar conexiunea dintre fetele externe ale membranelor plasmatice realizeaza mezaxonul intern (locul de contact al capatului terminal interior al celulei Schwann cu ea insasi).
Spatiul periaxonal (15-20 nm) este astfel in continuitate morfologica, prin intermediul mezaxonului intern, cu lamelele profunde ale tecii de mielina. Intr-o maniera similara, un alt inel fin de citoplasma este prezent la exteriorul tecii de mielina, reprezentand citoplasma celulei Schwann de la nivelul celuilalt capat terminal, care nu se inruleaza. Acest inel citoplasmatic este denumit colier extern. Conexiunea stabilita la acest nivel intre fetele externe ale membranelor plasmatice constituie mezaxonul extern (locul de contact al capatului terminal exterior al celulei Schwann cu ea insasi). Lamela cea mai externa a tecii de mielina este in continuitate morfologica, prin intermediul mezaxonului extern, cu membrana plasmatica a celulei Schwann.
In afara zonelor de colier intern si extern, exista posibilitatea ca citoplasma celulei Schwann sa mai persiste intre lamelele succesive de mielina, sub forma de mici insule denumite incizurile Schmidt-Lantermann, considerate defecte de fuzionare a membranelor plasmatice in procesul de inrulare. De asemenea, citoplasma se mentine in zonele perinodale, la nivelul nodurilor Ranvier. Toate aceste zone, confirmate prin microscopie electronica si traduse prin disociatii ale liniilor dense majore, permit o comunicare intre corpul celulei Schwann si lamboul mielinic format.
In sectiune longitudinala peste straturile de mielina, ca si in cazul fibrelor nervoase amielinizate, se observa ca celulele Schwann prezinta, la nivelul nodurilor Ranvier, o multitudine de expansiuni citoplasmatice ce se interdigiteaza cu expansiuni similare ale celulelor vecine (fig. IV.2).
Date din literatura de specialitate sustin existenta unor jonctiuni de tip distantat intre lamele de mielina din zona paranodala apartinand unui segment.
Astfel de jonctiuni nu s-au constatat intre componentele mielinice a doua segmente vecine.
Celula Schwann este invelita intr-o membrana bazala care, dupa ce acopera toate expansiunile citoplasmatice de la nivelul nodului Ranvier, fuzioneaza cu membrana bazala a celulelor Schwann vecine.
Consecutiv, de-a lungul unei fibre mielinizate, axolema este complet izolata de mediul extracelular pe distanta unui segment internodal.
Singura zona de comunicare cu mediul interstitial este nodul Ranvier, la nivelul caruia fenomenele de difuziune se realizeaza prin intermediul membranei bazale. De altfel, aceasta zona are o slaba rezistenta electrica, aici fiind concentrate canale de sodiu, legate de citoschelet prin filamente de ancrina.
Nodul Ranvier constituie deci zona capabila si responsabila de declansarea unui potential de actiune prin depolarizarea membranei.
|
Observatie numarul de ture/spirale creste prin miscari de rotatie ale celulei Schwann in jurul axonului; rotirea axonului are drept urmare tractiunea asupra lamboului in formare. Nici una dintre cele doua ipoteze nu este pe deplin acceptata. Mai mult, in literatura de specialitate se afirma ca procesul de mielinizare este neuniform in timpul dezvoltarii embrionare, existand diferente considerabile in raport cu localizarea, si, respectiv, functia anumitor arii ale sistemului nervos. Astfel, la nastere, mielinizarea este aproape incheiata pentru nervii motori, dar continua dupa nastere, cateva luni pentru nervii senzitivi si cativa ani pentru axonii comisurali si alte traiecte nervoase ale sistemului nervos central. |
CARACTERISTICI IN CADRUL SISTEMULUI NERVOS CENTRAL
In sistemul nervos central, celula mielinizanta este oligodendrocitul, care prezinta cateva prelungiri citoplasmatice.
Fiecare din aceste prelungiri genereaza un segment internodal pe o singura fibra nervoasa sau pe fibre nervoase diferite, astfel incat rotatia independenta a oligodendrocitului este imposibila. Corpul oligodendrocitului, mentinand citoplasma si nucleu, este localizat in vecinatatea fibrelor nervoase mielinizate prin prelungiri, la o oarece distanta de acestea.
Spre deosebire de celulele Schwann, oligodendrocitul nu este tapetat de o lamina externa.
Mielinizarea incepe in perioada de alungire a fibrei nervoase si nu poate fi acceptata ideea de rasucire desfasurata in paralel cu alungirea. Spre deosebire de sistemul nervos periferic, procesul de mielinizare din sistemul nervos central nu este pe deplin elucidat.
In prezent, se accepta, in general, ca este vorba despre activitati sincrone desfasurate in paralel in celula mielinizanta (oligodendrocitul) si fibra nervoasa, care realizeaza interactiuni reciproce. Date experimentale in vivo si in vitro, au permis sa se intrevada:
fenomene de recunoastere reciproca, in stadiul initial, intre celula mielinizanta si fibra nervoasa;
interactiuni reciproce prin semnale moleculare, responsabile, pe de o parte, pentru inducerea dimensiunilor si orientarii fibrei nervoase si, pe de alta parte, de stimularea activitatilor de sinteza ale celulelor mielinizante (pentru molecule de structura).
Astfel, dupa gazduirea unei fibre nervoase in santul creat intr-o prelungire citoplasmatica, aceasta se alungeste progresiv si se infasoara concentric in jurul axonului, din exterior spre interior, excluzand progresiv citoplasma.
Se creeaza un lambou membranar, iar miscarea de inrulare a prelungirii oligodendrocitului (in alungire progresiva) este considerata centripeta, spre deosebire de miscarea realizata de celula Schwann - centrifuga.
Intre teaca de mielina din sistemul nervos periferic si sistemul nervos central exista unele diferente:
in sistemul nervos central, nodurile Ranvier sunt mai mari iar segmentele internodale mai scurte; acest fapt determina o conducere saltatorie mai rapida;
incizurile Schmidt-Lantermann nu se observa in sistemul nervos central, deoarece citoplasma persista numai la nivelul colierului intern si in zona paranodala;
in sistemul nervos central, datorita absentei laminei externe oligodendrocitare si distantarii corpului celular de fibra/fibrele nervoase mielinizata/mielinizate, la nivelul tecii de mielina, stratul situat cel mai superficial este complet lipsit de citoplasma; consecutiv, in cazul in care prelungirile nervoase sunt foarte apropiate, ele vin in contact direct prin tecile de mielina care fuzioneaza, la nivelul unei linii intraperiodice care devine comuna;
componentele proteice din mielina sistemului nervos central nu sunt identice cu cele din mielina sistemului nervos periferic, determinand o alta periodicitate; proteolipidele apar ca molecule transmembranare si sunt reprezentate de PLP si DM20, care asigura coeziunea fetelor externe ale membranei celulare (linia intraperiodica); proteina bazica mielinica determina fuziunea fetelor interne ale membranelor celulare si formarea liniei dense majore; glicoproteinele existente apartin superfamiliei imunoglobulinelor si joaca rol de molecule de adeziune si de recunoastere, avand domenii extracelulare lungi.
ROLUL TECII DE MIELINA
Teaca de mielina are, in primul rand, un rol de izolator electric, impiedicand schimburi ionice intre axoplasma si mediul extracelular. Schimburile se desfasoara numai la nivelul nodurilor Ranvier, determinand depolarizarea electrica a membranei si generarea de potential de actiune. Acest potential va fi transmis prin axoplasma si prin exteriorul tecii de mielina, pana la urmatorul nod Ranvier, realizand astfel conducerea saltatorie cu o mai mare viteza. Excitatia fiind restransa la nivelul nodurilor Ranvier, consumul energetic pentru propagare este mai redus. S-a demonstrat ca viteza de conducere este proportionala cu diametrul fibrei nervoase. Astfel, s-a observat ca pentru a atinge o viteza a influxului de 100 m/sec, realizata de o fibra mielinizata cu diametrul de 20 μm, o fibra amielinizata ar trebui sa aiba un diametru de mai multi centimetri.
FIBRA NERVOASA AMIELINIZATA
Fibrele nervoase amielinizate se caracterizeaza prin existenta, de-a lungul lor, a unei teci celulare alcatuita din celule Schwann, teaca de mielina fiind absenta. Denumite si fibre Remack, sunt prezente la nivelul sistemului nervos periferic, unde formeaza fibrele nervoase simpatice postganglionare.
Absenta tecii de mielina se explica prin capacitatea celulei Schwann de a adaposti, in corpul ei, mai multe fibre nervoase, in mai multe depresiuni (pana la 20) (fig. IV.4). Fiecare depresiune creata poate contine un singur sau mai multi axoni (fascicule de axoni). Acest fapt face imposibila formarea mielinei in jurul tuturor cilindraxilor. Ca si in cazul fibrelor nervoase mielinizate, depresiunile citoplasmatice comunica prin mezaxon cu mediul extracelular, iar marginile jgheaburilor formate nu fuzioneaza complet, facand posibila comunicarea directa intre axolema si substantele extracelulare.
In organizarea unui nerv, un sirag de celule Schwann poate sa realizeze un numar extrem de mare de depresiuni, gazduind astfel mai multe fibre nervoase; in acest caz, este vorba de fibre poliaxonale.
Copyright © 2024 - Toate drepturile rezervate