|







 |  |
|
| |
 | Biologie | Chimie | Didactica | Fizica | Geografie | Informatica |
| Istorie | Literatura | Matematica | Psihologie |
CARACTERISTICILE CELULELOR EPITELIALE
Notiunile prezentate in continuare se refera la caracteristicile generale morfo-functionale enuntate anterior, particularizand pentru tesutul epitelial de tapetare. Notam insa faptul ca, in general, respectivele caracteristici se regasesc si in cadrul celorlalte doua varietati de tesut epitelial, secretor (glandular) si senzorial.
DENSITATEA CELULARA
Tesuturile epiteliale sunt formate dintr-o abundenta de celule, existand foarte putin material intercelular. De aceea, pe sectiunile histologice, un tesut epitelial se recunoaste si dupa nucleii celulelor - care apar foarte numerosi si apropiati intre ei, limitele celulare fiind dificil de distins (avand in vedere ca membrana plasmatica are o grosime sub limita de rezolutie a microscopului optic). Aceasta caracteristica este foarte bine exprimata la epiteliile de suprafata ce acopera exteriorul corpului, invelesc unele organe sau tapeteaza cavitati ale organelor sau cavitati naturale - exceptie facand numai cele ale articulatiilor mobile.
JONCTIUNILE INTERCELULARE
Jonctiuni multiple si variate asigura coeziunea, adezivitatea si comunicarea intercelulara, permitand o functionare coordonata. Se realizeaza astfel o juxtapozitie celulara stransa, importanta in mentinerea integritatii histoarhitectonice, in special la epiteliile de tapetare.
Desi ultrastructura si organizarea moleculara este diferita, jonctiunile intercelulare pot fi sistematizate in trei categorii:
jonctiuni ocludente sau stranse (lat. zonulae occludentes);
jonctiuni
de ancorare sau aderente
(lat. zonulae adherentes, maculae
adherentes);
jonctiuni
comunicante sau distantate
(eng. gap, nexus).
In situatia in care un tip de jonctiune se intinde ca o centura sau ca un cadran pe toata suprafata celulei, la un anumit nivel, ea poarta denumirea de zonula. In cazul in care jonctiunea apare numai din loc in loc, sub forma discontinua, ea poarta denumirea de macula.
Jonctiuni ocludente
Zonula ocludenta
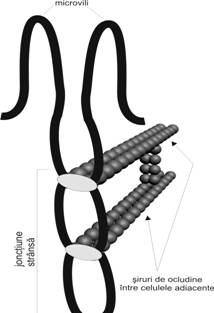
Zonula ocludenta (fig. I.1.1) apare in microscopia electronica ca o apozitie stransa a membranei plasmatice a doua celule vecine.
Foitele
externe ale celor doua plasmaleme fuzioneaza, apoi se desprind,
pentru ca ulterior aceasta secventa de fuzionare-desprindere
sa se repete de mai multe ori, pe o intindere de
0,1-0,3 μm. La locurile de fuzionare, fasii (lanturi) de
proteine transmembranare jonctionale provenite din cele doua membrane
se leaga unele de altele, rezultand un "sigiliu" care inchide spatiul
intercelular.
Prin tehnici de criofracturare a fost evidentiat aspectul clivat specific stratului bilipidic al membranei celulare, rezultand jumatatea interna - numita fata P - si jumatatea externa, numita fata E. Pe fata P a membranei se poate observa un ansamblu de fasii din proteine transmembranare anastomozate intre ele, iar pe fata E - o succesiune de "creste" sau santuri corespondente.
|
|
|
Fig. I.1.1. Jonctiune
stransa - organizare moleculara |
Din punct de vedere molecular, principala proteina implicata este ocludina, la care se asociaza proteine de placa (cingulina). Proteinele de placa se leaga cu spectrina, care la randul ei se ataseaza microfilamentelor de actina intracelulare.
In general, acest tip de jonctiune se gaseste in vecinatatea polului apical si se considera ca este principalul element care separa cele doua domenii, apical si laterobazal.
Prin structura sa zonula ocludenta realizeaza o stransa aderenta intercelulara, prevenind miscarea proteinelor membranare din domeniul apical in cel laterobazal. In plus, reprezinta o bariera care regleaza fluxul intercelular si, de asemenea, participa in transportul activ contra unui gradient de concentratie, impiedicand difuziunea retrograda a substantelor transportate. Astfel de jonctiuni se intalnesc in epiteliul intestinal, epiteliul tubilor uriniferi, epiteliul unor canale excretorii glandulare.
|
Observatie |
Jonctiuni aderente
(de ancorare)
Zonula aderenta
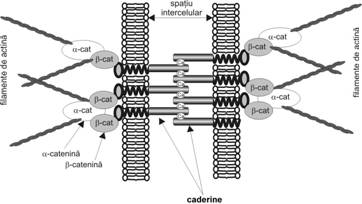
Zonula aderenta (fig. I.1.2) stabileste o ancorare intre doua celule vecine, formand un cordon complet in jurul polului apical. Electrono-microscopic se remarca o densificare a citoplasmei care urmeaza traiectul jonctiunii si care, la rezolutii inalte, prezinta un aspect fin, filamentos. In unele celule (de exemplu celula prismatica intestinala) aceasta densificare se continua cu asa-numitul cadru terminal, reprezentand o zona de citoplasma cu orientare transversala, bogata in filamente de actina cu dispozitie longitudinala (paralel cu si de-a lungul fetei interne a membranei celulare). Spatiul intercelular la nivelul acestui tip de jonctiune masoara 15-20 nm si contine un material de densitate electronica mai mica, cu aspect de fine striatii transversale.
Din
punct de vedere molecular, in structura jonctiunii intra caderine clasice calciu-dependente,
proteine transmembranare de adeziune intercelulara, care formeaza
legaturi homofile-homotipice (vezi subsectiunea 3.1). Spatiul
intercelular este ocupat de domeniul extracelular al caderinelor. Intracelular,
in apropierea jonctiunii, au fost identificate trei tipuri de catenine (α, β si
γ). α-catenina este legata de domeniul intracelular al caderinei
si de filamentele de actina. γ-catenina, considerata
identica cu plakoglobina, se
leaga si ea de caderina si de
α-catenina. Imunohistochimic a mai fost evidentiata
prezenta altor molecule, cum ar fi:
α-actinina si vinculina (care solidarizeaza intre ele si cu
plasmalema filamentele de actina
din cadrul terminal), miozina, radixina si tenuina. Activitatea cateninelor depinde de activarea
protein-kinazei C sub influenta unor protooncogene.
Prin structura sa, zonula aderenta stabileste legaturi intre citoscheletul celulelor adiacente, precum si legaturi intre citoscheletul celulelor din stratul bazal si materialul extracelular. Acest tip de jonctiune se gaseste in vecinatatea jonctiunilor ocludente.
|
Observatie |
Macula aderenta (desmozomul)
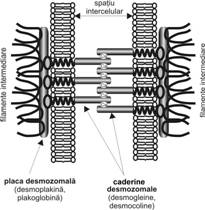
Macula aderenta sau desmozomul (fig. I.1.3) apare pe fetele laterale ale celulelor, ca mici structuri discoidale ovalare cu axul lung de 400-500 nm lungime si axul scurt de 100 nm lungime. Organizarea desmozomului implica existenta a doua celule. Microscopia electronica releva ca intre celule exista un spatiu de aproximativ 30 nm, iar citoplasma adiacenta fetei interne a celor doua plasmaleme prezinta o densificare numita placa de
|
|
|
Fig. I. Zonula aderenta - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
atasament sau desmozomala (400 x 250 x 10 nm), spre care converg filamente intermediare. In sectiune longitudinala, pe mijlocul acestui spatiu se observa o discreta banda sau linie electron-densa.
Principalele molecule prezente in spatiul intercelular apartin a doua grupe de glicoproteine transmembranare, din superfamilia caderinelor (vezi subsectiunea 3.1): (i) desmogleina si desmocolina si (ii) desmocolina . Placa desmozomala contine proteine de atasament neglicozilate: desmoplakina desmoplakina plakoglobina si desmiokina sau polipeptidul bazic. Aceste moleculele stabilesc legaturile cu filamentele intermediare de citokeratina in cadrul tesutului epitelial; acestea se insera in placa, dupa care ies din nou in citoplasma, traiectul fiind in "ac de par".
|
Observatie |
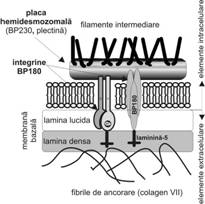
Hemidesmozomul
Hemidesmozomul (fig. I.1.4) are o structura asemanatoare desmozomului, continand aceleasi proteine de placa pe care se insera filamentele intermediare de citokeratina. De data aceasta insa filamentele patrund in placa si raman ancorate la acest nivel, fara a reveni in citoplasma si fara a realiza dispozitia in "ac de par", mai sus mentionata. Moleculele transmembranare sunt insa reprezentate aici de integrine, molecule proteice speciale care recunosc domenii din moleculele matricei extracelulare, legandu-se de laminina si colagenul de tip IV (vezi subsectiunea 3.4).
Hemidesmozomii sunt prezenti la polul bazal al celulelor epiteliului stratificat pavimentos care vin in raport cu membrana bazala, realizand atasarea de aceasta. Ei au rol in mentinerea coeziunii dintre epiteliu si membrana bazala, impiedicand posibila detasare care ar putea surveni in conditiile exercitarii unui stress mecanic important (lovituri, taieturi).
Contactul in focar
Contactul in focar (lat. puncta adherentes) se gaseste tot la polul bazal al celulelor si face legatura cu materialul extracelular. Electrono-microscopic structurile sunt mai greu de observat deoarece nu exista spatiu intercelular evident. Si in cazul contactului in focar, legatura intre citoscheletul celular si matricea extracelulara se stabileste prin intermediul unor integrine.
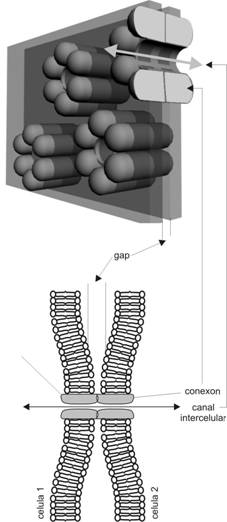
Jonctiuni comunicante
Jonctiunile comunicante (fig. I.1.5) sunt structuri implicate mai putin in stabilirea coeziunii intercelulare, reprezentand cai de comunicare biochimica. Aceste jonctiuni permit difuziunea selectiva intercelulara a ionilor si a unor molecule mici.
Electrono-microscopic se aseamana cu jonctiunile ocludente, fara insa a avea loc o reala fuziune intermembranara; exista un foarte mic spatiu intercelular de aproximativ 3 nm.
|
|
|
|
Fig. I.1.3. Desmozom - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
Fig. I.1.4. Hemidesmozom - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
In acest spatiu se pot observa structuri discrete, sub forma de punti care trec de la o membrana la cealalta sau de pori aposi care proemina, numite conexoni. Prin tehnici speciale ultrastructurale s-a putut demonstra o forma aproape cilindrica a conexonului, cu o lungime de aproximativ 7,5 nm, peretele cilindrului fiind format din 6 subunitati bastoniforme dispuse in jurul unui por central de 1,5-2 nm diametru. Prin aranjarea fata in fata a conexonilor celulelor vecine se asigura o comunicare tip canal, cu un diametru sub 2 nm, ce permite scurgerea unor molecule (aminoacizi, AMP ciclic, ioni, unii hormoni) dintr-o celula in alta.
Din
punct de vedere molecular, cele
6 subunitati bastoniforme sunt de natura proteica si
sunt numite conexine - membrii unei
mari familii de proteine, cu peste 12 molecule recunoscute pana in
prezent.
|
|
|
Fig. I.1.5. Jonctiune gap - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
Din
aceasta familie au fost evidentiate si caracterizate la om conexina 43 (α1),
conexina 32 (β1) si conexina 26 (β2).
O conexina contine 4 domenii transmembranare legate printr-o
bucla intracelulara si doua bucle extracelulare, astfel
incat capetele aminoterminal si carboxiterminal se gasesc intracelular.
Permeabilitatea jonctiunii este modificata de pH si de concentratia ionilor de calciu, care intervin in mecanismul de reglare a jonctiunii, astfel incat aceasta poate fi inchisa sau deschisa. Astfel, daca pH-ul celular este scazut sau concentratia ionilor de calciu este crescuta, canalul de comunicare este inchis; invers, are loc deschiderea canalului.
Jonctiunile comunicante apar in perioada embriogenezei, importanta lor constand in cuplarea electrica a celulelor si in distributia moleculelor informationale in interiorul maselor celulare in curs de migrare. La adult sunt prezente, pe langa tesutul epitelial, in tesutul muscular cardiac, tesutul muscular neted si chiar in tesutul nervos - unde formeaza sinapsele electrice.
|
Observatie |
MOLECULELE DE ADEZIUNE
Moleculele de adeziune celulara
(
In cadrul acestui ansamblu au fost diferentiate 4 superfamilii bine caracterizate: caderinele, selectinele, imunoglobulinele si integrinele; recent insa a fost adaugata si o a cincea familie, deocamdata mai putin studiata, ai carei membri contin importante cantitati de leucina.
Interactiunea CAM poate fi: homotipica, heterotipica, homofila si heterofila. Se pot realiza astfel 4 tipuri de interactiuni:
homotipica si homofila, in cazul in care moleculele de adeziune sunt identice si tipurile celulare implicate identice;
homotipica heterofila, in cazul in care moleculele de adeziune sunt diferite, iar tipurile celulare identice;
heterotipica homofila, in cazul in care moleculele de adeziune sunt identice, iar tipurile celulare diferite;
heterotipica heterofila, in cazul in care si moleculele de adeziune, si tipurile celulare sunt diferite.
Trebuie subliniat faptul ca moleculele de adeziune sunt elemente comune multor celule ale organismului, indiferent de tesutul de apartenenta. In continuare, insa, vom prezenta notiuni care caracterizeaza aceste molecule la nivelul tesutului epitelial, in general, precum si al tesutului epitelial de tapetare in particular.
Caderinele
Caderinele reprezinta o superfamilie de molecule calciu-dependente de natura glicoproteica si cu localizare transmembranara; se intalnesc in diferitele tipuri de jonctiuni intercelulare, in tesuturile adulte.
Aceste molecule formeaza complexe cu molecule intracitoplasmatice numite catenine, acestea din urma fiind localizate sub membrana plasmatica si fixandu-se pe filamentele de actina intracelulara.
Astfel, se poate vorbi despre complexe caderine-catenine.
Pierderea caderinelor favorizeaza fenotipul celular malign, facilitand dezagregarea celulelor care pot astfel invada local sau pot metastaza.
Dupa
localizarea lor, au fost impartite in
doua tipuri: caderine clasice si caderine
desmozomale.
Caderinele clasice au fost evidentiate in jonctiunile aderente, denumirile fiind in corelatie cu celulele la care apartin.
Pentru
tesutul epitelial este specifica
E-caderina, cu rol de mentinere
a integritatii straturilor epiteliale.
|
Observatie |
Caderinele desmozomale sunt prezente in structura desmozomilor si sunt reprezentate de catre desmogleine si desmocoline. S-a putut demonstra ca aceste molecule sunt codificate de gene prezente pe cromozomul 18 si sunt implicate in unele procese patologice (de exemplu in pemfigusul vulgar).
Cateninele
sunt de trei tipuri: α-catenina,
β-catenina, γ-catenina;
γ-catenina, similara cu plakoglobina,
este prezenta impreuna cu desmogleina 1 in structura desmozomilor;
α-catenina si β-catenina se pot lega la membrana celulara
si prin intermediul unor proteine, altele decat caderinele.
Selectinele
Selectinele reprezinta o alta familie moleculara, care asigura adeziunea intre unele elemente figurate din sange (leucocite si plachete) si endoteliul vascular (tesut epitelial). Expresia acestor molecule devine mai puternica sub actiunea unor factori (de tipul citokinelor) care actioneaza ca activatori ai acestor celule, favorizand exprimarea membranara a selectinelor.
Sunt numite selectine datorita faptului ca prezinta in structura un domeniu extracelular aminoterminal, similar cu lectinele implicate in legarea glucozei la mamifere. Selectinele se leaga prin domeniul lor de lectina de forme sialilate de oligozaharide (de exemplu, Lewis 10 sialilat) care, la randul lor, se leaga covalent de diferite glicoproteine asemanatoare mucinelor (de exemplu, GlyCAM-1, PSGL-1, ESL-1 si CD34).
Denumirea
lor este in raport cu tipul celular. Pentru tesutul epitelial
caracteristice sunt:
P-selectina - prezenta pe
celulele endoteliale normale si in
membrana granulelor intracitoplasmatice Weibel-Palade si E-selectina (CD62E, cunoscuta
anterior ca ELAM-1) - prezenta pe celulele endoteliale activate.
|
Observatie |
|
Imunoglobulinele
Aceste molecule sunt incadrate in superfamilia imunoglobulinelor datorita organizarii sub forma de bucle peptidice consolidate prin legaturi disulfidice. Stabilesc adeziuni intercelulare calciu-independente si, spre deosebire de celelalte clase, fara o implicare a structurilor citoscheletale. Pentru tesutul epitelial exista in aceasta clasa cateva molecule de adeziune cu localizare restransa la nivelul celulelor endoteliale: ICAM-1, VCAM-1, PECAM-1.
|
Observatie |
Integrinele
Integrinele constituie o superfamilie moleculara de importanta deosebita prin implicarea lor in comunicarea si interactiunea intre celule si materialul extracelular. Aceste molecule reprezinta de fapt receptori celulari membranari care mediaza atasamentul celular la matricea extracelulara. Au o raspandire extrem de larga, majoritatea celulelor avand mai mult decat o integrina pe suprafata lor. Adeziunea intercelulara este necesara, in unele cazuri, in procesul de proliferare celulara, astfel incat absenta integrinelor poate conduce la apoptoza.
Integrinele sunt proteine transmembranare formate din lanturi α si β; exista 14 tipuri de subunitati α si 8 tipuri de subunitati β, astfel incat se formeaza cel putin 20 de heterodimeri. Un singur lant β poate interactiona cu o serie de lanturi α, producand integrine care pot lega diferite componente ale matricei. Fiecare heterodimer are un domeniu intracelular, un domeniu transmembranar si un domeniu extracelular. Domeniul intracelular (intracitoplasmatic) se leaga, prin intermediul altor molecule intracelulare (talina, vinculina, α-actinina), de filamentele de actina. Domeniul extracelular prezinta situsurile de recunoastere pentru liganzii (moleculele) matricei extracelulare. De exemplu, integrina α2β1 recunoaste, in principal, colagenul de tip IV si laminina, integrina α5β1 recunoaste fibronectina.
Pentru
tesutul epitelial integrinele sunt prezente la nivelul celulelor stratului
bazal, localizat
pe membrana bazala, si sunt responsabile
de realizarea jonctiunii de tip contact
in focar. Acest tip de jonctiune implica legarea integrinelor de
citoschelet. Proteinele intracelulare care participa la formarea
contactului in focar sunt: talina, vinculina, α-actinina, tensina
si paxilina. Odata
asamblat, complexul integrina-citoschelet functioneaza similar
cu receptorii activati si recruteaza componenti ai
sistemelor de semnalare intracelulara.
Pentru
cele mai multe dintre integrine ADN-ul corespunzator a fost clonat si
secventat.
Pe langa un rol important in dezvoltarea embrionara si
mentinerea integritatii tesuturilor adulte, integrinele
reprezinta principalele molecule capabile sa receptioneze
semnale extracelulare destinate unei bune functionari celulare
(de exemplu, inductia expresiilor unor gene).
A fost descrisa si o familie de dezintegrine care functioneaza ca inhibitori ai integrinelor de tip competitiv, fata de liganzii corespunzatori (venin de sarpe). Desi nu in totalitate, majoritatea integrinelor sunt calciu-dependente.
|
Observatie |
POLARIZAREA SI SPECIALIZARILE
Polarizarea poate fi definita ca o organizare asimetrica a celulei, legata de (i) distributia organitelor celulare si de (ii) exprimarea unor caracteristici morfo-functionale diferite, in anumite zone ale suprafetei celulare. Aparitia si dezvoltarea unui anumit pol (domeniu), distinct din punct de vedere morfologic si biochimic, este in stransa dependenta cu functia pe care o indeplineste un anumit tip de celula in cadrul tesutului epitelial.
Marea
majoritate a celulelor, indiferent de tesut, au un anumit grad de polarizare in raport direct cu
functionalitatea acestora. Celulele tesutului epitelial de tapetare,
prin organizarea lor sub forma de adevarate "membrane celulare",
si prin pozitia lor in structura diferitelor organe cu functii
variate (mai ales in unele dintre organele cavitare, de exemplu, tubul
digestiv, caile urinare), prezinta o polarizare deosebit de
evidenta.
La nivelul membranei plasmatice, ca urmare a polarizarii, se diferentiaza
doua domenii: apical si laterobazal.
Domeniul apical
vine in raport cu suprafata libera sau cu lumenul unor organe,
asigurand deopotriva preluarea unor substante necesare celulei
si eliminarea produsilor de secretie. La acest nivel se
remarca prezenta a numeroase canale ionice,
proteine-caraus, enzime hidrolitice si
ATP-aza, precum si un invelis glicoproteic. Domeniul apical
realizeaza, prin organizare moleculara particulara,
specializari sau diferentieri ale membranei
celulare reprezentate de: microvilozitati,
cili ficsi (stereocili), cili vibratili (kinetocili),
placi membranare apicale.
Aceste specializari nu apar in mod obligatoriu la toate
varietatile de epiteliu; fiecare in parte constituie o anume adaptare
functionala, in raport cu pozitia unei varietati
epiteliale in cadrul unui organ.
Domeniul laterobazal vine in contact cu membrana bazala. El se particularizeaza prin faptul ca membrana plasmatica contine majoritatea receptorilor, a enzimelor (inclusiv ATP-aza sodiu-potasiu dependenta), a pompelor si canalelor ionice implicate in functionalitatea celulei, alaturi de moleculele implicate in comunicare intercelulara, adeziune intercelulara (prin interdigitari celulare si jonctiuni specifice) si adeziune la membrana bazala. Totodata, aici exista situsuri pentru secretia celulara. La acest nivel se pot individualiza doua zone: membrana plasmatica laterala si membrana plasmatica bazala.
Cele doua domenii sunt separate de structuri jonctionale specifice, stranse. Cu toate acestea, se pot produce miscari ale moleculelor caracteristice celor doua domenii, principalul rol in miscare fiind atribuit microtubulilor si microfilamentelor - fapt demonstrat de studii experimentale care, prin utilizarea colchicinei si a citochalazinei (substante care interfereaza cu polimerizarea tubulinei si a actinei) determina o dezorganizare a polarizarii.
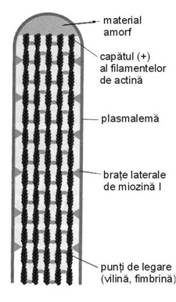
Microvilozitatile
Microvilozitatile (fig. I.1.6) reprezinta expansiuni citoplasmatice delimitate de membrana, mici si neregulate, de inaltime diferita, localizate la polul apical al celulelor epiteliale cilindrice; sunt structuri tranzitorii, motiv pentru care au primit si denumirea de microvilozitati banale. La marea majoritate a celulelor epiteliale existenta microvilozitatilor poate fi confirmata doar in microscopie electronica.
|
|
|
Fig. I.1.6. Microvilozitate - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
Unele celule epiteliale - de exemplu, cele intestinale si cele renale, implicate in absorbtie si resorbtie - prezinta insa o dezvoltare importanta a acestor structuri, care devin o trasatura constanta, cu ultrastructura si organizare moleculara caracteristice.
Clasic,
in microscopia optica, pentru microvilozitatile specifice
celulelor epiteliului intestinal se utilizeaza termenul de platou striat, in timp ce pentru cele
din tubul contort proximal renal se foloseste termenul de margine in perie. Cei doi termeni sunt
uneori diferit utilizati de catre scoala de histologie
franceza si, respectiv,
anglo-saxona. Consideram ca, in fond, nu exista
diferente importante din punct de vedere morfologic si cu
semnificatie functionala care sa justifice utilizarea
tintita a unuia sau celuilalt dintre termeni. In ambele
situatii, microscopia electronica pune in evidenta expansiuni digitiforme, de regula
rectilinii, cu diametrul in jur de 0,1 μm si lungimea de 1-2 μm,
cu un aranjament ordonat si un paralelism apreciabil. Acest fapt face ca,
in microscopia optica, polul apical al celulelor mentionate sa
apara cu o refringenta particulara si cu evidente
striatii paralele intre ele. Aceasta specializare mareste in mod considerabil suprafata
apicala a celulelor, facilitand functii ca absorbtia,
resorbtia si, in mai mica masura, eliminarea
secretiei.
Membrana celulara care inveleste microvilozitatile este acoperita de un strat de filamente foarte fine, ramificate, alcatuit din glicoproteine si enzime, denumit glicocalix. Glicocalixul reprezinta, in fapt, oligozaharidele terminale atasate de proteinele transmembranare specifice plasmalemei si are rol in protectie si recunoastere celulara. In microscopia optica acest strat poate fi vizualizat prin coloratii speciale (PAS) si prin evidentierea histoenzimologica a unor enzime (fosfataza alcalina). Electrono-microscopic acest strat apare amorf, omogen sau foarte fin granular, de densitate electronica medie.
Organizarea moleculara (fig. I.1.6) a structurii microvilozitatilor a fost evidentiata prin metode imunohistochimice. La varful lor se gaseste o zona amorfa a carei compozitie se considera a fi de natura proteica, care permite atasarea unui grup de 25-30 filamente de actina, impiedicandu-le depolimerizarea. Filamentele de actina din axul microvilozitatilor sunt consolidate intre ele prin microfilamente de fimbrina, fascina si vilina. Din zona amorfa descind fascicule paralele de filamente de actina (in sens longitudinal) care se ancoreaza la baza microvilozitatii, in citoplasma celulara apicala, pe o structura numita cadru terminal. Cadrul terminal este format din filamente de actina orientate paralel cu suprafata apicala a celulei (in sens transversal), consolidate unele cu altele si cu membrana apicala prin microfilamente de spectrina si filamente intermediare. Pe fata interna a membranei plasmatice a microvilozitatilor se gasesc, la intervale regulate, molecule de miozina 1 si calmodulina, care fac legatura intre actina din interior si membrana plasmatica, conferind astfel suport structural.
|
Observatie |
Stereocilii
Stereocilii au, in microscopia optica, aspectul unor expansiuni citoplasmatice mult mai
groase si mai lungi comparativ cu microvilozitatile, motiv
pentru care pot fi confundate, uneori, cu cilii.
Din punct de vedere structural, ultrastructural si molecular, stereocilii
reprezinta in fapt microvilozitati mult mai dezvoltate, extrem
de rigide datorita miezului actinic. Acest tip de specializare este
caracteristic celulelor epiteliului epididimului (unde au rol in cresterea
suprafetei de contact) si celulelor epiteliului senzorial din canalul
cohlear - urechea interna (unde au rol in generarea semnalului auditiv).
Cilii vibratili (kinetocilii)
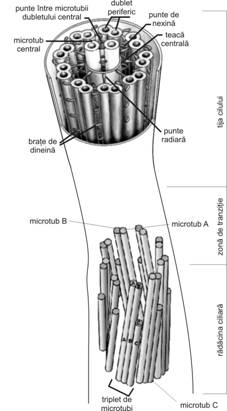
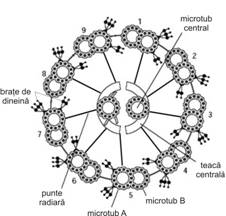
Kinetocilii (fig. I.1.7) reprezinta structuri specializate sub forma de expansiuni filiforme de 0,25 μm in diametru si 7-10 μm lungime, usor vizibili pe sectiuni histologice in microscopie optica. Aceste structuri sunt prezente in numar mare (de ordinul sutelor) la nivelul unor suprafete epiteliale specializate in deplasarea unidirectionala, actionand prin miscari ritmice rapide si facilitand transportul unor materiale fluide (de exemplu, in caile respiratorii si trompele uterine).
Microscopia electronica releva faptul ca un cil este alcatuit din membrana plasmatica la periferie si un citoschelet specializat, denumit axonema, in interior. Axonema este alcatuita dintr-un complex de microtubuli aranjati uniform, in jurul unei perechi centrale fiind dispuse 9 dublete periferice. Pentru perechea centrala microtubulii sunt denumiti C si C fiecare pereche periferica este formata dintr-un microtubul A si un microtubul B (fig. I.1.7). Un microtubul este format din 13 protofilamente, aranjate helicoidal, rezultate din polimerizarea unor subunitati de tubulina α, in alternanta cu subunitati de tubulina β. Toti microtubulii au aceeasi polaritate, cu o extremitate pozitiva (tubulina α) si o extremitate negativa (tubulina β).
In perechea centrala cei doi microtubuli sunt separati intre ei, fiind situati la oarecare distanta si existand o asa-zisa teaca centrala in jurul lor, care este discontinua.
In cazul dubletelor periferice, fiecare contine un microtubul A complet, cu 13 protofilamente (profil circular complet in sectiune transversala), la care este atasat un microtubul B incomplet, format din numai 10 protofilamente (profil circular incomplet in sectiune transversala), astfel incat apare un perete comun - 3 protofilamente apartinand ambilor microtubuli.
Pe
fiecare microtubul A exista scurte prelungiri formate din molecule de dineina, sub forma unor brate
care radiaza catre microtubulul B al dubletului vecin. Aceste
"brate", orientate in sensul acelor de ceas, apar distantate la
intervale de aproximativ 24 nm. Dineina este o proteina care prezinta
o zona cu activitate ATP-azica. Hidroliza ATP-ului determina
deplasarea moleculei de dineina, care imprima astfel o miscare
de flexie cilului, sub forma unei unde. La atasarea unui microtubul A al
unui dublet la microtubulul B al dubletului vecin contribuie si
filamentele de nexina, care se ataseaza la dineina;
moleculele de nexina apar la
intervale de aproximativ 86 nm.
De asemenea, de pe fiecare microtubul A, la
intervale de circa 29 nm, se desprind fibre radiare sau punti radiare (ca
niste spite) ce se indreapta spre perechea centrala,
atasandu-se la teaca centrala.
La
baza cilului, la nivelul polului apical se gaseste o structura -
corpusculul bazal - cu organizare
similara centrozomului (noua grupuri de cate trei microtubuli).
Corpusculul bazal se formeaza din organizatorii procentrozomului; pe
masura ce se adauga dimeri de tubulina procentrozomul se
alungeste, rezultand cele
9 grupuri de cate trei microtubuli. Odata edificat, corpusculul bazal
migreaza in citoplasma apicala, unde va contribui la aparitia
cilului. Din cele noua grupuri de
cate trei microtubuli (triplet) se vor
|
|
|
|
|
Fig. I.1.7. Cil - organizare moleculara - schema dupa (Ross, Kaye, Pawlina, 2003) |
dezvolta noua grupuri de cate doi microtubuli (dublet), iar pentru a asigura organizarea tipica, de 9 dublete periferice si o pereche centrala, se formeaza de novo cei doi microtubuli ai acesteia.
Cuticula (placa membranara)
Histologia clasica descrie cuticula ca o densificare a suprafetei apicale a celulelor stratului superficial din epiteliul de tranzitie (uroteliu). In microscopia optica, cuticula se evidentiaza printr-o tinctorialitate crescuta, comparativ cu cea a citoplasmei subjacente. Microscopia electronica a demonstrat ca, in realitate, cuticula constituie expresia optica a unui mod particular de organizare a membranei plasmatice apicale (comparatia fiind facuta cu aspectul in "solzi de peste"), care are o structura asimetrica - foita externa avand o grosime aproape dubla fata de cea a foitei interne. Foita externa este alcatuita din placi poligonale relativ rigide si groase, despartite intre ele prin punti subtiri de membrana plasmatica nemodificata.
Prin
tehnici de criofracturare s-a constatat ca aceste placi contin particule proteice intramembranare, strans aranjate sub forma hexagonala, de
aproximativ 12 nm; fiecare particula, la randul ei, este
alcatuita din mai multe subunitati. In structura acestor
particule s-au evidentiat 4 tipuri de molecule, denumite uroplakine IA, IB, II si III. Aceste molecule au mai multe
domenii transmembranare, iar domeniile extracelulare sunt mult mai bine dezvoltate
comparativ cu cele intracelulare.
La domeniile intracelulare sunt atasate filamente ale citoscheletului,
permitandu-se astfel plierea membranei apicale cu ajutorul puntilor
de membrana nemodificata, care functioneaza ca arii de tip
balama.
In citoplasma apicala se gasesc structuri sub forma unor vezicule lenticulare, numite vezicule discoide, considerate a fi de origine golgiana. Membrana veziculelor este similara membranei celulare apicale, avand aceeasi asimetrie. Aceste vezicule se pot atasa membranei apicale si se deschid realizand expansiunea acesteia, dupa care se pot detasa si inchide, revenind in citoplasma apicala a celulei.
Placile rigide reprezinta peste 75% din suprafata fetei apicale. Organizarea moleculara a acestei specializari confera celulelor superficiale din uroteliu posibilitatea de a-si mari substantial suprafata apicala, fara o lezare a membranei plasmatice, si de a reveni la forma initiala.
Cand vezica urinara este goala, placile sunt pliate, realizand contururi neregulate, angulare; acest aspect dispare pe masura ce epiteliul se destinde, odata cu procesul de umplere al vezicii. In acelasi timp, cuticula asigura o impermeabilitate mult mai mare - strict necesara, daca tinem cont ca urina din caile excretorii urinare este un lichid hiperton.
Labirintul bazal
Electrono-microscopic, se constata ca polul bazal al unor celule prezinta o dezvoltare importanta de invaginatii relativ profunde ale membranei plasmatice, sub forma de pliuri, compartimentand astfel citoplasma. Se realizeaza o importanta marire a suprafetei bazale - diferentierea ar putea reprezenta, prin analogie, inversul microvilozitatilor. Membrana plasmatica contine la acest nivel numeroase canale si pompe ionice, iar compartimentele citoplasmatice compacte determinate de pliurile bazale sunt foarte bogate in mitocondrii. Aceasta ultrastructura se exprima, in microscopia optica, prin aparitia unui aspect de striuri discrete, aproximativ paralele intre ele si perpendiculare pe polul bazal.
Labirintul bazal
este evident in celule implicate in transport
activ, mai ales ionic, mitocondriile furnizand energia necesara pentru
transportul activ de ioni, cu rol in stabilirea gradientilor osmotici care
asigura miscarea apei
de-a lungul unui epiteliu - de exemplu, in epiteliul tubilor renali
contorti si in epiteliul canalelor de excretie a unor glande
salivare.
CAPACITATEA DE REGENERARE
Celulele
epiteliale au o durata de viata limitata, iar unele dintre
ele sunt eliminate in mod continuu, prin exfoliere sau apoptoza, fiind
inlocuite cu alte celule rezultate prin diviziunea mitotica a unor celule stem (
Pozitia epiteliului de tapetare in contact cu mediul extern sau cu cavitatile interne supune celulele acestuia la un ridicat nivel de uzura, fapt ce impune o permanenta reinnoire a celulelor imbatranite, degenerate, pe seama celulelor stem.
Repartitia celulelor de regenerare este diferita in raport cu varietatea de epiteliu de tapetare. Astfel, in unele varietati de tesut epitelial simplu, celulele de regenerare apar izolate printre celulele diferentiate, fiind greu de evidentiat prin tehnici histologice de rutina.
In alte situatii celulele de regenerare sunt grupate sub forma unui strat regenerator, care apare ca strat bazal datorita contactului cu membrana bazala. Dispozitia este caracteristica pentru tesuturile epiteliale stratificate si pseudostratificate. In acest caz apar frecvent mitozele asimetrice, care asigura permanenta unei rezerve celulare de regenerare; astfel, dintr-o mitoza rezulta de cele mai multe ori doua celule fiice, dintre care una ramane in stadiu nediferentiat, iar cealalta evolueaza spre diferentiere. Date experimentale indica faptul ca, in acest proces, membrana bazala joaca un oarecare rol.
O a treia posibilitate, specifica pentru unele tesuturi epiteliale simple - ca epiteliul gastric si epiteliul intestinal - este reprezentata de existenta de zone de regenerare (zone germinale), in care sunt grupate celulele stem. Dupa mitoze, celulele in curs de diferentiere migreaza de-a lungul membranei bazale, perioada in care se desavarseste diferentierea; si in acest caz, relatiile cu moleculele din membrana bazala joaca un rol important.
Viteza de regenerare
poate fi apreciata prin cativa indici cum ar fi: indicele mitotic
(procentul celulelor in mitoza), indicele de marcaj (procentul de celule
care au incorporat un precursor marcat - timidina tritiata), timpul de
reinnoire
(din momentul aparitiei in celula a precursorului radioactiv
pana la disparitia acestuia).
Procesul de regenerare si diferentiere a celulelor epiteliale este deosebit de complex, fiind determinat si controlat de factori intrinseci (hormoni, factori de crestere, numeroase citokine cu efecte in special paracrine), ca si factori extrinseci (temperatura, radiatii, droguri antimitotice etc.).
VASCULARIZATIA
Traditional, tesutul epitelial a fost considerat ca un tesut avascular, lipsit deopotriva de vascularizatia de tip sanguin si cea de tip limfatic. Acest lucru este real in cazul epiteliilor de tapetare, unde nu exista capilare sanguine si limfatice. Pentru epiteliul glandular, insa, in jurul celulelor secretoare - cu histoarhitectonica speciala - se gasesc numeroase vase, fara insa a se stabili un contact direct.
INERVATIA
Unele varietati de epiteliu de
suprafata poseda terminatii
nervoase senzitive abundente
(de exemplu, epidermul, epiteliul mucoasei bucale, epiteliul mucoasei
vaginale), conferind acestora capacitate de receptie. Epiteliile
glandulare, pe de alta parte, poseda inervatie autonoma.
RELATIA STRANSA CU TESUTUL CONJUNCTIV
Tesutul
epitelial se gaseste intotdeauna intr-o stransa relatie cu tesutul conjunctiv, existand o interdependenta reciproca.
Jonctiunea intre cele doua tesuturi se
realizeaza printr-o structura acelulara (componenta
particulara a matricei extracelulare), denumita membrana bazala. Aceasta structura, cu o
grosime ce variaza intre
10-80 nm si o organizare moleculara complexa (detaliata in
subcapitolul 3 - Membrana bazala), este produsul de sinteza atat al
celulelor epiteliale, cat si al unora dintre celulele tesutului
conjunctiv. Membrana bazala asigura, pe de o parte,
o adeziune ferma a epiteliului
fata de tesutul conjunctiv, iar pe de alta
parte, datorita unei permeabilitati si
difuzibilitati selective, nutritia
celulelor epiteliale pe seama capilarelor sanguine prezente in tesutul
conjunctiv. De asemenea, functioneaza ca o zona de relatie a epiteliului cu
mediul intern pe planul comunicarii. In culturi celulare
s-a putut demonstra ca celulele epiteliale capata o anumita
diferentiere si o anumita histoarhitectonica in cazul in
care culturile contin si celule conjunctive. Astfel, culturi simple (exclusiv epiteliale) vor fi compuse din
celule epiteliale aglomerate sau dispersate, sferice sau poliedrice,
fara aspecte morfologice de diferentiere. In culturile mixte (epitelio-conjunctive) apare o jonctiune
intre cele doua tipuri de celule, limitata de o membrana
bazala, precum si aspecte morfologice de maturare si
diferentiere a celulelor epiteliale.
|
|
|
Fig. I.1.8. Epiteliu simplu pavimentos - schema in MO |
|
|
|
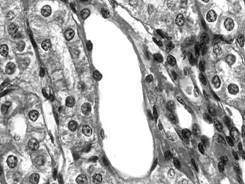
Fig. I.1.9. Epiteliu simplu pavimentos ansa Henle (HE, x 400) |
|
|
|
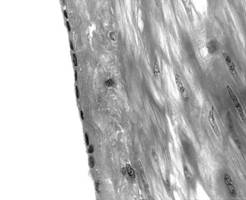
Fig. I.1.10. Epiteliu simplu pavimentos peritoneu (HE, x 400) |
|
|
|
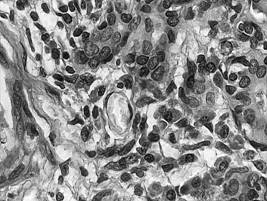
Fig. I.1.11. Epiteliu simplu pavimentos capilar (HE, x 400) |
Copyright © 2024 - Toate drepturile rezervate