|







 |  |
|
| |
 | Afaceri | Agricultura | Economie | Management | Marketing | Protectia muncii |
| Transporturi |
METODE MODERNE DE AMELIORAREA PLANTELOR
1. Generalitati
Prin aparitia unui domeniu nou al geneticii, si anume, genetica moleculara, in scurt timp a fost elucidata structura fina a genei, structura moleculara a cromozomilor la procariote si s-au elaborat metode noi de izolare, sinteza si transfer interspecific al genelor.
Tehnicile si metodele prin care se sintetizeaza artificial genele si se realizeaza manipularile genetice in afara reproducerii sexuate constituie ingineria genetica, unul din domeniile recente ale biologiei moleculare.
Daca metodele ingineriei genetice duc la scurtarea proceselor de obtinere a noilor soiuri, eliminand incertitudinea metodelor clasice, acestea necesita, in schimb, cunoasterea profunda a fenomenelor vietii si nu in ultimul rand dotarea laboratoarelor cu mijloacele necesare modelarii de noi genotipuri. Aplicarea acestor metode este posibila in conditiile culturii "in vitro" a celulelor si tesuturilor vegetale.
Tehnica culturii "in vitro" elaborata independent de G a u t h e r e t si W h i t e, este folosita atat pentru studierea unor probleme fundamentale de fiziologia plantelor cat si in cadrul unor programe cu caracter aplicativ pentru agricultura, horticultura si silvicultura.
Introducerea acestei tehnici a permis acumularea de cunostinte privind totipotenta, diferentierea, diviziunea, nutritia, metabolismul si conservarea celulei vegetale.
In momentul de fata este posibila obtinerea de clone, pornind de la o singura celula iar din meristeme, calusuri sau celule izolate se pot obtine plante prin androgeneza sau embriogeneza.
A devenit posibila obtinerea de plante cu grade diferite de ploidie prin culturi de tesuturi sau de endosperm.
La unele specii se pot obtine organisme haploide prin cultura embrionilor dupa o hibridare sexuata interspecifica urmata de eliminarea cromozomilor unuia dintre genitori.
In inmultirea vegetativa a unor specii pomicole, floricole sau la vita de vie, se recurge tot mai des la culturi de meristeme in vederea obtinerii unor plante libere de viroze.
Depasirea barierelor legate de sterilitate sau de incompatibilitate intre genitori este posibila prin culturile de embrioni, ovule sau ovare.
Prin culturi de tesuturi sau de celule "in vitro", denumim generic, cresterea pe medii sintetice, sterile, a unor fragmente vegetale (explante sau inoculi) macroscopice (culturi de organe) sau microscopice (tesuturi, celule, protoplasti).
O cultura se considera reusita atunci cand celulele explantului, respectiv inoculului, se multiplica si pot fi efectuate subculturi precum si daca materialul obtinut se mentine in timp si raspunde pozitiv tratamentelor de directionare a proceselor de morfogeneza, de organogeneza, potrivit obiectivelor urmarite de ameliorator.
Explantul cultivat "in vitro" constituie o unitate vie, independenta de locul de unde a fost prelevat, care detine integral informatia genetica preluata de la planta de origine, precum si toate functiile pe care le indeplinesc tesuturile respective.
Provenienta explantelor, natura lor, specia, starea fiziologica si dimensiunile, sunt factori care determina acomodarea si integrarea acestora in conditiile mediului aseptic, in reluarea proceselor de crestere si dezvoltare.
Un factor, de asemenea, determinant il constituie compozitia mediului de cultura.
Un mediu de cultura standard consta dintr-un amestec echilibrat de macro si microelemente (cloruri, azotati, sulfati, fosfati, si ioduri de calciu, magneziu, potasiu, azot, mangan si bor), vitamine, o sursa de carbon, factori organici de crestere (aminoacizi, uree si peptone), o sursa de azot redus si fitohormoni.
Ca urmare a cercetarilor efectuate au fost introduse treptat, trei categorii de medii si anume: de inducere, de crestere si de diferentiere.
Materialul vegetal folosit este format din segmente de tulpini si flori, parenchim cortical, vascular si medular, tesuturi epidermale si subepidermale, portiuni de tesut cambial, parenchim din tuberculi, petale, radacini, varfuri de crestere, mezofilul frunzelor, cotiledoane, ovule, ovare, endosperm, fragmente de antere etc.
Inainte de a fi folosit, materialul biologic se va steriliza cu ajutorul unor solutii de apa bromurata 1%, hipoclorit 1% sau clorura mercurica 0,1%. Manipularea materialului biologic este obligatoriu sa fie facuta in conditii de asepsie totala, in camere cu presiune pozitiva sau sub hote cu flux laminar.
2. Crearea de linii izogene prin androgeneza
Asa cum s-a aratat anterior, la plantele superioare exista o alternanta regulata intre cele doua faze, diploida si haploida.
Celula ou sau zigotul, rezultat in urma fecundarii, va da nastere plantei diploide, care in celulele somatice contine un numar dublu de cromozomi (2n).
Tabelul 1.
Compozitia unor medii de baza folosite pentru
cultura anterelor (mg/l)
|
Elementele |
Murashige si Skoog (1962) |
White (1963) |
Nitsch (1969) |
|
MACROELEMENTE | |||
|
Ca(NO3)2 . 4 H2O | |||
|
KNO3 | |||
|
NH4NO3 | |||
|
KCl | |||
|
KH2PO4 | |||
|
NaH2PO4 . 4 H2O | |||
|
CaCl2 . 2 H2O | |||
|
MgSO4 . 7 H2O | |||
|
Na2SO4 | |||
|
MICROELEMENTE | |||
|
Fe2(SO4)3 |
| ||
|
FeSO4 . 7 H2O | |||
|
Na2EDTA | |||
|
MoSO4 . 4 H2O | |||
|
H3BO3 | |||
|
ZnSO4 . 4 H2O | |||
|
Kl | |||
|
Na2MnO4 . 2 H2O | |||
|
CuSO4 . 5 H2O | |||
|
CaCl2 . 6 H2O | |||
|
COMPU{I ORGANICI | |||
|
Biotina | |||
|
Glicina | |||
|
Inozitol | |||
|
Acid nicotinic | |||
|
Piridoxina - HCl | |||
|
Tiamina - HCl | |||
|
Acid folic | |||
|
Zaharoza |
La maturitate, in floare, la nivelul androceului si gineceului are loc diviziunea meiotica in urma careia iau nastere gametofitul mascul si femel.
Gametii care se vor forma contin un numar redus la jumatate de cromozomi (n), caracteristic pentru fiecare specie.
La plante, gametofitul mascul este reprezentat prin graunciorul de polen care contine doi nuclei (vegetativ si spermatic), iar gametofitul femel prin sacul embrionar, celulele sinergide si antipode.
In momentul germinarii graunciorului de polen, nucleul spermatic se divide, rezultand doi nuclei, din care unul va fecunda oosfera si va forma zigotul (2n), iar cel de-al doilea va fecunda nucleul secundar al sacului embrionar, formand endospermul.
In mod natural, cu o frecventa mica, sau artificial, unul din cei doi nuclei ai graunciorului de polen, de obicei cel vegetativ, poate intra intr-o diviziune celulara rezultand, in cazul acesta, o planta care are in celulele corpului un numar redus la jumatate de cromozomi (n), deci o planta haploida.
Fenomenul prin care ia nastere o planta haploida dintr-o celula mascula poarta numele de androgeneza.
Androgeneza a fost indusa pentru prima data prin cultura 'in vitro' a anterelor la Datura inoxia, de catre G u h a si M a h e s w h a r i (1964), iar mai tarziu s-a reusit crearea de haploizi si la alte genuri de plante. Cultura de antere ridica, insa, un impediment. Pe langa plante haploide, din tesuturile anterei pot rezulta si plante diploide.

In anul 1971, S u n d e r l
a n d si R o b e r t s au elaborat o metoda
eficienta de cultivare 'in vitro' a graunciorilor de polen, pe medii
de cultura artificiale corespunzatoare.
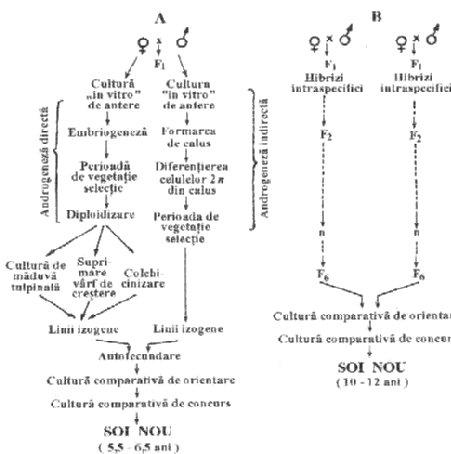
Androgeneza este de doua tipuri: directa si indirecta (fig. 1).
Fig. 1. Haploidia prin androgeneza
Prin androgeneza directa, nucleul haploid, de regula cel vegetativ, modificandu-si programul normal de dezvoltare, intra in procesul de diviziune mitotica, formandu-se mai intai un embrioid si apoi o planta haploida.
In cazul androgenezei indirecte, diviziunile repetate ale nucleului haploid formeaza mai intai un tesut nediferentiat, numit calus.
Prin cultura calusului pe medii de cultura speciale, care contin si substantele chimice necesare diferentierii celulelor, se pot forma fie embrioizi, fie lastari si radacini, din care rezulta apoi plante intregi.
Formele poliploide apar datorita multiplicarii numarului de cromozomi, fara ca membrana nucleara sa se fregmenteze, fenomen numit endomitoza.
Obtinerea plantelor haploide prin androgeneza este conditionata de o serie de factori, cum ar fi: varsta anterelor, temperatura, lumina, compozitia chimica a mediului de cultura etc.
Haploidia prin androgeneza prezinta o importanta deosebita. Avand un numar redus la jumatate de cromozomi, si anume cate un exemplar din fiecare pereche, se vor manifesta fenotipic atat genele dominante, cat si cele recesive.
Se poate afirma ca, la haploizi, structura genetica sau genotipul reflecta cel mai fidel fenotipul. Pentru ameliorarea plantelor, haploidia reprezinta o realizare cu adevarat promitatoare, in realizarea de soiuri si hibrizi.
Trebuie precizat ca, datorita starii haploide, procesul de formare a celulelor sexuale este puternic pertubat, de aceea aceste organisme sunt in totalitate sterile. Prin diverse metode, dar in mod curent prin tratamente cu colchicina, se dubleaza numarul de cromozomi ai haploizilor, revenindu-se la starea diploida, normala si, deci, fertila.
Prin diploidizare plantele vor avea pe cromozomii omologi gene identice, prin urmare vor fi complet homozigote.
Plantele diploide obtinute prin dublarea numarului de cromozomi ai plantelor haploide se numesc linii izogene, pure genetic, puritate care se mentine prin autopolenizarea stricta, in absenta mutatiilor.
O caracteristica importanta a liniilor izogene este variabilitatea genotipica pronuntata, de la linie la linie, deoarece fiecare grauncior de polen poate diferi de altul, datorita recombinarilor genetice ce pot avea loc in procesul de meioza.
La recombinarile genetice se adauga si mutatiile, care la plantele haploide se vor manifesta in totalitate fie ca sunt dominante, fie ca sunt recesive.
Importanta folosirii liniilor dublu haploide in procesul de ameliorare deriva din identificarea perechilor de alele care conditioneaza un anumit caracter. In acest caz, prin autopolenizari controlate se asigura o uniformitate perfecta a indivizilor in cadrul unei generatii sau de la o generatie la alta.
Metoda obtinerii de linii izogene a fost asimilata ca metoda de ameliorare pentru urmatoarele avantaje (M i n o d o r a P a t r a s c u, 1981):
- economie de timp in producerea liniilor homozigote valoroase si, implicit, scurtarea perioadei de obtinere a noilor soiuri sau hibrizi (fig. 2);
- posibilitatea izolarii unui mare numar de genomuri individuale necesare lucrarilor de recombinare si selectie;
- identificarea unor combinatii utile de gene dintr-un volum de material mult mai redus, comparativ cu metodele conventionale;
- posibilitatea purificarii
si omogenizarii generatiilor avansate de selectie;
Fig.2. Ciclul de obtinere a soiurilor prin metoda androgenezei (A),
comparativ cu metoda clasica (B).
- evaluarea si selectia practicate in cadrul unui material care poseda numai gene in stare homozigota;
- estimarea rapida a capacitatii combinative a genitorilor, prin utilizarea unui volum foarte redus de material;
- realizarea homozigotiei complete, fara a supune materialul la presiunea selectiei determinata de unele conditii de mediu nespecifice;
- posibilitatea concentrarii lucrarilor de creare a noi linii intr-un singur centru, eliminandu-se, astfel, alterarea plasticitatii ecologice;
- fixarea primei generatii hibride provenite de la combinatii valoroase sub forma de linii izogenice;
- fixarea gametilor in forme stabile, reproductibile, in stadiu timpuriu (F1 - F3), evitandu-se, astfel, pierderea unor combinatii valoroase;
- simplificicarea sistemului de ereditate tetrasomica al unor forme aneuploide;
- reducerea numarului de indivizi implicati in programul de ameliorare (tab. 2).
Tabelul 2
Numarul de combinatii posibile la formele diploide si haploide, in functie de numarul de gene care conditioneaza un caracter
|
Forma |
Numarul de gene care conditioneaza un caracter |
||||||||
|
Diploida | |||||||||
|
Haploida |
| ||||||||
Pe aceasta cale procesul de ameliorare beneficiaza de o scurtare considerabila a timpului de obtinere a liniilor pure genetic, de micsorarea volumului de lucru si de o precizie mai mare in identificarea indivizilor homozigoti.
Daca prin consangvinizare sunt necesare 8-9 generatii, pentru obtinerea liniilor pure, prin haploidie, sunt suficiente numai 1-2 generatii.
Haploidia prin androgeneza s-a aplicat cu succes la un numar mare de specii cultivate.
La noi in tara, colectivul de geneticieni de la Universitatea din Bucuresti (R a i c u, B a d e a, G r e g o r i a n , S c r i p c a r u) au obtinut in perioada 1976-1980 haploizi la tutun, petunie si Datura inoxia, folositi apoi in procesul de ameliorare ca genitori.
3. Hibridarea somatica
Unirea a doua organisme superioare pe o alta cale decat cea a hibridarii sexuate este cunoscuta sub numele de hibridare somatica.
Aceasta unire este posibila prin fuziunea a doua celule somatice carora li s-a indepartat peretele celular, rigid, pecto-celulozic, celule numite protoplasti.
La inceputul secolului al XX-lea indepartarea membranei pecto-celulozice s-a realizat pe cale mecanica, metoda enzimatica elaborandu-se abia in anul 1960 de catre C o c k i n g , in Anglia.
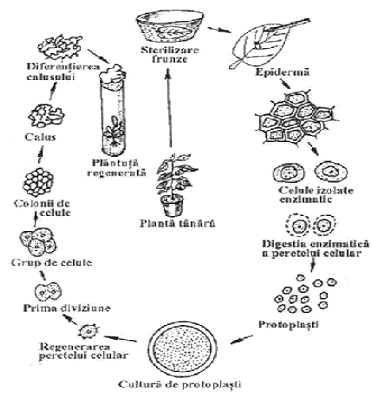
Protoplastii pot fi izolati
din diferite tesuturi vegetale: mezofilul sau epiderma frunzei, radacinii,
nodozitati ale radacinilor, petale, graunciori de polen, din culturi de celule
'in vitro' etc. Materialul folosit curent il constituie epiderma
inferioara a frunzelor. In urma tratamentului enzimatic, celulele se izoleaza
prin dizolvarea lamelei mediane, iar apoi este dizolvata si membrana
pectocelulozica, rezultand, astfel, protoplastii (fig. 3).
Fig.3. Obtinerea de protoplasti
Acestia, cultivandu-se pe medii de cultura adecvate, s-a constatat ca isi refac in cateva zile membrana celulara, iar apoi, prin diviziuni mitotice, formeaza mai intai un tesut nediferentiat denumit calus, din care pot regenera plante mature (in aproximativ 4-6 luni).
Protoplastii ofera numeroase avantaje pentru ameliorarea plantelor: se pot cultiva intr-un numar foarte mare de medii artificiale, pot fuziona intraspecific sau interspecific, pot fi folositi ca receptori de ADN exogen in transformarile genetice, pentru transferul de cromozomi, nuclee sau organite citoplasmatice.
Prin culturi de celule si protoplasti se poate realiza si o variabilitate genetica enorma, folosita apoi in selectia formelor utile.
Comparativ cu hibridarea sexuata, hibridarea somatica prezinta superioritate prin avantajele oferite (P a t r a s c u, 1981), permitand:
- realizarea hibridarilor intre organisme foarte indepartate pe scara evolutiei care, obisnuit, nu se pot hibrida pe cale sexuata;
- obtinerea de hibrizi care nu mai segrega in urmatoarele generatii;
- crearea de hibrizi intre plante care se reproduc vegetativ;
- obtinerea hibrizilor intre plante care nu se pot incrucisa, datorita unor anomalii in morfogeneza genitorilor;
- obtinerea de hibrizi asimetrici care sa posede material genetic de la unul din parinti si numai cativa cromozomi (sau gene, sau numai citoplasma si organitele celulare) de la celalalt partener;
- realizarea hibrizilor multiparentali rezultati in urma fuziunii simultane a mai mult de doua celule parentale;
- obtinerea hibrizilor heterozigoti pentru genele extranucleare.
Cercetarile au demonstrat ca fuziunea spontana a protoplastilor are loc cu o frecventa diferita de la specie la specie. In ultimul timp s-au gasit metode de marire a frecventei celulelor fuzionate, in special prin folosirea unor agenti chimici.
O problema ce trebuie rezolvata in cazul hibridarii somatice este selectia celulelor hibride si eliminarea formelor parentale.
In acest sens s-au elaborat medii de cultura selective, lipsite de fitohormoni (S m i t h s.a., 1976), sau medii la care se adauga o serie de substante, cum ar fi actinomicina D (C o c k i n g s.a., 1976).
Selectia hibrizilor celulari se poate face si folosind liniile mutante albinotice, lipsite de clorofila. Realizandu-se fuzionarea protoplastilor unei linii albinotice dintr-o specie cu protoplastii cu clorofila de la alta specie, se selecteaza apoi coloniile de culoare verde, formate ca rezultat al cresterii selective.
Hibridarea somatica a pus in evidenta un fenomen important, si anume, pierderea preferentiala a cromozomilor uneia din cele doua specii din genomul hibridului. De aceea, in ameliorarea plantelor se pot folosi fie amfidiploizii, rezultati prin fuziunea protoplastilor celor doua specii, fie formele la care s-au transferat numai cativa cromozomi sau fragmente de cromozomi pe care se gasesc anumite gene utile.
Desi cercetarile privind cultura si fuziunea protoplastilor sunt destul de recente, s-a reusit inducerea fuziunii la un numar mare de specii.
Totusi, regenerarea completa a plantelor hibride s-a realizat, in general, la speciile la care hibrizii se pot obtine usor pe cale sexuata.
Un alt aspect important al hibridarilor somatice la plante il reprezinta organitele citoplasmatice ce detin informatie genetica (cloroplastele, mitocondriile, particulele infectioase).
In procesul de hibridare somatica devine posibila manipularea genetica si a acestor organite, putand fi obtinuta o gama larga de organisme: cu nucleu de la un genitor sau de la ambii, cu cloroplastele de la altul sau de la ambii, la fel putandu-se intampla cu mitocondriile si particulele infectioase.
Primele hibridari somatice la plante au fost realizate in anul 1972 de C a r l s o n s.a. intre doua specii ale genului Nicotiana: Nicotiana glauca (2n = 24) si Nicotiana langsdorffii (2n = 18).
Ulterior, s-au realizat hibrizi somatici intre protoplasti de cartof si tomate (hibrid denumit 'Pomato'), intre Arabidopsis si Brassica (denumit 'Arabidobrassica'), dar care, deocamdata, nu au valoare comerciala.
In ultimii ani s-a pus la punct o noua metoda ce induce fuziunea protoplastilor, cu o eficacitate mare, de aproape 100%, denumita 'electrofuziune celulara' (Z i m m e r m a n n s.a., 1981). Aceasta metoda se bazeaza pe folosirea unui camp electric uniform, pentru alaturarea protoplastilor, iar fuzionarea lor este produsa de o variatie scurta, de o anumita intensitate a curentului electric.
Desi pana la ora actuala nu au fost obtinuti hibrizi somatici cu valoare agronomica ridicata, aceasta metoda ramane o cale de mare perspectiva in invingerea barierelor dintre specii, genuri sau alte unitati sistematice.
4. Provocarea mutatiilor 'in vitro'
Este cunoscut faptul ca frecventa mutatiilor naturale (spontane) este variabila, depinzand de specie, individ sau tipul de celule.
Utilizarea factorilor mutageni (fizici, chimici sau biologici) ridica foarte mult frecventa mutatiilor, dar numarul mic de indivizi cu care se lucreaza face ca rezultatele sa nu fie intotdeauna incununate de succes.
Culturile de celule si tesuturi 'in vitro' reprezinta un material foarte valoros pentru crearea mutatiilor genice din urmatoarele considerente: se pot trata cu agenti mutageni si se poate cerceta un numar enorm de celule, intr-un spatiu restrans; este evitata formarea de himere (care apar frecvent la organismele multicelulare); celulele mutante pot fi izolate cu multa usurinta.
Organismele haploide constituie un material initial foarte util pentru inducerea mutatiilor, datorita faptului ca fiecare locus este reprezentat de o singura alela (starea de homozigotie).
La aceste organisme actiunea agentilor mutageni este mai eficienta si poate fi usor controlata. Protoplastii haploizi sunt, de asemenea, utilizati pentru inducerea mutatiilor "in vitro", deoarece toate genele se manifesta fenotipic. In cazul dublarii numarului de cromozomi, genele mutante ajung in stare homozigota.
Includerea in mediul de cultura a agentilor mutageni creeaza o gama foarte variata de mutatii biochimice si metabolice: auxotrofe (caracterizate prin incapacitatea de a sintetiza unele baze azotate din acizii nucleici, aminoacizi, vitamine), mutatii ce confera o buna adaptare la temperaturi ridicate, salinitate, aciditate etc.
Mutatii biochimice si metabolice au fost induse pentru prima data si de P. S. C a r l s o n (1967, 1970) la haploizii de tutun.
Cu toate dificultatile legate de cultura celulelor normale sau a protoplastilor diploizi sau haploizi, faptul ca agentii mutageni pot fi incorporati in mediul de cultura si faptul ca o celula ce prezinta o mutatie poate fi usor regenerata, confera mutagenezei "in vitro" o eficienta foarte mare sub aspect economic, cat si sub aspectul duratei de obtinere a unei forme mutante.
S-au obtinut linii celulare de tutun rezistente la boala denumita "focul salbatic" (Pseudomonas tabaci) prin introducerea in mediul de cultura a toxinei produsa de aceasta bacterie.
Pentru inducerea mutatiilor rezistente la pesticide, in mediul de cultura al celulelor se introduc, in diferite concentratii, erbicide, insecticide, fungicide etc. Pentru majoritatea celulelor aceste substante au efect letal, insa unele celule, in urma unor mutatii genice, manifesta rezistenta, putand fi apoi selectate.
In acelasi mod s-au obtinut linii celulare mutante dotate cu o rezistenta mare la salinitate, rezistente la seceta, la temperaturi coborate.
Copyright © 2024 - Toate drepturile rezervate